

Экология, 2021, № 2, стр. 134-142
Пространственные вариации обилия рыб в дельте Меконга
Е. П. Карпова a, b, c, *, А. Р. Болтачев a, b, c, Э. Р. Аблязов a, b, c, Д. Н. Куцин a, b, c, Ку Нгуен Динь c, Чыонг Ба Хай c, Нгуен Ван Тхинь c, Чинь Тхи Лан Чи c
a Федеральный исследовательский центр им. А.О. Ковалевского РАН
299011 Севастополь, просп. Нахимова, 2, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
c Южное отделение Российско-Вьетнамского тропического центра
Хошимин, Вьетнам
* E-mail: karpova_jey@mail.ru
Поступила в редакцию 20.09.2019
После доработки 21.02.2020
Принята к публикации 04.03.2020
Аннотация
Впервые получены количественные оценки плотности рыбного населения в дельте р. Меконг на территории Социалистической Республики Вьетнам. Исследования проводили в основном течении двух главных рек – Хау и Тиен, на которые разделяется Меконг в своем нижнем течении, и в устьевых рукавах дельты. При сравнении показателей обилия рыб в 28 районах дельты найдены статистически значимые отличия средней удельной численности между основным течением рек и миксогалинной зоной эстуарного экотона, в которой показатели обилия рыб значительно ниже. При корреляционной оценке связей динамики обилия рыб с различными экологическими факторами природного и антропогенного характера выявлена слабая достоверная связь с показателями солености, удаления от устья, типом рыбного сообщества и количеством мусора антропогенного происхождения на дне. В дельте наблюдаются значительные флуктуации показателей обилия рыб, связанные с агрегированным распределением таких видов, как Polynemus melanochir, Arius maculatus, и ряда представителей семейств Akysidae, Cynoglossidae и Ariidae. Полученные результаты могут быть использованы как базовые для последующего мониторинга динамики обилия придонных сообществ рыб дельты Меконга.
Дельта реки Меконг является одной из самых обширных в мире, ее экосистема отличается очень сложной структурой, высокой динамичностью характеристик местообитаний, уникальным видовым богатством гидробионтов и исключительной значимостью для сельского хозяйства и рыболовства. Рыболовство в бассейне реки является одним из самых продуктивных в мире. Согласно ФАО, 2% всей рыбы, пойманной в результате морского и внутреннего рыболовства в мире, получают из бассейна р. Меконг [1], причем около 2.6 млн т добывается ежегодно из бассейна нижнего Меконга [2]. Промысловый лов рыбы в речных системах обычно недооценивается, в том числе в официальной статистике [3].
Оценки вылова от внутреннего рыболовства в развивающихся странах, как правило, основаны на более масштабных данных по аквакультуре и уловов коммерческого рыболовства в озерах или очень крупных реках. Согласно комплексным оценкам, промысел в Меконге составляет от 7 до 22% мировой добычи пресноводной рыбы, при этом около 29% вылова приходится на Вьетнам. В настоящее время средний улов на одного рыбака имеет тенденцию к снижению, но участие населения в промысле очень высоко. Хотя общий вылов, по-видимому, достаточно стабилен из года в год, все больше людей ловят рыбу, и уловы на одного рыбака стали меньше, чем в прошлом. Беспорядочный промысел в бассейне Меконга изменяет структуру рыбного сообщества, приводя к истощению видового разнообразия, деградации популяционной структуры видов, что, по-видимому, подвергает их высокому риску [4]. В первую очередь это относится к видам донно-придонного комплекса. Добыча песка, гравия и ила, ведущаяся всеми странами бассейна Меконга, достигла огромных масштабов, сравнимых с экологической катастрофой. Исследования показывают [5], что общий объем (34.48 млн м3, или 55.2 млн т) донных отложений был добыт в основном течении Меконга в 2011 г., из них около 22% (7.75 млн м3) приходится на Вьетнам.
Разрушение донных биотопов лишает среды обитания целый ряд видов, для многих из которых доказана тесная связь с донными отложениями [4]. Интенсивный промысел ведется в реке с использованием донных тралов, при этом происходит полное изъятие всех водных живых ресурсов на дне и в придонном слое воды. При этом, несмотря на высокую продуктивность, система Меконга недостаточно изучена по многим аспектам ее ресурсов и экологии, включая рыб и беспозвоночных [6–8]. Исследования рыбных ресурсов были локальными и в основном сосредоточены на биологических оценках богатства и разнообразия, а также добычи ресурсов на промысловое усилие [9–11]. Первые исследования удельной численности и биомассы рыб донно-придонного комплекса в системе р. Тиен бассейна Меконга в меженный период были проведены в 2018 г. [12].
