

Экология, 2022, № 2, стр. 120-128
Новые данные о динамике растительности малоземельской тундры в голоцене на основе комплексного палеоботанического изучения озерно-болотных отложений в районе озера Голодная Губа
Е. Г. Лаптева a, *, О. М. Корона a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: lapteva@ipae.uran.ru
Поступила в редакцию 19.10.2021
После доработки 27.10.2021
Принята к публикации 03.11.2021
- EDN: ZBUAGU
- DOI: 10.31857/S0367059722020056
Аннотация
На основе палеоботанических данных, полученных из озерно-болотных отложений в районе оз. Голодная Губа, охарактеризованы стадии заболачивания существовавшего палеоводоема и сукцессии растительности Малоземельской тундры в течение голоцена. Установлено, что в раннем голоцене на эту территорию мигрировали древовидные березы и ель сибирская, которые в среднем голоцене вместе с лиственницей формировали таежные леса. В позднем голоцене леса сменились тундрами. Зарастание палеоводоема началось в раннем голоцене, и образовались низинные болота, которые к концу среднего голоцена через переходную стадию трансформировались в олиготрофные болота.
В настоящее время во всем мире уделяют большое внимание изучению динамики экосистем в течение голоцена – современной геологической эпохи, охватывающей последние ~11.5 тыс. лет. Экосистемы арктических и субарктических регионов наиболее чувствительны к климатическим изменениям. Изучение озерно-болотных отложений этих регионов с помощью палеоэкологических методов позволяет получить информацию о природно-климатических изменениях регионального и глобального характера в различные периоды голоцена.
Малоземельская тундра простирается на запад по левобережью низовья р. Печоры до Тиманского кряжа, на севере – по береговой линии Баренцева моря, а на юге – до р. Сула. Эта часть Российской Арктики в палеоэкологическом плане слабо изучена в отличие от Большеземельской тундры (правобережье р. Печоры до Полярного Урала), для которой реконструирована хронология изменений природной среды в голоцене на основе детальных палинологических летописей в сочетании с результатами палеокарпологического и радиоуглеродного анализов [1–8]. Для Малоземельской тундры детальные палеоботанические данные отсутствуют, и лишь из озерно-болотных отложений Тиманского кряжа получена серия палинологических записей [9, 10]. Однако для этой территории имеются сборы субфоссильной древесины и результаты ботанического определения макроостатков (хвои, коры, древесины) из торфяников, подкрепленные радиоуглеродными датировками [11–13].
Для реконструкции природной среды Малоземельской тундры в голоцене в 2013 г. было начато комплексное палеоботаническое изучение торфяных отложений в районе оз. Голодная Губа – самого большого пресноводного озера Ненецкого автономного округа, расположенного на территории заказника “Нижнепечорский”. Цель настоящей работы – описать и обсудить первые результаты спорово-пыльцевого и палеокарпологического анализов, которые получены при изучении отложений одного из торфяников около озера и дают представление о сукцессиях суходольной и водно-болотной растительности, зарастании и заболачивании палеоводоема.
РАЙОН, ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЯ
Район исследования – окрестности оз. Голодная Губа (67°51′40″ с.ш., 52°40′34″ в.д.), расположенного на северо-востоке Малоземельской тундры между дельтой р. Печоры на востоке и Ненецкой грядой на западе, в 26 км к северу от г. Нарьян-Мара в Заполярном районе Ненецкого автономного округа Архангельской области. Климат территории – субарктический, с длительной зимой и коротким нежарким летом. Вблизи г. Нарьян-Мар проходят изотермы января (–16°С) и июля (12°С). Средняя годовая температура в различных пунктах на окраинах Малоземельской тундры колеблется в пределах 2.2–3.6°С ниже нуля. Годовое количество осадков составляет около 600–800 мм [14]. Согласно геоботаническому районированию, оз. Голодная Губа находится в Малоземельско-Западно-Большеземельском округе Восточноевропейской подпровинции Европейско-Западносибирской тундровой провинции и приурочено к подзоне южных тундр, зональным типом растительности которой являются кустарниковые (ерниковые и ивняковые) тундры [15].
