

Экология, 2021, № 6, стр. 420-429
Выживание ранних личинок налима (Lota lota L., 1758) в пойме Оби
А. Р. Копориков a, *, В. Д. Богданов a, **
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: Koporikov@mail.ru
** E-mail: Bogdanov@ipae.uran.ru
Поступила в редакцию 18.04.2021
После доработки 30.04.2021
Принята к публикации 05.05.2021
Аннотация
Проанализированы многолетние данные (2000–2017 гг.) учета численности личинок налима (Lota lota L., 1758) во время ската с мест инкубации икры на нерестовых притоках р. Оби (реки Войкар и Собь) и на нагуле в пойменной системе. Средние абсолютные численности нагульных личинок налима в р. Войкар – 12.4 млн экз, в р. Собь – 7.3 млн экз, средняя доля выживания перешедших на экзогенное питание личинок от числа вылупившихся составила 0.78 и 0.59% соответственно. Общая численность нагульных личинок определяется количеством молоди, скатившейся с нерестового притока: для р. Войкар – rSp = 0.741, p = 0.006, для р. Собь – rSp = 1.000, p < 0.001. Выявлена корреляционная зависимость выживания личинок налима в пойме от факторов среды, воздействующих на молодь до полного перехода на экзогенное питание. Например, корреляция выживания с длительностью затопления на уровне 7 м для р. Войкар – rSp = 0.732, p = 0.007; со среднесуточной температурой воды за 14 сут – rSp = 0.921, p < 0.001; с числом суток прогрева воды выше 8°С – rSp = = 0.780, p = 0.003; для р. Собь корреляция выживания со среднесуточной температурой воды за 14 сут – rSp = 0.900, p = 0.037. Уравнение линейной регрессии, связывающей выживание личинок со среднесуточной температурой воды за 14 сут, имеет следующий вид: y = 0.15x – 0.35, R2 = 0.95. Выдвинуто предположение, что главный фактор, влияющий на выживание нагульных личинок, – доступность стартового корма (фитопланктона), развитие которого определяется уровнем освещенности и температурой воды.
Живые организмы на ранних этапах жизни наиболее подвержены воздействию негативных факторов внешней среды. Оценка и прогноз успешности выживания особей в этом возрасте важны для понимания процессов устойчивости изучаемых популяций. Налим (Lota lota L., 1758) – циркумарктический пресноводный вид рыб с высокой плодовитостью (до 5.5 млн икринок) и относительно высокой продолжительностью жизни (в Обском бассейне может доживать до возраста 22 лет) и нерестящийся 3 и более раз на протяжении жизни [1, 2]. Исходя из такой стратегии размножения, можно предположить, что у налима должно наблюдаться очень низкое выживание особей, особенно на ранних этапах личиночного развития.
Известно, что при инкубации в искусственных условиях выживаемость икры налима составляет около 80% [3, 4]. В естественных водоемах этот показатель значительно ниже. Так, на притоках оз. Байкал доля покатной молоди от фонда отложенной икры составила около 0.15% [5]. В зависимости от условий выращивания выживание личинок налима в искусственных условиях во время перехода на экзогенное питание колеблется от 25 до 90% [6–9 и др.]. Литературные данные по выживанию личинок налима в естественных условиях от вылупления (первый этап личиночного развития) до полного перехода на экзогенное питание (третий этап) отсутствуют.
Цель настоящей работы – оценить уровень выживания личинок налима от вылупления до первичного распределения по местам нагула (до начала третьего личиночного этапа развития), определить влияние условий среды на выживаемость. Для решения поставленной задачи необходимо:
1) оценить численность покатных личинок на нерестовых притоках Войкар и Собь (бассейн нижней Оби);
2) определить экологическую плотность личинок налима в пойменной системе нерестового притока и в расположенной ниже по течению пойме р. Обь;
3) оценить площадь акватории, на которой покатные личинки налима из нерестового притока распределяются на нагул;
4) рассчитать абсолютную численность личинок, распределившихся на нагул в пойме;
5) определить выживание личинок налима на местах нагула, используя данные по численности покатной и нагульной молоди;
6) оценить влияние абиотических условий среды на выживаемость молоди.
МАТЕРИАЛ И МЕТОДИКА
Периодизация ранних личинок налима
В зарубежных и отечественных исследованиях оценка (количество и продолжительность) личиночных этапов развития налима различается. По данным отечественных авторов [5] продолжительность первого этапа личиночного развития, характеризующегося эндогенным питанием, составляет 3–4 сут. В зарубежной литературе [10] продолжительность первого этапа (yolk sac stage) оценивается в 8 сут. Отечественное определение второго личиночного этапа развития (смешанное питание, продолжительность с 5-х по 20-е сутки после вылупления) и начало третьего этапа (полное экзогенное питание) соответствует второму этапу развития (с 9-х по 26-е сутки с момента вылупления), принятому в зарубежной периодизации (preflexion stage). Различие в периодизации личиночных этапов развития налима может заключаться не столько в разных методах оценки возраста, сколько в разных условиях наблюдения за личинками (естественные или искусственные условия среды; температура; условия освещенности, проточности; доступность корма и т.д.).