В целом для дельты Меконга характерен чрезвычайно высокий негативный антропогенный пресс, который обусловлен несколькими основными факторами. Это прежде всего очень высокая плотность населения и промышленных центров не только непосредственно в границах южной части Вьетнама, но и вдоль всего течения реки – от Китая до Камбоджи, следствием которой является чрезмерный сброс промышленно-бытовых стоков. В дельте Меконга Вьетнама выращивается около 47% злаков, преимущественно риса, а применяемые для этого удобрения и пестициды также сбрасываются в дельту. Одна из причин эвтрофирования и загрязнения антибиотиками – обильно развитая в дельте аквакультура рыбы и креветок, ежегодный объем которой превышает 2 млн т. В связи с интенсивным судоходством воды и донные отложения также загрязнены нефтепродуктами. Поэтому для лучшего мониторинга, управления и сохранения промыслов в нижнем Меконге необходимо обновить информацию о пространственном и временно́м разнообразии рыб, структуре сообществ и моделях распределения, особенно с учетом растущих экологических воздействий, которые имеют место в бассейне реки. Цель нашей работы – оценить пространственную динамику обилия рыб донно-придонного комплекса в нижнем Меконге на территории Вьетнама и проанализировать их зависимость от различных экологических факторов естественной и антропогенной природы. Актуальность поставленной задачи определяется тем, что сохранение и рациональное использование водных биологических ресурсов дельты р. Меконг во многом может определить уровень продовольственной независимости Вьетнама на долговременный период, а также сохранить уникальную природу этого региона для будущих поколений.
МАТЕРИАЛ И МЕТОДИКА
Материал собран в меженный период в нижней части дельты Меконга на территории Социалистической Республики Вьетнам в основном течении двух рукавов Меконга – реках Хау (Бассак) и Тиен (Меконг), и восьми устьях –Тиеу, Дай, Балай, Хамлуонг, Кочьен, Кунг-Хау, Динь Ан и Чань Де, на которые разделяются реки Тиен и Хау при впадении в море. В период с 06.04.2018 г. по 10.06.2018 г. и с 17.01.2019 г. по 29.03.2019 г. было выполнено 224 траления промысловым бимтралом шириной 4 м и высотой 0.4 м с ячеей сетного полотна 10 мм, который повсеместно используется местными рыбаками для промысла рыбы. Траления были сгруппированы в 28 локалитетов (рис. 1), в пределах которых незначительно менялись такие гидрологические параметры, как сток и морфология русла. Основные характеристики участков приведены в табл. 1.
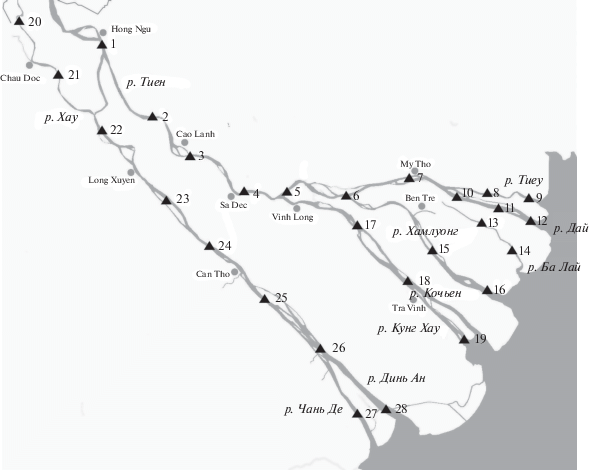
Таблица 1.