В торфянике на левом берегу оз. Голодная Губа в устье Черного ручья обследованы озерно-торфяные отложения, видимая мощность которых составила более 3 м. Из разреза отложений были отобраны сплошной колонкой 61 и 29 образцов для спорово-пыльцевого анализа и определения состава растительных макроостатков соответственно, а также образцы для радиоуглеродного анализа. Пробы, отобранные для палеоботанического изучения, в лабораторных условиях обрабатывали и анализировали по стандартным методикам [16, 17]. Обработку полученных данных и построение диаграмм выполняли с помощью пакета программ Tilia v. 2.0.41 [18]. На спорово-пыльцевой диаграмме доля пыльцы таксонов деревьев и кустарников, кустарничков и трав, а также спор высших споровых растений была рассчитана от общей суммы пыльцы древесных и травянистых растений, принятой за 100%. На диаграмме растительных макроостатков показано количество остатков того или иного таксона в каждом изученном образце объемом около 300–400 мл.
Возраст отложений установлен на основе серии радиоуглеродных дат (AMS-даты) (табл. 1), которые были получены в лаборатории AMS-датирования Национального Тайваньского Университета (the NTUAMS Lab, Department of Geosciences, National Taiwan University). Значения календарного возраста при калибровке радиоуглеродных дат рассчитаны с использованием калибровочной кривой CalPal_HULU в программе CalPal-2007 (quickcal2007 ver.1.6, http://www.calpal-online.de/) [19]. На основе полученной серии радиоуглеродных дат построена глубинно-возрастная модель в программе Bchron v. 4.7.3 [20]. Для определения хронологии изменений природной среды применяли схему периодизации голоцена Блитта-Сернандера, модифицированную для территории европейской части России Н.А. Хотинским [21].
Таблица 1.
Список радиоуглеродных дат, полученных с помощью ускорительной масс-спектрометрии для образцов торфа из разреза озерно-болотных отложений у оз. Голодная Губа (AMS-даты)
| Глубина, см | Лабораторный индекс и номер | Радиоуглеродный возраст, лет | Календарный возраст, л.н. (1σ) |
|---|---|---|---|
| 60–65 | NUTAMS-2266 | 1786 ± 34 | 1715 ± 65 |
| 125–130 | NUTAMS-2265 | 3287 ± 62 | 3530 ± 70 |
| 170–175 | NUTAMS-1182 | 6407 ± 32 | 7350 ± 50 |
| 260–265 | NUTAMS-2267 | 8661 ± 169 | 9775 ± 230 |
| 280–285 | NUTAMS-1183 | 9796 ± 60 | 11 220 ± 35 |
РЕЗУЛЬТАТЫ
Литологическое описание и радиоуглеродный анализ. Литологическое строение разреза представлено на рис. 1. Верхняя часть разреза образована слаборазложившимся торфом мощностью 175 см, в основании которого залегают стволы и ветки берез. Ниже расположен плотный сильно разложившийся торф, который подстилается с уровня 285 см сизыми песками. По образцам торфа изученного разреза получено 5 радиоуглеродных дат, которые хорошо согласуются между собой в пределах разреза (см. табл. 1).
Рис. 1.
Литологическое строение и модель роста отложений изученного разреза: 1 – дерн; 2 – торф слабо разложившийся, светло-коричневый; 3 – торф слабо разложившийся, коричневый; 4 – торф сильно разложившийся (до однородной массы), заиленный, темно-коричневый; 5 – пески сизые; 6 – стволы и ветки березы; 7 – 14СAMS даты, лет; 8 – 95%-ный доверительный интервал; ЛПЗ – локальные пыльцевые зоны, КМ – комплексы макроостатков. Пунктирная линия в модели – рассчитанный возраст на основе линейной экстраполяции.