Мы изучали личинок налима, находящихся в интервале развития от 0 и до 25-х суток с момента вылупления. В уральских нерестовых притоках к местам нагула с нерестилищ скатываются личинки, находящиеся на первом этапе развития (покатники). При попадании в пойменные водоемы личинки переходят на экзогенное питание (нагульные личинки). Под первичным распределением личинок налима по местам нагула мы подразумеваем распределение в пойме на этапе перехода на экзогенное питание (второй этап личиночного развития). В более позднем возрасте (при переходе к мальковому периоду) молодь налима начинает вторичное распределение на местах нагула.
Район исследования
Материал собирали на уральских нерестовых притоках нижней Оби – в реках Войкар, Собь и в расположенной ниже по течению пойме р. Обь (рис. 1, 2). Общая продолжительность сбора материала (с 2000 г. по 2017 г.) составила 14 лет (табл. 1, 2).
Рис. 1.
Карта-схема района работ: 1–5 – места отбора проб по нагулу личинок налима в пойме Оби ниже по течению от р. Войкар; 6–11 – ниже по течению от р. Собь; а – пойма нерестового притока р. Войкар; б – пойма нерестового притока р. Собь.
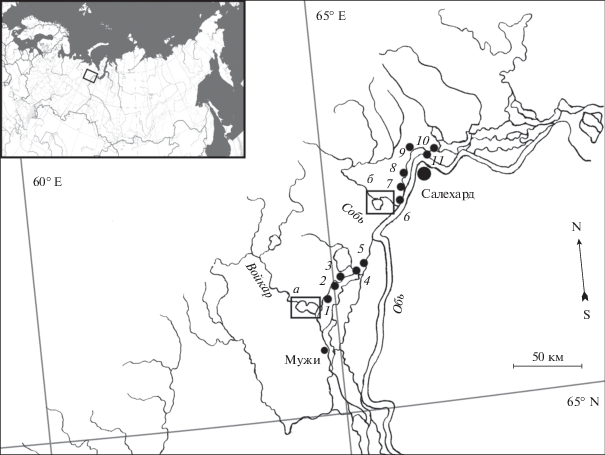
Рис. 2.
Карта-схема поймы нерестовых притоков р. Войкар (а) и р. Собь (б). Река Войкар: 1–11 – стандартные места отбора проб в литорали по нагулу личинок налима, скатившихся с р. Войкар, 12–15 – стандартные места отбора проб в пелагиали. Река Собь: 1, 2 – места отбора проб в литорали пойменного водоема Лор-Лох, 3–6 – в литорали пойменного водоема Пом-Лор, 7, 8 – в литорали пойменного водоема Сус-Пугол, 9–11 – в литорали пойменного водоема Урьях-Лор, 12, 13 – в литорали пойменного водоема Катравожский сор, 14 – в устье р. Собь.

Таблица 1.
Численность учтенных личинок налима в р. Войкар во время покатной миграции, на местах нагула в пойме и доля выживших нагульных личинок от общего числа покатников
| Год | Численность покатных личинок, млн экз. | Время с момента последнего пика численности покатных личинок налима на учетном створе притока и сбором нагульных личинок в пойме, сут | Средняя экологическая плотность личинок на местах нагула, экз/м2 | Рассчитанное число нагульных личинок, млн экз. | Доля выживших, % | |||
|---|---|---|---|---|---|---|---|---|
| в пойме притока | в пойме Оби | в пойме притока | в пойме Оби | сумма | ||||
| 2000 | 874 | 8 | 1.7 | 0.9 | 1.3 | 0.6 | 1.9 | 0.22 |
| 2001 | 2911 | 19 | 7.0 | 44.9 | 5.2 | 30.1 | 35.3 | 1.21 |
| 2003 | 530 | 15 | 3.7 | 1.0 | 2.8 | 0.6 | 3.4 | 0.65 |
| 2007 | 3483 | 15 | 6.6 | 28.0 | 4.9 | 18.8 | 23.7 | 0.68 |
| 2008 | 3506 | 9 | 9.4 | 28.5 | 7.0 | 19.1 | 26.1 | 0.74 |
| 2009 | 970 | 10 | 8.5 | 9.7 | 6.3 | 6.5 | 12.8 | 1.32 |
| 2011 | 2798 | 10 | 28.7 | 0.3 | 21.6 | 0.2 | 21.8 | 0.78 |
| 2012 | 450 | 19 | 3.9 | 0.5 | 3.0 | 0.3 | 3.3 | 0.73 |
| 2013 | 628 | 10 | 3.6 | 5.5 | 2.7 | 3.7 | 6.4 | 1.01 |
| 2014 | 738 | 9 | 6.9 | 0.0 | 5.1 | 0.0 | 5.1 | 0.70 |
| 2015 | 676 | 7 | 1.1 | 0.1 | 0.8 | 0.1 | 0.9 | 0.13 |
| 2017 | 637 | 5 | 3.1 | 7.8 | 2.3 | 5.2 | 7.5 | 1.18 |
| Среднее | 1517 | 11 | 7.0 | 10.6 | 5.3 | 7.1 | 12.4 | 0.78 |
| Стандартное отклонение | 1247 | 5 | 7.3 | 14.9 | 5.5 | 10.0 | 11.5 | 0.37 |
| Пределы | 450–3506 | 5–19 | 1.1–28.7 | 0.0–44.9 | 0.8–21.6 | 0.0–30.1 | 0.9–35.3 | 0.13–1.32 |
Таблица 2.