Данные о траловых ловах на различных участках дельты Меконга
| № участка* | Кол-во тралений | Координаты, град | |||
|---|---|---|---|---|---|
| начала участка | окончания участка | ||||
| с.ш. | в.д. | с.ш. | в.д. | ||
| 1 | 18 | 10.8056 | 105.2498 | 10.5839 | 105.4299 |
| 2 | 6 | 10.5359 | 105.4908 | 10.5012 | 105.5611 |
| 3 | 6 | 10.4097 | 105.6104 | 10.4014 | 105.6931 |
| 4 | 6 | 10.2969 | 105.7750 | 10.3033 | 105.8384 |
| 5 | 9 | 10.2799 | 105.9175 | 10.3032 | 106.0372 |
| 6 | 8 | 10.2852 | 106.0593 | 10.2500 | 106.2538 |
| 7 | 5 | 10.3278 | 106.2751 | 10.3400 | 106.3255 |
| 8 | 7 | 10.3100 | 106.4916 | 10.2764 | 106.6499 |
| 9 | 8 | 10.2790 | 106.6583 | 10.2706 | 106.7421 |
| 10 | 4 | 10.2968 | 106.4536 | 10.2603 | 106.5501 |
| 11 | 4 | 10.2428 | 106.6102 | 10.2088 | 106.7006 |
| 12 | 3 | 10.1980 | 106.7160 | 10.1700 | 106.7790 |
| 13 | 4 | 10.1834 | 106.5814 | 10.1474 | 106.6326 |
| 14 | 4 | 10.1362 | 106.6500 | 10.0249 | 106.6979 |
| 15 | 12 | 10.1719 | 106.3651 | 9.9969 | 106.5241 |
| 16 | 4 | 9.9866 | 106.5665 | 9.9406 | 9.9406 |
| 17 | 4 | 10.2653 | 105.9544 | 10.1677 | 106.1764 |
| 18 | 8 | 10.0634 | 106.2597 | 9.9758 | 106.3574 |
| 19 | 7 | 9.8638 | 106.4688 | 9.7097 | 106.5861 |
| 20 | 6 | 10.8538 | 105.0885 | 10.7834 | 105.1218 |
| 21 | 16 | 10.7068 | 105.1340 | 10.5982 | 105.2253 |
| 22 | 11 | 10.4845 | 105.3464 | 10.3957 | 105.4339 |
| 23 | 12 | 10.3026 | 105.5198 | 10.1914 | 105.6194 |
| 24 | 10 | 10.1760 | 105.6315 | 10.0555 | 105.8000 |
| 25 | 11 | 10.0155 | 105.8149 | 9.8831 | 105.9497 |
| 26 | 15 | 9.8515 | 105.9804 | 9.7010 | 106.1520 |
| 27 | 11 | 9.6431 | 106.1306 | 9.4593 | 106.2413 |
| 28 | 5 | 9.6451 | 106.2033 | 9.5683 | 106.3030 |
При помощи портативного эхолота определяли координаты начала и окончания тралений, продолжительность и длину пути, пройденного тралом. Траления осуществляли в донном варианте, они длились преимущественно 15–20 мин, дистанция составляла от 206 до 4395 м, в среднем около 1500 м, диапазон глубин менялся от 1.5 до 49 м, в пределах одного траления размах обычно не превышал 5 м. Во время каждого траления отбирали пробы воды у поверхности и дна при помощи батометра и замеряли их соленость и температуру. Температура как поверхностного, так и придонного слоев воды изменялась мало – от 28 до 31.5°С, соленость воды в миксогалинной зоне достигала 30‰.
Анализ уловов включал таксономическую идентификацию и подсчет количества экземпляров рыб каждого семейства, определение их массы, а также массы растительного субстрата и мусора антропогенного происхождения (макропластика), поднятых тралом.
Выбор конструкции трала и методики лова были обусловлены, с одной стороны, необходимостью получения репрезентативной и сопоставимой информации о численности и биомассе рыб донно-придонных сообществ в различных частях дельты [12], а с другой – широким использованием местными рыбаками аналогичных бимтралов для промысла в дельте Меконга, в связи с чем на основании полученных данных можно оценить различные аспекты воздействия донных тралений на фауну рыб.
На основании полученных данных о численности и массе уловов и площади дна, обловленной тралом, проводился подсчет удельной численности (N, экз/100 м2) и биомассы (W, г/100 м2) рыб. Коэффициент уловистости бимтрала принимали равным единице ввиду невозможности его определения в условиях дельты, в связи с этим полученные значения количественных показателей соответствовали минимальным.