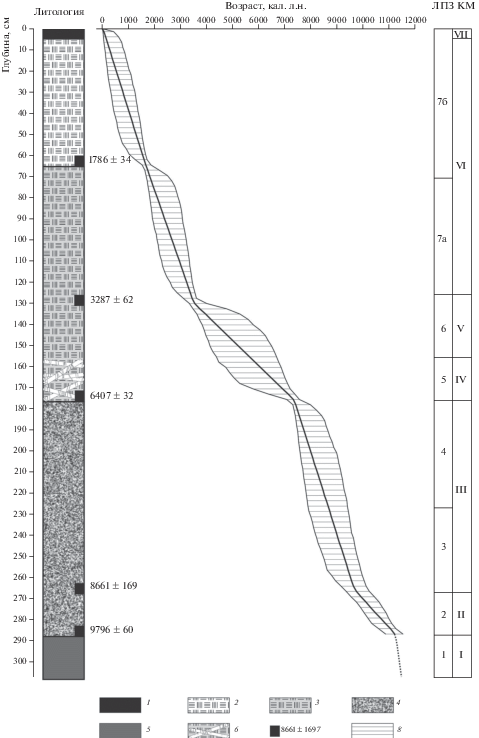
Построенная на основе полученных радиоуглеродных дат глубинно-возрастная модель в общем виде отражает скорость накопления органогенных отложений с раннего голоцена и до современности – примерно за ~11.2 тыс. кал. л.н. (см. рис. 1). Согласно расчетам, в интервале ~11.2–9.7 тыс. кал. л.н. скорость вертикального прироста отложений была низкая и составляла ~0.1 мм/год. Затем с 9.7 до 7.3 тыс. кал. л.н. скорость торфонакопления увеличилась до 0.4 мм/год. В интервале 7.3–3.5 тыс. кал. л.н. скорость торфонакопления снизилась до минимальной (~0.1 мм/год), а после 3.5 тыс. кал. л.н. снова возросла до 0.3–0.4 мм/год. Радиоуглеродные даты и возрастная модель накопления отложений использованы для определения календарного возраста границ локальных пыльцевых зон, комплексов растительных макроостатков и реконструированных фаз динамики растительности.
Спорово-пыльцевой и палеокарпологический анализы. По изменению таксономического разнообразия и содержания микро- и макроостатков растений ведущих таксонов выделено по семь локальных пыльцевых зон и комплексов макроостатков растений (рис. 2, 3), которые отражают процесс зарастания и заболачивания палеоводоема и сукцессионные изменения растительности.
Рис. 2.
Спорово-пыльцевая диаграмма отложений разреза торфяника у оз. Голодная Губа: точкой (•) обозначены находки устьиц; условные обозначения к литологии см. рис. 1. Дублирующая кривая обилия таксонов – увеличение в 10 раз.
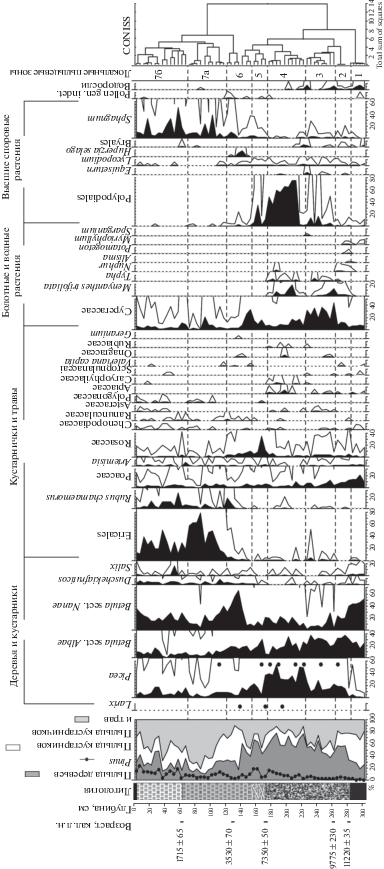
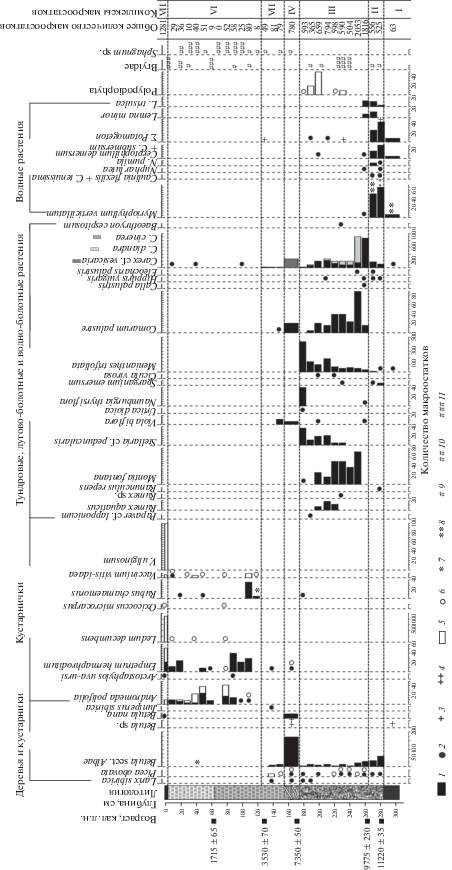
Рис. 3.