Численность учтенных личинок налима в р. Собь во время покатной миграции, на местах нагула в пойме и доля выживших нагульных личинок от общего числа покатников
| Год | Численность покатных личинок, млн экз. | Средняя экологическая плотность личинок на местах нагула, экз/м2 | Рассчитанное число нагульных личинок, млн экз. | Доля выживших, % | |||
|---|---|---|---|---|---|---|---|
| в пойме притока | в пойме Оби | в пойме притока | в пойме Оби | сумма | |||
| 2008 | 2039 | 0.051 | 8.5 | 0.039 | 11.1 | 11.1 | 0.55 |
| 2009 | 198 | 0.001 | 1.0 | 0.001 | 1.3 | 1.3 | 0.64 |
| 2010 | 414 | 0.000 | 2.2 | 0.000 | 2.8 | 2.8 | 0.69 |
| 2012 | 364 | 0.020 | 1.8 | 0.015 | 2.4 | 2.4 | 0.65 |
| 2016 | 4229 | 9.1* | – | – | 18.71 | 0.44 | |
| Среднее | 1449 | 0.018** | 3.4** | 0.014 | 4.4 | 7.3 | 0.59 |
| Стандартное отклонение | 1724 | 0.024** | 3.5** | 0.018 | 4.5 | 7.5 | 0.10 |
| Пределы | 198–4229 | 0.00–0.05** | 1.0–8.5** | 0.00–0.04 | 1.3–11.1 | 1.3–18.7 | 0.44–0.69 |
Нами не были рассмотрены отдельные годы (для р. Войкар – 2002, 2004–2006, 2010, 2016 гг., для р. Собь – 2011, 2013–2015 гг.) либо из-за полного отсутствия наблюдений по скату молоди (р. Собь, 2013 и 2014 гг.) или ее нагулу в пойменной системе (р. Войкар, 2016 г.), либо из-за эпизодичности этих наблюдений (р. Войкар, 2002, 2004–2006, 2010 гг.; р. Собь, 2011 и 2015 гг.).
Река Войкар (протяженность от истоков ручья Лёк-Хойла 225 км, площадь бассейна 8.1 тыс. км2) в отличие от других уральских нерестовых притоков в предгорной части своего течения протекает через крупное мелководное оз. Варчато [11], в котором обитает оседлая группировка налима и нагуливается его ранняя молодь. Учетный створ на реке оборудован в 36 км от устья (14 км выше пос. Вершина Войкар). В устьевой части реки расположен пойменный водоем Войкарский сор с площадью водного зеркала в период паводка 85 км2 и протяженностью периметра 55 км.
Река Собь (протяженность 185 км, площадь водосбора 5.9 тыс. км2) берет начало в ледниковом озере Полярного Урала на высоте 360 м над ур. м. Учетный створ оборудован в 38 км от устья (район ур. Тусигорт). В устье реки находится несколько пойменных водоемов, заполняемых во время паводка: Лор-Лох – площадь 1.8 км2, протяженность периметра 7.2 км; Пом-Лор – площадь 11.4 км2, протяженность периметра 16 км; Сус-Пугол – площадь 2.3 км2, протяженность периметра 8.2 км; Урьях-Лор – площадь 2.1 км2, протяженность периметра 7 км; Катравожский сор – площадь 3.1 км2, протяженность периметра 8.6 км.
Протяженность пойменной системы р. Обь на участке от устья р. Войкар до устья протоки Тоупуголпосл по руслу реки составляет 171 км. Ширина пойменной системы на этом участке изменяется от 50 км в районе устья р. Войкар до 6.5 км в районе г. Салехард.
Учет численности покатных личинок с мест инкубации икры
Изучение покатной миграции [12–14] личинок налима и учет их численности проводили на одних и тех же створах на протяжении всех лет наблюдений. Створы учета оборудованы в районе или ниже по течению от нижней границы нерестовых участков. Материал собирали на льду (до ледохода) и по открытой воде (после ледохода). Отбор проб дрифта по льду осуществляли с помощью ряда майн, прорубленных через равное расстояние перпендикулярно течению реки. Пробы дрифта по открытой воде отбирали в стандартных точках наблюдения с поставленной на якорь лодки. Применяемые для сбора ихтиопланктонные конусные сети были изготовлены из капронового мельничного газа № 21 и имели стандартную площадь входного отверстия 0.25 м2. Глубину отбора проб регулировали с помощью шестов (подо льдом) или системы груз–поплавок, прикрепляемой к конусной сети (по открытой воде). Скорость течения воды определяли при помощи измерителя скорости водного потока ГР-21М.