Ввиду ненормальности распределения исходных данных по удельной численности и биомассе рыб, а также “остатков” при проведении тестов, в том числе при попытке нормализации с помощью логарифмирования, для проверки достоверности различий в показателях обилия рыб (удельной численности N и биомассы W) на различных участках дельты использовали критерий Краскела-Уоллиса. В целях поиска взаимосвязи между экологическими факторами воздействия на рыбные сообщества и показателями обилия был проведен анализ методами многомерной статистики. Построение корреляционной матрицы проводилось на основе коэффициента ранговой корреляции Спирмена (r).
РЕЗУЛЬТАТЫ
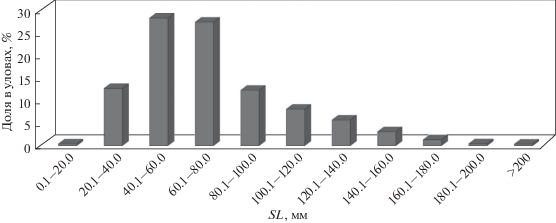
В уловах донных тралений в дельте Меконга были зарегистрированы рыбы 45 семейств, при этом таксономический облик ихтиофауны определяли представители 8 семейств, доля которых составляла по численности в совокупности более 90% вылова. Это представители семейств Polynemidae (26.5%, преимущественно Polynemus melanochir), Cynoglossidae (15.2%, Cynoglossus microlepis и C. feldmani), Ariidae (12.3%, в основном Arius maculatus, Osteogeneiosus militaris и Cephalocassis borneensis), а также ряд видов семейств Pangasiidae (11.1%), Engraulidae (9.1%), Akysidae (8.0%), Sciaenidae (6.7%) и Cyprinidae (5.0%). Размерный ряд уловов состоял из рыб со стандартной длиной 10.3–990.3 мм, при этом преобладали размерные группы 40–80 мм (рис. 2). Половозрелые особи составляли около 35%.
Крайние значения удельной численности в дельте Меконга колебались от 0.13 до 31.96 экз/100 м2 (табл. 2), т.е. отличались более чем в 300 раз, среднее значение составляло 3.4 экз/100 м2, для системы р. Тиен несколько больше – 4.0 экз/100 м2, для системы р. Хау, напротив, достигало лишь 3.1 экз/100 м2.
Таблица 2.
Вариации удельной численности рыб на различных участках дельты
| № участка* | N, экз/100 м2 | CV, % | W, г/100 м2 | CV, % | ||||
|---|---|---|---|---|---|---|---|---|
| среднее | min | max | среднее | min | max | |||
| 1 | 3.87 | 0.17 | 9.15 | 70.7 | 25.60 | 1.19 | 51.60 | 55.9 |
| 2 | 7.69 | 2.70 | 17.38 | 70.0 | 21.54 | 8.96 | 38.61 | 57.7 |
| 3 | 5.18 | 1.61 | 9.82 | 67.9 | 21.51 | 9.04 | 35.32 | 48.5 |
| 4 | 5.75 | 0.17 | 12.50 | 91.9 | 24.13 | 2.47 | 53.58 | 76.2 |
| 5 | 5.92 | 1.04 | 12.69 | 68.9 | 17.88 | 6.69 | 36.95 | 49.3 |
| 6 | 4.75 | 2.75 | 6.48 | 31.8 | 21.88 | 14.80 | 29.16 | 20.4 |
| 7 | 4.55 | 0.64 | 12.66 | 110.4 | 12.20 | 6.21 | 22.13 | 51.4 |
| 8 | 1.81 | 1.01 | 3.14 | 40.2 | 20.61 | 9.27 | 30.41 | 32.6 |
| 9 | 4.13 | 1.09 | 19.78 | 154.2 | 53.40 | 14.11 | 227.74 | 133.9 |
| 10 | 5.18 | 2.34 | 8.02 | 77.5 | 29.91 | 25.30 | 34.51 | 21.8 |
| 11 | 1.50 | 1.40 | 1.59 | 9.0 | 9.01 | 5.93 | 12.10 | 48.3 |