Диаграмма состава растительных макроостатков отложений разреза торфяника у оз. Голодная Губа: 1–4 – семена/плоды: 1 – обильные (более 5), 2 – единичные (менее 5), 3 – единичные фрагменты (менее 5), 4 – обильные фрагменты (более 5); 5–8 – листья/веточки: 5 – обильные (более 5), 6 – единичные (менее 5) , 7 – единичные фрагменты (менее 5), 8 – обильные фрагменты (более 5); 9–11 – остатки мхов: 9 – мало, 10 –много, 11 – обильно. Условные обозначения к литологии см. рис. 1.
ОБСУЖДЕНИЕ
По полученным палеоботаническим данным восстановлены следующие этапы заболачивания территории района исследования и сукцессии зональной и локальной растительности в течение голоцена.
После регрессии Баренцева моря в конце позднеледниковья и начале голоцена (13.7–10.0 тыс. кал. л.н.) в районе исследования началось формирование бассейна р. Печоры. Как такового оз. Голодная Губа не существовало, а по эрозионному котловану, который в настоящее время заполнен водами этого озера, протекала р. пра-Танюй [22].
Согласно палеоботаническим данным, полученным из подстилающего торфяник слоя сизых песков, в палеоводоеме со слабопроточной водой (старице) встречались роголистник погруженный (Ceratophyllum demersum L.), рдесты (Potamogeton berchtoldii Fieb. и P. pectinatus L.), уруть мутовчатая (Myriophyllum verticillatum L.) и водоросли (Pediastrum sp., Botryococcus sp., Cyclotella meneghiniana Kützing, Cosmarium sp. и др.). По берегам водоема произрастали осоки, в том числе осока пузырчатая (Carex cf. vesicaria L.), и вахта трехлистная (Menyanthes trifoliata L.). На плакоре доминировали ландшафты тундрового типа с ерниковыми зарослями и злаковники. Несмотря на то, что песчаные отложения в основании разреза не содержат в достаточном количестве органического вещества, необходимого для определения абсолютного возраста, на основании линейной экстраполяции радиоуглеродных дат из залегающих выше горизонтов торфа рассчитан вероятный возрастной интервал данного этапа около 11.5–11.2 кал. л.н. (см. рис. 1), хронологическое положение которого соответствует первой половине пребореального периода голоцена [21].
Около 11.2 тыс. кал. л.н., во второй половине пребореального периода, началось активное зарастание речных берегов и стариц. Стали формироваться низинные (эвтрофные) травяные болота вблизи палеоводоема. На них обильно произрастала осока пузырчатая, которая предпочитает обитать на низинных болотах, по берегам рек и озер, иногда в прибрежных мелководьях, на болотистых лугах, по окраинам стариц, в канавах, заболоченных кустарниках и лесах [23]. В близлежащем палеоводоеме еще активно развивалась водная макрофлора: роголистник погруженный, уруть мутовчатая, каулинии (Caulinia flexilis Willd. и C. tenuissima (A. Braun ex Magnus) Tzvelev), рдесты (Potamogeton alpinus Balb., P. berchtoldii Fieb., P. natans L., P. obtusifolius Mert. & W.D.J. Koch и P. praelongus Wulf), кубышки (Nuphar lutea (L.) Sm. и N. pumila (Timm) DC.) и ряски (Lemna minor L. и L. trisulca L.). О начале заболачивания территории во второй половине пребореального периода также свидетельствуют и ранее полученные из старичных отложений р. пра-Танюй датировки, определяющие возраст торфяных отложений около 11.0–10.5 тыс. кал. л.н. [22].