Абсолютную численность личинок, прошедших через учeтный створ, определяли по формуле
(1)
$N = \frac{{{{S}_{{\text{p}}}}{{V}_{{\text{p}}}}mT}}{{{{S}_{{{\text{лов}}}}}{{V}_{{{\text{лов}}}}}}},$Погрешность метода учета покатных личинок составляла от 30 до 40% [12].
Учет численности личинок в пойменной системе
Всю акваторию пойменных водоемов и водотоков условно разделили на зоны литорали и пелагиали. Под литоралью понимали зону пойменного затопления наземной или высшей водной растительности до ее нижней границы или аналогичную по глубине акваторию, на которой отсутствует водная растительность (песчано-галечные пляжи и т.д.). Толщу воды вне литоральной зоны мы определяем как пелагиаль.
В пойменной зоне литорали были выделены 4 типа биотопов:
заливной луг – мелководье с затопленной растительностью, грунт – мелкий песок, вода хорошо прогрета, течение практически отсутствует – обычно это затопленные сенокосные участки в пойме Оби;
обрывистый (подмывной) берег характеризуется высокой скоростью течения, большой глубиной, низкой температурой воды, растительность отсутствует;
непроточный песчано-галечный пляж – мелководье с полным или почти полным отсутствием затопленной наземной или высшей водной растительности, грунт песчано-галечный, вода хорошо прогревается, течение отсутствует – чаще всего это заливы вдали от основного русла водотока или в пойменном водоеме;
проточный песчано-галечный пляж – мелководье с полным или почти полным отсутствием затопленной наземной или высшей водной растительности, грунт песчано-галечный, течение присутствует и относительно высокое, вода плохо прогрета, близка к температуре основного течения – чаще всего это участки поймы, близкие к русловому участку водотока или проточного пойменного водоема.
Для отбора проб нагульной молоди налима использовали только участки литорали с полным или почти полным отсутствием течения (биотопы заливной луг и непроточный песчано-галечный пляж), так как ранее была установлена невозможность нагула молоди налима на проточных прибрежных участках [15].
Материал для оценки экологической плотности [16] нагульной молоди налима в зоне литорали отбирали сетью Киналева, изготовленной из капронового мельничного газа № 21 со сторонами 0.4 и 0.6 м. В пелагиали траление проводили ихтиопланктонной конусной сетью, изготовленной из капронового мельничного газа № 21 с площадью входного отверстия 0.25 м2. Глубину траления регулировали с помощью системы груз–поплавок, прикрепляемой к конусной сети. Скорость траления составляла около 1 м/с и контролировалась с помощью GPS навигатора, протяженность траления обычно составляла около 200 м. Траление в зоне пелагиали осуществляли для контроля завершения процесса первичного распределения ранней молоди налима в пойменной системе.
Учет экологической плотности личинок налима в литорали проводили после полного завершения покатной миграции на учетном створе и в пелагической зоне поймы. В среднем наблюдения в пойме проводили через 11 сут после последнего пика численности покатной молоди на учетном створе (крайние значения 5 и 19 сут). В расчет не принимали численность личинок в прибрежных биотопах, на которых в момент наблюдения были сильные прибойно-волновые явления, так как в это время молодь налима прижимается ко дну, прячется в укрытиях и недоступна для учета.
Расчет экологической плотности на прибрежных станциях определяли по формуле
где ρэкол – экологическая плотность молоди на исследуемой станции (экз/м2); n – численность пойманной молоди (экз); l – расстояние траления ловушкой (м); D – ширина ловушки (м).Расчет абсолютного количества личинок налима, распределившихся на нагул в пойменной системе нерестового притока и в расположенной ниже по течению пойме, определяли по формуле
(3)
$\begin{gathered} \Sigma n = ({{\rho }_{{{\text{Ср}}{\text{.Экол}}{\text{.Приток}}}}} \times {{S}_{{{\text{НагулПриток}}}}}) + \\ + \,\,({{\rho }_{{{\text{Ср}}{\text{.Экол}}{\text{.ПоймаОбь }}}}} \times {{S}_{{{\text{НагулПоймаОбь}}}}}), \\ \end{gathered} $(5)
${{S}_{{{\text{НагулПоймаОбь}}}}} = {{l}_{{{\text{ПоймаОбь}}}}} \times {{b}_{{{\text{ПоймаОбь}}}}}.$В анализе абиотических условий в районе акватории р. Войкар использованы данные, зафиксированные на гидропосту в пос. Мужи (Ямало-Ненецкий автономный округ, растояние от гидропоста до устья р. Войкар – 35.5 км): количество дней с момента затопления поймы до уровня 5, 6, 7 и 7.5 м (уровень затопления низкой поймы – 7.34 м); среднесуточные температуры воды за 5, 7, 10 и 14 сут; количество дней прогрева воды выше 5, 8 и 10°С. Все факторы рассматривали на отрезке времени до дня отбора проб по нагулу личинок в пойме.