| 12 | 1.18 | 0.86 | 1.44 | 24.9 | 17.88 | 13.12 | 21.52 | 24.1 |
| 13 | 3.19 | 2.15 | 5.13 | 43.2 | 26.35 | 18.63 | 44.89 | 47.1 |
| 14 | 6.72 | 0.45 | 18.00 | 118.8 | 16.38 | 4.38 | 31.68 | 79.2 |
| 15 | 1.85 | 0.53 | 3.10 | 45.4 | 14.16 | 2.19 | 21.77 | 51.0 |
| 16 | 1.75 | 0.70 | 4.28 | 97.1 | 14.77 | 8.11 | 25.92 | 52.4 |
| 17 | 9.68 | 5.19 | 14.18 | 65.7 | 11.57 | 6.12 | 17.01 | 66.6 |
| 18 | 2.06 | 1.03 | 4.28 | 55.5 | 17.73 | 3.74 | 48.03 | 93.5 |
| 19 | 1.82 | 0.96 | 2.55 | 25.7 | 17.73 | 8.95 | 34.34 | 55.0 |
| 20 | 1.77 | 0.39 | 2.82 | 49.4 | 10.92 | 3.01 | 20.78 | 54.7 |
| 21 | 8.09 | 2.32 | 15.50 | 45.9 | 38.11 | 6.73 | 88.49 | 61.1 |
| 22 | 3.28 | 1.12 | 7.38 | 54.9 | 14.33 | 4.94 | 42.11 | 69.9 |
| 23 | 5.27 | 0.42 | 31.96 | 161.8 | 16.07 | 2.64 | 31.74 | 60.7 |
| 24 | 2.32 | 0.13 | 6.51 | 96.3 | 7.61 | 0.30 | 16.92 | 77.9 |
| 25 | 1.74 | 0.87 | 2.78 | 42.4 | 21.47 | 4.45 | 72.47 | 102.7 |
| 26 | 1.17 | 0.21 | 2.74 | 53.6 | 9.28 | 3.08 | 20.83 | 55.7 |
| 27 | 1.68 | 0.38 | 3.37 | 56.2 | 11.30 | 6.51 | 17.48 | 38.9 |
| 28 | 0.72 | 0.17 | 1.45 | 75.8 | 6.42 | 3.39 | 11.03 | 49.9 |
Результаты тралений более чем в 95% случаев находились в интервале до 10 экз/100 м2, преобладал класс значений 1.5–2.5 экз/100 м2. Большие показатели коэффициента вариации удельной численности (9.0–161.8%) свидетельствуют о значительной неоднородности выборок в пределах каждого участка. Аналогичные закономерности наблюдались в отношении удельной массы, пределы колебаний которой составляли 0.30–227.74 г/100 м2, среднее значение 18.8 г/100 м2, в р. Тиен – 21.2 г/100 м2, а в р. Хау – 15.6 г/100 м2. В целом распределение этого показателя в дельте было довольно равномерным (рис. 3а). Коэффициент вариации удельной массы менялся от 20.4 до 133.9% и для большинства участков был заметно ниже, чем для удельной численности.
Рис. 3.
Диаграммы размахов (медианы с квартилями) удельной численности (экз/100 м2) и биомассы (г/100 м2) рыб на участках дельты.
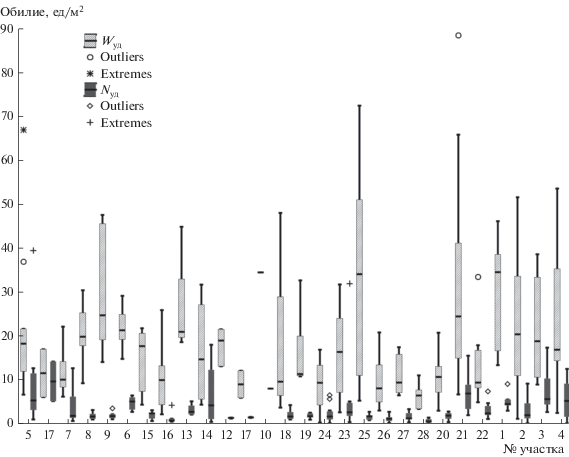
Минимальные значения средней плотности (менее 2 экз/100 м2) были приурочены к устьевым рукавам дельты, за исключением устьев Тиеу и Ба Лай. В реке Ба Лай плотина, расположенная на удалении около 17 км от устья, не дает возможности свободно перемещаться гидробионтам в процессе миграций, и у нижнего бьефа плотины скапливалась в значительных количествах молодь ариевых сомов Arius maculatus, представители этого семейства доминировали также в устье Тиеу, отличающемся крайне малым стоком [13, 14]. В реке Хау зона низких показателей обилия простиралась значительно дальше устьев и включала участок пресноводной зоны (ст. 24, 25), кроме того, район с низкой плотностью рыбного населения был отмечен в верхней части основного течения реки (ст. 20) (рис. 3б).