В это же время произошло проникновение древовидных берез в район исследования. В ЛПЗ 2 и КМ II, соответствующих возрасту 11.2–9.7 тыс. кал. л.н. (см. рис. 2 и 3), обнаружено большое количество крылаток и пыльцы Betula sect. Albae. Известно, что пыльца берез распространяется на сотни километров [17], в то время как основная масса семян рассеивается в пределах 200 м от источника, и лишь единичные семена могут разноситься ветром до 1 км [24]. Следовательно, обилие микро- и макроостатков Betula sect. Albae свидетельствует о произрастании древовидных берез в окрестностях палеоводоема во второй половине пребореального периода, которому соответствует полученная дата 11 220 ± 35 кал. л.н. (см. табл. 1). Ранее по стволу березы, обнаруженному в основании одного из торфяников оз. Голодная Губа, был установлен возраст около 10.5 тыс. кал. л.н. [22]. Южнее района исследования из разрезов, расположенных вверх по течению р. Печоры, также по стволам берез получена серия радиоуглеродных дат в диапазоне 10.1–11.8 тыс. кал. л.н. [1]. Соответственно новые полученные данные не противоречат мнению, что примерно с 11 тыс. кал. л.н. древовидные березы широко распространились на территории европейской части Российской Арктики [11, 13, 25]. Вместе с остатками древовидных берез обнаружены единичные семена ели сибирской (Picea obovata Ledeb.), а доля ее пыльцы достигает 15% (см. рис. 2 и 3). Семена ели преимущественно рассеиваются ветром в пределах 60–80 м от стены леса [26]. Находки семян ели являются доказательством произрастания данной породы в окрестностях существовавшего в пребореальном периоде палеоводоема. Датировки субфоссильной древесины или хвои ели, соответствующие этому временному интервалу, пока отсутствуют.
На произрастание ели в районе исследования в середине бореального периода указывают находки не только семян, но и хвоинок Picea obovata, которые маркируются датой 9775 ± 230 кал. л.н. (см. рис. 3), присутствуют макроостатки древовидных берез. В спорово-пыльцевых спектрах, соответствующих этому возрасту, пыльца ели и древовидных берез также обильна (см. рис. 2). Вероятно, в среднебореальное потепление [21] на водоразделах произрастали еловые и березовые леса с развитым моховым покровом. Ранее для субфоссильной древесины ели был определен возраст 9.8–9.4 тыс. кал. л.н. [13], что также соответствует среднебореальному времени. Согласно обилию пыльцы и находкам устьиц Picea в спорово-пыльцевых спектрах торфяников в дельте р. Печоры, туда ель мигрировала также во время среднебореального потепления около 10–9.9 тыс. кал. л.н. [6]. О широком распространении древовидных берез в высоких широтах в бореальном периоде свидетельствуют ранее полученные по их субфоссильным стволам датировки в диапазоне от 9.9 до 8.8 тыс. кал. л.н. для Малоземельской тундры и 10.5–8.6 тыс. кал. л.н. для Большеземельской тундры [11, 13, 25].
В это же время в районе существовавшего палеоводоема началась трансформация эвтрофных болот в мезотрофные. На данный процесс указывают высокое обилие растительных остатков таких мезотрофных растений, как осоки пузырчатая и двутычинковая (Carex diandra Schrank), вахта трехлистная и сабельник болотный (Comarum palustre L.), и почти полное исчезновение макроостатков и пыльцы макрофитов (см. рис. 2 и 3).
Согласно реконструкциям палеоклиматичских условий раннего голоцена в интервале 11.5–8.5 тыс. кал. л.н., полученным на основе пыльцевых данных и изучения растительных макроостатков из ряда разрезов Большеземельской тундры, средняя температура вегетационного периода составляла 15–17°С [27]. Вероятное превышение современной температуры вегетационного периода на 2–3°С в раннем голоцене способствовало расселению ели и древовидных берез в высоких широтах; сначала формировались редколесья, которые позднее – в среднем голоцене, приобрели таежный облик.
Широкое распространение ельников и березняков таежного типа с участием папоротников в напочвенном покрове было характерно для атлантического периода и первой половины суббореального периода с 8.6 до 3.5 тыс. кал. л.н., о чем свидетельствует обилие микро- и макроостатков древесных пород и папоротников в ЛПЗ 4, 5 и КМ III–V, сопоставимых с данным этапом (см. рис. 2 и 3). В интервале ~8.0(7.9)–4.3 тыс. кал. л.н. в окрестностях встречалась лиственница (Larix sibirica Ledeb.), поскольку найдены ее семена и устьица в отложениях, соответствующих данному времени. В образце из нижней части коричневого торфа, датируемым интервалом ~4.8–4.3 тыс. кал. л.н., обнаружены семена можжевельника (Juniperus sibirica Burgsd.). В настоящее время в окрестностях оз. Голодная Губа встречаются сообщества, не типичные для зональных тундр: островки из ели сибирской, березняки из Betula tortuosa Ledeb., заросли из Juniperus sibirica и папоротниковые сообщества, образованные щитовником шартским (Dryopteris carthusiana (Vill) H.P. Fuchs) [28].