Так как для р. Собь данные, полученные на гидропосту в пос. Мужи, имеют слабую актуальность (расстояние между гидропостом и устьем р. Собь 140 км), то для нерестового притока использованы только показатели среднесуточных температур воды.
Статистический анализ
Для характеристики параметров численности покатной и нагульной молоди применяли методы описательной статистики (средняя арифметическая, стандартное отклонение, пределы). Для оценки зависимости абсолютного количества нагульной молоди от количества молоди налима, родившейся в нерестовых притоках Войкар и Собь, а также зависимости выживания нагульной молоди от абиотических факторов среды (температура воды, уровень и продолжительность паводка) применяли коэффициент ранговой корреляции Спирмена (rSp) и проводили линейный регрессионный анализ. Статистическая обработка материала выполнена с использованием программ SPSS Statistics 14.0 [17] и Microsoft Office Excel 2003.
РЕЗУЛЬТАТЫ
Численность личинок, скатившихся с нерестовых притоков Войкар и Собь, а также их средняя экологическая плотность на местах нагула в пойменной системе нерестового притока и расположенной ниже по течению пойме р. Обь приведены в табл. 1 и 2. В связи с тем, что условия нагула в пойменной системе нерестового притока (куда молодь попадает в первую очередь) и пойме нижней Оби значительно различаются (площадь нагульных участков, температурный режим, количество доступных кормов и т.д.), расчеты проводили отдельно для каждого из этих нагульных участков. Ширина прибрежной зоны, пригодная для нагула молоди, в пойме нерестового притока в среднем нами определена в 20 м, для поймы Оби ниже устья нерестового притока (водотоки и временно затопляемые водоемы) – в среднем 10 м. По нашим расчетам, для поймы нерестового притока р. Войкар (Войкарский сор) протяженность пригодной для нагула береговой линии составила 37.5 км, для р. Собь – 38 км (суммарно для пойменных водоемов Лор-Лох, Пом-Лор, Сус-Пугол, Урьях-Лор и Катравожский сор).
Ранее [15] было показано, что распределение молоди налима в пойменной системе нижней Оби зависит от уровня водности, но, как правило, совмещение зон нагула для личинок, родившихся в разных уральских нерестовых притоках Оби, не происходит (удаление устьев нерестовых притоков Войкар и Собь 108 км). Большая часть нагульной молоди распределяется в 30 км от устья нерестового притока. Исключением могут быть годы с катастрофически низким уровнем паводка, когда низкая пойма не затапливается и молодь скатывается на большие расстояния по руслу водотока.
Личинки, родившиеся в р. Войкар, распределяются на участке поймы Оби от устья р. Войкар до слияния проток Кельчиас и Малая Обь. В среднем за годы наблюдений протяженность нагульных участков ниже устья р. Войкар составляет 67 км суммарно для западного и восточного берегов проток Горная и Малая Обь, для р. Собь подобная протяженность составила 130 км (восточный берег русла Оби, протоки Выл-Посл, Шома-Посл и др.) – от устья р. Собь до протоки Тоупуголпосл. Для р. Войкар рассчитанный показатель нагульной площади в зоне нерестового притока (SНагулПриток) составил 750 тыс. м2, в расположенной ниже по течению пойме Оби (S НагулПоймаОбь) – 670 тыс. м2; для р. Собь нагульная площадь в зоне нерестового притока (SНагулПриток) – 760 тыс. м2, в расположенной ниже по течению пойме Оби (SНагулПоймаОбь) – 1300 тыс. м2.
Используя данные по площади акватории, мы рассчитали абсолютные значения численности личинок налима на местах нагула и долю выжившей молоди от общего числа покатников (см. табл. 1, 2). Корреляционные расчеты показали, что суммарное количество нагульных личинок зависит от численности скатившейся с нерестового притока молоди: для р. Войкар – rSp = 0.741, p = 0.006, n = 12; для р. Собь – rSp = 1.000, p < 0.001, n = 5. В то же время доля выжившей молоди не зависит от численности покатников: для р. Войкар – rSp = = 0.189, p = 0.557, n = 12; для р. Собь – rSp = –0.600, p = 0.285, n = 5.
Показатели корреляции, описывающие выживание личинок налима в зависимости от абиотических условий среды, приведены в табл. 3. На рис. 3 показана зависимость выживания личинок, родившихся в р. Войкар, при их переходе на экзогенное питание от среднесуточных температур воды в пойменной системе за 14 сут: наблюдаемая линия тренда (сплошная линия) выживания нагульных личинок налима от среднесуточных температур воды за 14 сут, уравнение регрессии, величина достоверности аппроксимации. Цифры перед точками указывают время (в сут) с момента последнего пика численности покатников на учетном створе и до наблюдения в пойме личинок, перешедших на экзогенное питание.
Таблица 3.