Около 5% тралений отличались чрезвычайно высокими показателями обилия. Они были сосредоточены преимущественно в системе р. Тиен, в меньшей степени – в основном течении р. Хау. При этом высокие значения достигались за счет наличия в р. Тиен в основном русле представителей семейств Polynemidae (Polynemus melanochir) и Akysidae (виды рода Pseudobagarius), в устьях – ариевых сомов Arius maculatus и Osteogeneiosus militaris, среди которых преобладали самцы, вынашивающие икру в ротовой полости. В основном русле р. Хау экстремально высокая численность рыб была обусловлена выловом больших количеств молоди пангасиевых сомов (семейство Pangasiidae, преимущественно Helicophagus leptorhinchus) либо левосторонних морских языков (семейство Cynoglossidae, виды Cynoglossus microlepis и C. feldmani). Очевидно, перечисленные рыбы при определенных условиях могут образовывать плотные скопления.
Показатели обилия рыбного населения в дельте Меконга были подвержены значительной вариабельности – пределы колебаний и средние величины заметно отличались не только в районах дельты, различных по своим гидрологическим и гидрохимическим параметрам (см. табл. 2), но и в пределах нескольких соседних тралений на одном участке, не имеющем заметных биоценотических отличий (рис. 4). Для выявления причин значительного разброса обилия была проведена оценка потенциального влияния на удельную численность и биомассу рыб различных факторов воздействия. К основным экологическим факторам относили соленость и температуру придонного слоя, глубину траления, количество донного растительного субстрата в трале, косвенно характеризующее характер биотопа. Кроме того, учитывали географическое положение, выраженное удалением станции от устья, а также распределение стока по рукавам и протокам [13]. К биотическим факторам относили показатели, характеризующие структуру доминирования в рыбных сообществах – доля представителей основных семейств в улове. Для оценки степени антропогенного воздействия анализировали влияние макропластика, поднятого тралом со дна. Кроме того, в оценке отклика принимали участие параметры тралений – скорость и продолжительность. При анализе были выявлены мультиколлинеарные факторы – температура и соленость придонного слоя, а также удаление от устья, причем наибольшая относительная сила влияния на отклик обнаруживалась у фактора удаление от устья.

При сравнении данных 2018 и 2019 гг. не отмечено достоверных различий в показателях обилия по удельной численности (H(1;n=220) = 0.001; p = 0.9801) и массе (H(1;n=220) = 3.59; p = 0.0582), что свидетельствует как о повторяемости результатов при использовании описанной методики сбора материала, так и об отсутствии существенных межгодовых колебаний удельной численности и биомассы рыб. При оценке влияния технических характеристик тралений на результаты отрицательная корреляция обнаружена со скоростью траления, однако коэффициент корреляции, а следовательно, и влияние этого параметра на удельную численность (rN = – 0.26, n = 220) и биомассу (rW = – 0.19, n = 220) рыб в уловах были весьма низкими.
Статистически значимая слабая положительная корреляция численности и биомассы рыб (р < < 0.05) была отмечена с таким фактором, как удаление от устья (rN = 0.39, rW = 0.16, n = 220). На удельную численность также оказывала влияние глубина траления (rN = 0.33, n = 220).
Для того чтобы исключить возможное разнонаправленное воздействие перечисленных факторов в районах с принципиально отличающимися экосистемами, их влияние было оценено отдельно в районах миксогалинных вод (эстуарном экотоне) и пресноводной части (основном течении рек). Отличия удельной численности рыб между основным течением и устьями были статистически значимы как в дельте в целом (H(1; n = 220) = 8.25; p = = 0.0041), так и в каждой из главных рек – Хау (H(1;n = 97) = 7.38; p = 0.0066) и Тиен (H(1; n = 123)= = 12.67; p = 0.0004). В основном течении рек наблюдалась взаимосвязь численности и биомассы при уровне значимости р < 0.05 с удалением от устья (rN = 0.37, rW = 0.33, n = 126). Помимо этого, удельная численность рыб зависела от глубины (rN = 0.32, n = 126) и очень слабо – от характера доминирования в рыбном сообществе (rN = 0.20, n = 126). В устьевых рукавах дельты некоторая отрицательная зависимость обилия наблюдалось от удельных показателей донных отложений как антропогенного (rN = –0.26, rW = –0.32, n = 94), так и природного (rN = –0.27, rW = –0.33, n = 94) происхождения. Здесь обилие также зависело от типа рыбного сообщества (rN = 0.25, rW = 0.21, n = 94). На численность рыб оказывало влияние также удаление от устья (rN = 0.35, n = 94), на удельную массу – объем стока (rW = –0.42, n = 94).