О широком распространении лесных формаций в среднем голоцене на севере европейской части в Малоземельской и Большеземельской тундрах свидетельствуют многочисленные радиоуглеродные даты по субфоссильным стволам берез и ели [11, 13, 25]. Датировки субфоссильных стволов лиственницы в одном из разрезов торфяника Малоземельской тундры соответствуют интервалу 6.1–3.8 тыс. кал. л.н. [12], что соотносится с полученными палеоботаническими данными. Радиоуглеродная датировка ствола сосны (Pinus sp.) из торфяника в Малоземельской тундре, севернее района исследования, соответствует возрасту 3.7–3.5 тыс. кал. л.н. [12], но в изученных торфяных отложениях ее макроостатки не обнаружены. Хотя доля пыльцы Pinus в спорово-пыльцевых спектрах среднеголоценового возраста возросла до 10–20% с единичного содержания в раннем голоцене, вопрос о произрастании сосны в районе исследования остается открытым. Палеоботанические данные из разрезов в дельте р. Печоры и Большеземельской тундры также свидетельствуют о широком распространении таежных формаций вплоть до побережья Баренцева моря в среднем голоцене [2–4, 6].
Во второй половине атлантического периода произошло усиление болотообразовательного процесса, вероятно, за счет потепления и увеличения влагообеспеченности. Согласно реконструкциям палеоклимата по палинологическим данным из разрезов Большеземельской тундры [2, 29], в этот период годовая температура могла превышать современные температуры на 2–3°С, количество осадков было больше на 100 мм. В результате смены гидрологического режима мезотрофные болота в районе исследования стали трансформироваться в олиготрофные. Из подошвы слаборазложившегося коричневого торфа получена дата 7350 ± 50 кал. л.н. (см. табл. 1 и рис. 1), маркирующая обилие остатков осоки пепельно-серой (Carex cinerea Pollich), которая является растением-олиготрофом и предпочитает условия равномерного сыролугового и болотно-лугового увлажнения, плохо аэрируемые, небогатые питательными веществами оглеенные или торфяные почвы [23]. Остатки мезотрофных растений с этого уровня практически не встречаются, а появляются олиготрофные растения: водяника (Empetrum hermaphroditum Hagerup.) и сфагновые мхи (Sphagnum sp.).
Со второй половины суббореального периода происходит формирование тундрового комплекса. В образце торфа на глубине 125–130 см, имеющего дату 3530 ± 70 кал. л.н. (см. табл. 1 и рис. 1), и выше по разрезу резко снизилась доля пыльцы деревьев, возросло обилие кустарниковых берез (Betula sect. Nanae) и ольховника (Duschekia fruticosa (Rupr.) Pouzar); макроостатки древесных растений не обнаружены (см. рис. 2 и 3). Подобные изменения в растительном покрове отмечены и по данным из разрезов в дельте р. Печоры [3, 4, 6]. Активно формировались верховые болота с доминированием олиготрофных растений, преимущественно верескообразных кустарничков и сфагновых мхов, на что указывает обилие их микро- и макроостатков (см. рис. 2 и 3).
В настоящее время непосредственно над изученным торфяником в растительном покрове произрастают багульник (Ledum decumbens (Aiton) Lodd. ex Steud.), голубика (Vaccinium. uliginosum L.), морошка (Rubus chamaemorus L.), водяника, брусника (Vaccinium vitis-idaea L.), подбел узколистный (Andromeda polifolia L.), карликовая березка (Betula nana L.) и ряд других видов, макроостатки которых обильны в образце дерна (см. рис. 3). Несмотря на присутствие на водоразделах единичных еловых и березовых островков среди зональной тундровой растительности, макроостатки деревьев в образце дерна над торфяником не обнаружены. При изучении субрецентных проб дерна современной растительности оз. Голодная Губа макроостатки ели сибирской и березы извилистой обнаружены лишь в образцах непосредственно из соответствующих лесных островков или ближайших к ним не далее 100–200 м от деревьев. Субрецентные спорово-пыльцевые спектры отражают зональную тундровую растительность, лишь в палиноспектрах из еловых островков преобладает пыльца Picea. В спорово-пыльцевых спектрах березового островка из березы извилистой с карликовой березкой в кустарниковом ярусе наблюдается практически равное содержание пыльцевых зерен Betula sect. Albae и B. sect. Nanae.