Зависимость (коэффициент ранговой корреляции Спирмена rSp) выживания (доли выживших нагульных личинок от общего числа покатников) личинок налима от внешних условий среды во время нагула в р. Войкар (n = 12) и р. Собь (n = 5)
| Пара- метр |
Число суток с момента затопления пойменной водой уровня, м | Среднесуточные температуры воды за период, сут | Число суток прогрева воды выше | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 6 | 7 | 7.5 | 5 | 7 | 10 | 14 | 5°С | 8°С | 10°С | |
| Река Войкар | |||||||||||
| rSp | 0.600 | 0.674 | 0.732 | 0.403 | 0.839 | 0.937 | 0.902 | 0.921 | 0.640 | 0.780 | 0.858 |
| p | 0.039 | 0.016 | 0.007 | 0.194 | 0.001 | <0.001 | <0.001 | <0.001 | 0.025 | 0.003 | <0.001 |
| Река Собь | |||||||||||
| rSp | – | – | – | – | 0.900 | 0.900 | 0.900 | 0.900 | – | – | – |
| p | – | – | – | – | 0.037 | 0.037 | 0.037 | 0.037 | – | – | – |
Рис. 3.
Зависимость выживания личинок налима от среднесуточной температуры воды за 14 сут, предшествующих времени наблюдения в пойменной системе, р. Войкар, 2000–2017 гг. (пояснения приведены в тексте).
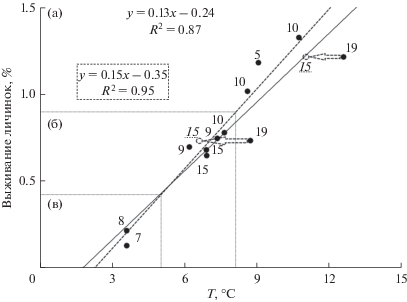
Данные по выживанию личинок налима в пойменной системе р. Войкар и расположенной ниже по течению пойме Оби позволяют выделить годы с разными условиями нагула (см. рис. 3): с неблагоприятными (пессимум) (2000 и 2015 гг.) – в (выживание личинок менее 0.4%), субоптимальными (2003, 2007, 2008, 2011, 2012, 2014 гг.) – б (от 0.4 до 0.9%) и с оптимальными (2001, 2009, 2013, 2017 гг.) – а (от 0.9% и выше). При неблагоприятных условиях (в) среднесуточная температура воды в пойме за 14 сут до момента наблюдения нагульных личинок не превышает 5°С, при субоптимальных (б) – изменяется от 5 до 8°С, при оптимальных (а) – более 8°С.
ОБСУЖДЕНИЕ
Результаты исследований по искусственному выращиванию ранней молоди налима в контролируемых условиях среды показали [9, 10 ] , что массовая смертность личинок наблюдается с 6-х по 15-е сутки с момента вылупления. На основании литературных данных мы предположили, что наблюдаемое выживание нагульных личинок в пойменной системе р. Войкар на 19-е сутки (2001, 2012 гг.) не имеет значимых отличий от тех, которые наблюдаются к 15-м суткам нагула. Среднесуточные температуры воды к 15-м суткам нагула имеют более низкие значения, так как ближе ко времени ледохода.
На рис. 3 внесенные изменения за 2001 и 2012 гг. изображены пунктиром (линия тренда, сдвиг точек выживания до 15-х суток, уравнение регрессии y = 0.15x – 0.35, величина достоверности аппроксимации R2 = 0.95). Полученная номограмма зависимости выживания ранних личинок налима на местах нагула в пойме от среднесуточных температур воды позволяет прогнозировать выживание по температурным данным.
При наличии двух пиков покатной миграции в р. Войкар, как это было в 2017 г., общая численность личинок налима, скатившихся во время второго пика, значительно больше (25% от общей численности покатных личинок – 161.5 млн экз.), чем вся нагульная молодь в пойменной системе (7.5 млн экз.). Такое соотношение скатившихся и нагульных личинок (см. табл. 3) свидетельствует о том, что массовая гибель личинок в пойме происходит до 5-х суток с момента вылупления.
Установлено, что массовая гибель личинок налима в естественной среде наблюдается гораздо раньше, чем в контролируемых условиях, и косвенно может указывать на то, что в природе они вылупляются на более поздних этапах эмбрионального развития из-за длительного инкубационного периода (около 150 сут). Как правило, в контролируемых искусственных условиях время инкубации икры налима происходит в более сжатые сроки [4, 18]. Личинки вылупляются физиологически менее развитыми, но с большими жировой каплей и желтком, которые позволяют дольше обходиться без пищи.
Анализ численности личинок налима в пойменной системе после перехода на экзогенное питание показывает, что в возрасте 11 ± 4 сут с момента вылупления выживание в естественных условиях в среднем составляет менее 1%.
