

Экология, 2022, № 5, стр. 334-344
Влажность почвы в урбанизированных местообитаниях с доминированием чужеродного Acer negundo
Д. И. Дубровин a, *, О. С. Рафикова a, Д. В. Веселкин a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: dubrovin_di@ipae.uran.ru
Поступила в редакцию 12.05.2022
После доработки 16.05.2022
Принята к публикации 16.05.2022
- EDN: HWLJVS
- DOI: 10.31857/S0367059722050031
Аннотация
Оценивали влажность почвы в урбанизированных местообитаниях с доминированием чужеродного (инвазивного) в Евразии вида дерева Acer negundo. Предполагали, что из-за густого полога листьев в местообитаниях с A. negundo почва может быть более влажной, чем в аналогичных условиях при доминировании других видов деревьев. Пробные площади в урбанизированных местообитаниях в г. Екатеринбурге (Средний Урал, Россия) были объединены в пары – с доминированием A. negundo или доминированием других видов деревьев. В паре местообитаний условия, кроме вида доминирующего дерева, были максимально схожи. В 2019–2021 гг. выполнили 170 регистраций объемной влажности верхних 5 см почвы – по 85 регистраций в местообитаниях с доминированием A. negundo и других древесных видов. Средняя объемная влажность почвы в местообитаниях с доминированием A. negundo (20.0 ± 0.9%) была ненамного, но значимо выше, чем в зарослях других видов деревьев (18.1 ± 0.8%). Близкими различия оказались в течение трех вегетационных периодов. Также различия влажности почвы между местообитаниями с доминированием A. negundo и других древесных видов были устойчивы при учете особенностей погодных условий и положения пробных площадей в рельефе. Таким образом, A. negundo является видом-трансформером в отношении режима влажности в местообитаниях-мишенях. Механизмы средопреобразующего воздействия A. negundo могут быть связаны с особенностями надземных органов, накоплением и разложением опада и подстилки, а также другими почвенными процессами.
Чужеродные растения могут оказывать средо-преобразующее воздействие на аборигенные экосистемы, проявляя свойства эдификаторов [1]. Такие растения, если они расширяют ареал и спектр местообитаний, относят к категории видов-трансформеров; их распространение вызывает наибольшие опасения [1]. Средопреобразующие воздействия, изменяющие условия или объем ресурсов в сообществах, учитываются в качестве ведущих объяснений в нескольких гипотезах успеха чужеродных видов: Global Competition Hypothesis, Sampling Hypothesis, Ideal Weed Hypothesis [2]. Средопреобразующие воздействия чужеродных растений могут реализовываться путем влияния на световой режим сообществ [3, 4], особенности круговорота питательных веществ [4], разные компоненты биоты [5–9].
При исследовании влажности как фактора среды в экологии растений преобладает представление о прямой причинно-следственной связи. При этом режим увлажнения (влажность почвы, осадки) рассматривается как фактор, влияющий как на видовой состав сообществ, так и на состояние отдельных особей [10]. Обратная задача – о влиянии растений на влажность почвы – рассматривается реже. Например, она возникает при изучении послерубочной трансформации лесных местообитаний [11], при обсуждении феномена гидравлического лифта [12], при создании засухоустойчивых сортов.
Чужеродные растения могут нуждаться в большем, меньшем или таком же количестве воды, как местные (обзоры [13, 14]). Некоторые из них способны более эффективно использовать почвенную влагу, что позволяет им быть более конкурентоспособными по сравнению с местными растениями в условиях недостатка влаги [15, 16]. В местообитаниях, занятых чужеродными растениями, наблюдалось и снижение [13, 17], и увеличение [13, 18, 19] доступной почвенной влаги по сравнению с сообществами аборигенных растений. Иногда трансформации режима увлажнения при доминировании чужеродных видов не было [13, 20, 21]. Косвенные доводы также свидетельст-вуют о возможности разнонаправленных изменений влажности почвы в зарослях чужеродных растений. Например, подтверждено, что чужеродные растения создают более густой полог листьев, чем аборигенные [3, 22–24]. Обусловленное этим затенение может способствовать росту влажности под пологом чужеродных видов [25], но, с другой стороны, большая масса листьев активно транспирирует, что может приводить к иссушению почвы [14, 26].