ЗАКЛЮЧЕНИЕ
Таким образом, полученные палеоботанические данные характеризуют динамику растительности и процесс заболачивания в районе исследования за последние 11.2 тыс. лет. Во второй половине пребореального периода появились березы древовидные и ель сибирская и стали образовываться островки древесной растительности среди тундровых сообществ. Вблизи палеоводоема со слабо проточным режимом начался процесс зарастания, образовывались старицы и накапливались эвтрофные болотные отложения.
В бореальный период, вероятно, во время среднебореального потепления, ель сибирская наравне с древовидными березами широко распространилась, формируя редколесья, которые позднее приобрели таежный облик. Эвтрофные болота трансформировались в мезотрофные.
Леса таежного типа из ели сибирской, древовидной березы и лиственницы с участием папоротников в напочвенном покрове просуществовали до середины суббореального периода. Затем, после 3.5 тыс. кал. л.н., они деградировали, уступая место тундровым сообществам, которые в настоящее время являются зональным типом растительности района оз. Голодная Губа. Согласно анализу современной флоры района исследования, доля бореальных видов составляет более 48%, а арктических, включая гипоарктические, – менее 27% [28]. Возможно, обилие растений бореальной группы является следствием широкого распространения таежных лесов в среднем голоцене. В окрестностях палеоводоема в атлантическом периоде активизировался болотообразовательный процесс, вследствие чего мезотрофные болота стали модифицироваться в олиготрофные, которые активно формировались в позднем голоцене.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН № АААА-А19-119031890086-0. Авторы выражают благодарность Н.М. Николаевой (ГБУК “Ненецкий краеведческий музей”, г. Нарьян-Мар, НАО), А.С. Николаеву (г. Нарьян-Мар) и Л.И. Агафонову (ИЭРиЖ УрО РАН, г. Екатеринбург) за помощь в поиске, изучении разрезов торфяников и отборе образцов для спорово-пыльцевого и палеокарпологического анализов во время экспедиции на оз. Голодная Губа на территории заказника “Нижнепечорский” в 2013 г.
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Арсланов Х.А., Лавров А.С., Никифорова Л.Д. О стратиграфии, геохронологии и изменениях климата среднего и позднего плейстоцена и голоцена на северо-востоке Русской равнины // Плейстоценовые оледенения Восточно-Европейской равнины. М.: Наука, 1981. С 37–52.
Болиховская Н.С., Болиховский В.Ф., Климанов В.А. Климатические и криогенные факторы развития торфяников Европейского Северо-Востока СССР в голоцене // Палеоклиматы голоцена европейской территории СССР. М.: Ин-т географии АН СССР, 1988. С. 36–43.
Левашенко Д.В., Малясова Е.С. Климатический оптимум голоцена в дельте Печоры // Изв. РАН. Серия географ. 2007. № 4. С. 125–132.
Kaakinen A., Eronen M. Holocene pollen stratigraphy indicating climatic and tree-line changes derived from a pat section at Ortino, in the Pechora lowland, northern Russia // The Holocene. 2000. V. 10(4). P. 611–620. https://doi.org/10.1191/095968300677433659
Andreev A.A., Manley W.F., Ingόlfsson O. et al. Environmental changes on Yugorski peninsula, Kara Sea, Russia, during the last 12 800 radiocarbon years // Global and Planetary Change. 2001. V. 31. P. 81–91. https://doi.org/10.1016/S0921-8181(01)00123-0
Väliranta M., Kaakinen A., Kuhry P. Holocene climate and landscape evolution East of Pechora Delta, East-European Russian Arctic // Quaternary Research. 2003. V. 59. P. 335–344. https://doi.org/10.1016/S0033-5894(03)00041-3
Kultti S., Oksanen P., Väliranta M. Multiproxy record of Holocene environmental change in the Nenets region, East-European Russian arctic // J. of Canadian Earth Science. 2004. V. 41. P. 1141–1158. https://doi.org/10.1139/E04-058
Oksanen P.O., Kuhry P., Alekseeva R.N. Holocene development of the Rogovaya River peat plateau, East-European Russian arctic // The Holocene. 2001. V. 11. P. 25–40. https://doi.org/10.1191/095968301675477157
Paus A. Interpretative problems and sources of error related to pollen-analytical studies of the Holocene on the Timan ridge, western Pechora basin, northern Russia // Skrifter Arkeologisk Museum i Stavanger. 2000. V. 16. P. 111–126.