В наших предыдущих исследованиях [19, 20] было показано, что на выживание ранних личинок сиговых рыб, находящихся в тех же биотопах, что и личинки налима, оказывает влияние время между затоплением пойменной системы и массовым заходом в нее личинок (r = 0.58, p < 0.05), что связано с концентрацией кормовых организмов в момент перехода на активное питание (корреляция с выживанием личинок пеляди r = 0.761). У личинок налима также наблюдается статистически значимая зависимость выживания от фактора “число суток с момента затопления пойменной водой уровня 5, 6 и 7 м” (см. табл. 3). В то же время можно заметить, что фактор температуры оказывает большее влияние на их выживание (см. табл. 3, рис. 3).
Известно, что стартовым кормом для личинок налима в течение первых трех суток после начала перехода на экзогенное питание является фитопланктон [6], затем они переходят на питание зоопланктонными организмами. Если в первые сутки перехода на экзогенное питание личинки лишены доступа к фитопланктону, то среди них наблюдается повышенная смертность.
Максимальная скорость роста фитопланктона достигается при комбинировании факторов температуры и освещения [21]. На момент наблюдений (конец мая–июнь) в исследуемом нами районе солнце фактически не заходит за линию горизонта на протяжении суток. Таким образом, фактор температуры напрямую влияет на темп увеличения концентрации фитопланктона в пойменной системе, что определяет выживаемость личинок налима.
Наблюдаемая корреляционная зависимость выживаемости личинок налима на втором этапе личиночного развития определяется именно доступностью кормовых организмов, концентрация которых в свою очередь обусловлена абиотическими условиями (в большей степени температурой воды и освещенностью). Следовательно, описываемые факторы среды (температура, время паводка) играют опосредованную роль и выступают своего рода факторами “ложной корреляции”, где скрытой переменной, по нашему мнению, выступает доступность корма.
Стабильная величина выживания личинок налима в пойме Оби в различные годы свидетельствует о существовании сбалансированной системы связей, позволяющих обеспечивать устойчивый уровень воспроизводства при различных условиях среды.
ВЫВОДЫ
1. Разработан метод оценки абсолютной численности нагульных личинок налима, перешедших на экзогенное питание. Сделан расчет абсолютной численности нагульной молоди в пойменной системе уральских нерестовых притоков Войкар и Собь и расположенной ниже по течению пойме Оби.
2. Впервые в естественных условиях определен уровень выживания ранних личинок налима при переходе на экзогенное питание. За первые два личиночных этапа развития в условиях поймы нижней Оби выживание за годы наблюдений изменялось от 0.13 до 1.32%, для р. Войкар выживание в среднем составляло 0.78%, для р. Собь – 0.59%.
3. Впервые за историю изучения вида показано, что, несмотря на огромную смертность (около 99%), абсолютная численность нагульной молоди в пойменной системе зависит от общего количества личинок, скатившихся с мест инкубации икры.
4. Выявлена зависимость выживания личинок налима от факторов среды: количества суток с момента затопления поймы; среднесуточных температур воды; количества суток прогрева воды выше 5°С.
5. Построено уравнение регрессии (номограмма) выживания личинок налима при переходе на экзогенное питание в зависимости от среднесуточных температур воды за 14 суток: y = 0.15x – 0.35, R2 = 0.95.
6. Все перечисленные факторы внешней среды оказывают опосредованное влияние на выживание личинок налима. Основным определяющим фактором выступает доступность кормовых организмов. На протяжении первых трех суток нагула наибольшее значение в качестве стартового корма имеет фитопланктон.
7. Установлено, что соотношение неблагоприятных, субоптимальных и оптимальных лет нагула для ранних личинок налима в пойменной системе р. Войкар и расположенной ниже по течению пойме Оби за годы наблюдений составило 2 : 6 : 4 лет соответственно, т.е. неблагоприятные условия для выживания ранних личинок налима формируются довольно редко.
8. Наиболее сложная трофическая ситуация для ранних личинок налима складывается в годы, когда личинки попадают в недавно затопленную пойму с низкой температурой воды.
9. Высокая смертность ранних личинок налима компенсируется очень большой плодовитостью самок, что обеспечивает успешное поддержание численности популяции в нестабильных условиях среды.
Работа выполнена в рамках Государственного задания ФГБУН ИЭРиЖ УрО РАН по теме “Состояние и динамика разнообразия животного мира Уральского региона, разработка научных основ его мониторинга и охраны” №АААА-А19-119031890085-3.
Авторы выражают благодарность д.б.н. М.Г. Головатину за многолетнюю помощь в сборе материала и к.б.н. Я.А. Кижеватову за предоставленный материал по скату и нагулу молоди налима в р. Собь.
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Тюльпанов М.А. Налим Обь-Иртышского бассейна (биолого-промысловый очерк). Томск: Изд-во Томск. гос. ун-та, 1966. 20 с.
Богданов В.Д., Копориков А.Р. Воспроизводство налима нижней Оби. Екатеринбург: УрО РАН, 2011. 156 с.
Володин В.М. Влияние температуры и pH на эмбриональное развитие налима // Бюл. Ин-та биол. водохранилищ АН СССР. 1960. № 7. С. 26–30.
Корляков К.А., Копориков А.Р., Новиков А.Л. Биотехнология искусственного выращивания налима (Lotidae) на Южном Урале и перспективы его использования в качестве биомелиоратора // Рыбн. хоз-во. 2013. № 1. С. 91–94.