ОБСУЖДЕНИЕ
Оценка обилия рыб донно-придонных сообществ дельты Меконга ранее не проводилась, несмотря на очевидное снижение рыбных запасов на протяжении последних десятилетий. В результате проведенных комплексных ихтиологических исследований нами впервые для нижней части дельты Меконга получены данные о численности и особенностях пространственного распределения рыб. В настоящее время мы имеем дело с сообществами, в значительной степени деградировавшими под натиском хозяйственной деятельности человека, поэтому оценить произошедшие количественные изменения уже невозможно, однако полученные данные могут служить точкой отсчета для дальнейшего мониторинга ситуации. На всех участках дельты наблюдаются значительные флуктуации биоресурсов, что связано с ярко выраженным агрегированным типом распределения для некоторых наиболее массовых видов рыб. Очевидно, распределение их плотности формируется в результате сложного комплексного влияния многих факторов, в том числе взаимоисключающих, и к тому же подвержено значительной динамике в результате приливно-отливных явлений и процессов миграции. Для такой сложной экосистемы, как дельта крупной реки, чрезвычайно трудно учесть все факторы, воздействующие на биоту и определяющие закономерности ее распределения. Об этом свидетельствует зависимость обилия от удаления от устья, фактора, который, очевидно, включает как пространственное изменение гидрологических и гидрохимических показателей, так и другие аспекты воздействия.
В границах Вьетнама в дельте Меконга существуют две крупные экосистемы. Одна расположена в пресных водах, где основу сообществ составляют преимущественно пресноводные гидробионты. Другая – переходная эстуарная миксогалинная зона, локализованная в устьях дельты и простирающаяся вверх по течению на несколько десятков километров. Положение ее верхней границы подвержено динамичной пространственно-временной изменчивости и определяется на суточном масштабе высотой приливов и отливов, связанных с фазой луны, гидрометеорологическими условиями. Кроме того, в течение года она зависит от смены влажного и сухого сезонов, а также происходят межгодовые изменения, связанные с глобальной климатической цикличностью. В зависимости от распространения миксогалинных вод изменяются и структурные характеристики сообществ гидробионтов, формирующихся в этом экотоне. Основу сообществ составляют эвригалинные морские и пресноводные по своему происхождению животные, а также специфические солоноватоводные виды, обитающие в пределах этого экотона. Но особенности сообществ эстуарной и пресноводной областей дельты проявляются не только в таксономическом составе гидробионтов, но и в показателях обилия. При этом граница распространения более низких значений обилия (ст. 8, 9, 13, 15, 16, 18, 19, 27, 28) достаточно точно совпадала с границей миксогалинных вод.
Взаимосвязь обилия с характером рыбного сообщества определялась структурой доминирования, т.е. преобладанием видов какого-либо из восьми наиболее массовых семейств. В пресноводной части (основном течении) наблюдалась концентрация на глубине молоди пальцеперов, в основном P. melanochir, и циноглоссовых C. microlepis и C. feldmani. Скопления этих видов формировались на удалении от берегов и отсутствовали на мелководьях, что также определяло связь обилия с глубиной. В эстуарном экотоне к доминирующему комплексу в разных рукавах дельты принадлежали Coilia rebentischii (Engraulidae), A. maculatus (Ariidae) либо виды семейства Sciaenidae. В рукавах дельты, имеющих малый объем стока, концентрировались ариевые сомы, крупные и массивные рыбы, на течении обычно преобладали более мелкие коилии. При этом отрицательная корреляция с удельными показателями мусора и прочих отложений указывает на более тесную связь видов, населяющих миксогалинную зону, с донным субстратом.