Paus A., Svendsen J.I., Matiouchkov A. Late Weichselian (Valdaian) and Holocene vegetation and environmental history of the northern Timan Ridge, European Arctic Russia // Quaternary Science Reviews. 2002. V. 22. P. 2285–2302. https://doi.org/10.1016/S0277-3791(03)00136-7
Кременецкий К.В., МакДональд Г.М., Галабала Р.О. и др. Об изменении северной границы ареалов некоторых видов деревьев и кустарников в голоцене // Бот. журн. 1996. Т. 81. № 4. С. 10–25.
Величко А.А., МакДональд Г.М., Кременецкий К.В. и др. Новые данные об изменении северной границы леса в голоцене // Докл. РАН. 1997. Т. 352. № 5. С. 690–692.
Kremenetski C.V., Sulerzhitsky L.D., Hantemirov R. Holocene history of the northern range limits of some trees and shrubs in Russia // Arctic and Alpine Research. 1998. V. 30. № 4. P. 317–333. https://doi.org/10.2307/1552004
Атлас Арктики. М.: Главное управление геодезии и картографии при Совете Министров СССР, 1985. 204 с.
Геоботаническое районирование Нечерноземья европейской части РСФСР. Л.: Наука, 1989. 64 с.
Никитин В.П. Палеокарпологический метод. Томск: Изд-во Томского ун-та, 1969. 89 с.
Гричук В.П., Заклинская Е.Д. Анализ ископаемой пыльцы и спор и его применение в палеогеографии. М.: Географгиз, 1948. 224 с.
Grimm E. Tilia software 2.0.2. Illinois State Museum Research and Collection Center Springfield, 2004.
Weninger B., Jris O., Danzeglocke U. CalPal-2007. Cologne Radiocarbon Calibration & Palaeoclimate Research Package. 2008. URL: http://www.calpal.de/ [дата посещения – 2015 г.]
Parnell A.C., Haslett J., Allen J.R.M. et al. A flexible approach to assessing synchroneity of past events using Bayesian reconstructions of sedimentation history // Quaternary Science Reviews. 2008. V. 27. № 19–20. P. 1872–1885. https://doi.org/10.1016/j.quascirev.2008.07.009
Хотинский Н.А., Алешинская З.В., Гуман М.А. и др. Новая схема периодизации ландшафтно-климатических изменений в голоцене // Изв. РАН. Серия географ. 1991. № 3. С. 30–42.
Лавров А.С., Потапенко Л.М. Неоплейстоцен Печорской низменности и Западного Притиманья (стратиграфия, палеогеография, хронология). М., 2012. 191 с.
Егорова Т.В. Осоки (Carex L.) России и сопредельных государств (в пределах бывшего СССР). Санкт-Петербург: Санкт-Петербургская гос. химико-фармацевтическая академия; Сент-Луис: Миссурийский ботанич. сад, 1999. 772 с.
Денисов С.А. Березняки Среднего Поволжья (биология, экология и комплексное хозяйство в березняках из В. pendula Roth и В. pubescens Ehrh.): Автореф. дис. … докт. с.-х. наук. Йошкар-Ола, 1999.
Binney H.A., Willis K.J., Edwards M.E. et al. The distribution of late-Quaternary woody taxa in northern Eurasia: evidence from a new macrofossil database // Quaternary Science Reviews. 2009. V. 28. P. 2445–2464. https://doi.org/10.1016/j.quascirev.2009.04.016
Евстигнеев О.И., Мурашев И.А., Коротков В.Н. Анемохория и дальность рассеивания семян деревьев восточноевропейских лесов // Лесоведение. 2017. № 1. С. 45–52.
Väliranta M., Salonen J.S., Heikkilä M. et al. Plant macrofossil evidence for an early onset of the Holocene summer thermal maximum in northernmost Europe // Nat. Commun. 2015. V. 6: 6809. https://doi.org/10.1038/ncomms7809
Николаева Н.М. Растительность и флора сосудистых растений окрестностей озера Голодная Губа // Озеро Голодная Губа. Заказник “Нижнепечорский”. Нарьян-Мар: ГБУК “Ненецкий краеведческий музей”, 2013. С. 15–21.
Andreev A.A., Klimanov V.A. Quantitative Holocene climatic reconstruction from Arctic Russia // J. of Paleolimnology. 2000. V. 24. P. 81–91.
Дополнительные материалы отсутствуют.