Сорокин В.Н. Налим озера Байкал. Новосибирск: Наука, 1976. 144 с.
Harzevili A.S., De Charleroy D., Auwerx J. et al. Larval rearing of burbot (Lota lota L.) using Brachionus calyciflorus rotifer as starter food // J. of Appl. Ichthyol. 2003. V. 19. P. 84–87.
Żarski D., Sasinowski W., Kucharczyk D. et al. Mass initial rearing of burbot Lota lota (L.) larvae under controlled conditions // Pol. J. Natur. Sc. 2009. V. 24(1). P. 76–84. https://doi.org/10.2478/v10020-009-0007-9
Wocher H., Harsányi A., Schwarz F.J. Larviculture of burbot (Lota lota L.): larval rearing using Artemia and weaning onto dry feed // Aquacult. Res. 2012. V. 44. №. 1. P. 106–113. https://doi.org/10.1111/j.1365-2109.2011.03015.x
Palińska-Żarska K., Żarski D., Krejszeff S. et al. Dynamics of yolk sac and oil droplet utilization and behavioural aspects of swim bladder inflation in burbot, Lota lota L., larvae during the first days of life, under laboratory conditions // Aquacult. Int. 2014. V. 22. №. 1. P. 13–27. https://doi.org/10.1007/s10499-013-9663-4
Kupren K., Trąbska I., Żarski D. et al. Early development and allometric growth patterns in burbot Lota lota L. // Aquacult. Int. 2014. V. 22. №. 1. P. 29–39. https://doi.org/10.1007/s10499-013-9680-3
Кеммерих А.О. Гидрография Северного, Приполярного и Полярного Урала. М.: Изд-во АН СССР, 1961. 139 с.
Богданов В.Д. Изучение динамики численности и распределения личинок сиговых рыб реки Северной Сосьвы. Препринт. Свердловск: УНЦ АН СССР, 1987. 60 с.
Павлов Д.С., Назаров Д.Ю., Звездин А.О. и др. Покатная миграция ранних личинок европейской речной миноги Lampetra fluviatilis // Докл. РАН. 2014. Т. 459. № 2. С. 248–251. [Pavlov D.S., Nazarov D.Y., Zvezdin A.O. et al. Downstream migration of early larvae of the European river lamprey Lampetra fluviatilis // Dokl. Biol. Sci. 2014. V. 459. № 1. P. 344–347. doi 10.1134/S0012496614060039.]https://doi.org/10.7868/S0869565214320231
Pavlov D.S., Mikheev V.N. Downstream migration and mechanisms of dispersal of young fish in rivers // Can. J. Fish. Aquat. Sci. 2017. V. 74. №. 8. P. 1312–1323. https://doi.org/10.1139/cjfas-2016-0298
Копориков А.Р., Богданов В.Д. Особенности пространственно-биотопического распределения ранних личинок полупроходного налима (Lotidae) в пойме нижней Оби // Экология. 2011. № 4. С. 309–313. [Koporikov A.R., Bogdanov V.D. Spatial and biotopic distribution patterns of semianadromous burbot, Lota lota L. (Lotidae), Early Larvae in the Lower Ob Floodplain // Rus. J. of Ecology. 2011. V. 42. №. 4. P. 339–343. doi 10.1134/S1067413611040084]
Одум Ю. Экология. В 2-х т. М.: Мир, 1986. Т. 2. 376 с.
Pallant J. SPSS survival manual. A step by step guide to data analysis using SPSS for Windows. Maidenhead: Open University Press, 2007. 335 p.
Kucharczyk D., Mamcarz A., Kujawa R. et al. Burbot (Lota lota L.) reproduction in captivity: comparison of different methods // Elevage de la lotte (Lota lota): Recherche et perspectives. Nancy, 2004. P. 12–16.
Богданов В.Д., Богданова Е.Н. Использование факторов среды для оценки смертности личинок пеляди в условиях поймы Нижней Оби // Поведение рыб: Мат-лы IV Всерос. конф. с междунар. участием. Борок, 2010. С. 5–8.
Богданов В.Д., Богданова Е.Н. Трофический фактор во время перехода на активное питание личинок пеляди Coregonus peled (Gmelin) в нативном ареале // Экология. 2016. №. 2. С. 117–129. [Bogda-nov V.D., Bogdanova E.N. Trophic factor during the transition of peled (Coregonus peled Gmelin) larvae to active feeding in their native range // Rus. J. of Ecology. 2016. V. 47. № 2. P. 167–178. doi: 10.1134/S1067413616020041]https://doi.org/10.7868/S0367059716020049
Edwards K.F., Thomas M.K., Klausmeier C.A. et al. Phytoplankton growth and the interaction of light and temperature: A synthesis at the species and community level // Limnology and Oceanography. 2016. V. 61. № 4. P. 1232–1244. https://doi.org/10.1002/lno.10282
Дополнительные материалы отсутствуют.