Таким образом, при корреляционной оценке связей динамики обилия рыб с различными экологическими воздействиями природного и антропогенного характера было выяснено, что для прогнозирования количественных показателей рассмотренные экологические факторы мало пригодны, корреляция с исследованными факторами наблюдалась весьма слабая. Вероятно, причиной этого являлась сложность структурно-функциональной организации сообществ дельты и наличие неучтенных факторов воздействия. На данном этапе единственным надежным способом определения трендов изменения и флуктуаций обилия рыбного населения пока остаются тотальные учеты численности и биомассы рыб при проведении регулярных учетных съемок по единой методике.
Исследования выполнены по теме “Эколан Э-3.4”, раздел “Влияние климатических изменений и антропогенных факторов на экосистемы нижнего течения р. Меконг” и частично в рамках Государственного задания ФИЦ ИнБЮМ “Закономерности формирования и антропогенная трансформация биоразнообразия и биоресурсов Азово-Черноморского бассейна и других районов Мирового океана” (№ АААА-А18-118020890074-2).
Авторы выражают глубокую благодарность участникам экспедиций, научным сотрудникам Нгуен Тхе Май и Нго Хыу Тханг и многим коллегам из Южного отделения Тропцентра (г. Хошимин) за содействие в организации и проведении работ, чете рыбаков Чан Тхай Зыонг и Нгуен Тхи Чи Там за всестороннюю помощь во время экспедиций.
Список литературы
Poulsen A., Griffiths D., Nam S., Nguyen T.T. Capture-based aquaculture of Pangasiid catfishes and snakeheads in the Mekong River Basin // Capture-based aquaculture. Global Overview. FAO Fisheries Technical Paper № 508. Lovatelli A., Holthus P.F. Eds. Rome, 2008. P. 69–91.
Dugan P. Mainstream dams as barriers to fish migration: international learning and implications for the Mekong // Catch and Culture. 2008. V. 14. № 3. P. 9–15.
Mekong River Commission. Monitoring Fisheries in the Lower Mekong Basin. 2017. http://www.mrcmekong.org/ news-and-events/news/monitoring-fisheries-in-the-lower-mekong-basin/
Baran E., Guerin E., Nasielski J. Fish, sediment and dams in the Mekong. Penang, Malaysia: WorldFish, and CGIAR Research Program on Water, Land and Ecosystems (WLE), 2015. 108 p.
Bravard J.-P., Goichot M., Gaillot S. Geography of sand and gravel mining in the lower Mekong river: First survey and impact assessment // EchoGeo. 2013. № 26. P. 1–20. https://echogeo.revues.org/13659
Dudgeon D. The Ecology of Tropical Asian Rivers and Streams in Relation to Biodiversity Conservation // Annual Review of Ecology, Evolution, and Systematics. 2000. № 31. P. 195–229.
Dudgeon D. The contribution of scientific information to the conservation and management of freshwater biodiversity in tropical Asia // Hydrobiologia. 2003. № 500. P. 295–314.
Sabo J.L., Ruhi A., Holtgrieve G.W. et al. Designing river flows to improve food security futures in the Lower Mekong Basin // Science. 2017. № 358. P. 1–11.
Nguyen T. Hai Yen, Kengo Sunada, Satoru Oishi et al. The spatial distribution of fish species catches in relation to catchment and habitat features in the floodplain lot fisheries of Tonle Sap lake, Cambodia // J. of Fisheries and Aquatic Science. 2008. № 3. P. 213–227.
Halls A.S. Monitoring fish abundance and diversity in the Mekong Basin // Catch and Culture. 2008. V. 14. № 1. P. 24–25.
Ngor P.B., Theng L., Phem S., So N. Monitoring fish abundance and diversity around the Tonle Sap Lake // Catch and Culture. 2014. № 20. P. 20–27.
Болтачев А.Р., Карпова Е.П., Статкевич С.В. и др. Особенности количественного распределения рыб и десятиногих ракообразных в дельте реки Меконг в меженный период 2018 г. // Морской биологич. журн. 2018. Т. 3. № 4. С. 14–28. https://doi.org/10.21072/mbj.2018.03.4.02
Хо Чонг Тиен. Изучение изменений боковой приточности речных каналов в провинции Бенче, планирование решений по инженерной защите территорий. СРВ, Бенче: Научно-исследовательский институт водного хозяйства юга, 2001. 98 с.
Михайлов В.Н., Аракельянц А.Д. Особенности гидрологических и морфологических процессов в устьевой области р. Меконг // Водные ресурсы. 2010. Т. 37. № 3. С. 259–273.
Дополнительные материалы отсутствуют.