Подобная разнородность результатов обусловлена, по-видимому, функциональным разнообразием инвазивных растений и подчеркивает необходимость проверки гипотезы о режиме влажности почвы в отношении каждого конкретного вида. Цель настоящей работы – проверка гипотезы об особенностях влажности почвы в местообитаниях с доминированием инвазивного Acer negundo. Специальные исследования режима влажности в сообществах инвазивного дерева A. negundo неизвестны. Неоднократно установлено [27–29], что он создает большее затенение, чем местные деревья. Известно тяготение A. negundo к пониженным и пойменным местообитаниям в первичном [30] и вторичном [31] ареалах, хотя данный вид способен произрастать и в сухих плакорных местообитаниях в первичном [30] и вторичном [32] ареалах. На основании способности к высокому затеняющему воздействию мы предположили и проверили гипотезу, что в местообитаниях с A. negundo может быть более высокая влажность почвы, чем под другими деревьями в аналогичных условиях.
МАТЕРИАЛ И МЕТОДЫ
Район. Исследование выполнено в 2019–2021 гг. в черте и окрестностях г. Екатеринбурга: население – 1.5 млн человек, это административный центр Свердловской области, расположенный в южно-таежной подзоне бореально-лесной зоны. В растительном покрове преобладают сосновые (Pinus sylvestris L.) леса на дерново-подзолистых почвах и буроземах [33]. Климат умеренно континентальный; зима длительная и холодная с устойчивым снежным покровом; лето короткое. Среднегодовая температура +3.0°C, средняя температура января −12.6°C, июля +19.0°C. Среднегодовая сумма осадков 550–650 мм. Максимум осадков приходится на теплый сезон (май–август), в течение которого выпадает около 60–70% годовой суммы. Высота стояния солнца в истинный полдень в июне ─ июле составляет 52–56°C.
Acer negundo – дерево высотой до 20(25) м и диаметром ствола до 90(100) см. Первичный ареал охватывает Северную Америку от Скалистых гор до Атлантического побережья и от Канады до Флориды [30]. Интродуцирован в Европу в XVII в., в России известен со второй половины XVIII в. A. negundo входит в списки опасных инвазивных видов Европы [34], Беларуси [35], России [36, 37] и Свердловской области [38]. В первичном ареале растет в пойменных, мезотрофных лиственных и хвойных лесах, в дубовых редколесьях, в прериях, на полях и болотах [30]. Во вторичном ареале заселяет прибрежные фитоценозы, мезофитные и сосновые леса, занимает нарушенные и полуестественные местообитания, считается видом-трансформером [35, 36]. На Среднем Урале распространен повсеместно в рудеральных и других антропогенно преобразованных местообитаниях, активно возобновляется в городских лесопарках [39]. В зарослях A. negundo снижается α- и γ-разнообразие трав [40, 41], а высокая сомкнутость его крон – один из механизмов влияния на напочвенный покров [42].
Пробные площади. Общий дизайн исследования – блочное рандомизированное наблюдение или пассивный эксперимент. Для измерения влажности почвы подобраны объединенные в пары пробные площади (ПП). Блоком (рис. 1) считался участок, т.е. территория, включающая пару ПП с доминированием Acer negundo (An+) и контрольную к ней с доминированием другого вида дерева (An–). Доминирующим считали вид, общее число стволов которого на ПП было больше числа стволов любого другого вида. В одном блоке ПП An+ и An– были одинакового размера – 10 × 10 или 20 × 20 м – и находились в схожих условиях: а) они были однородны по положению в рельефе и типу ландшафта; б) располагались на расстоянии не более 0.4 км друг от друга; в) были близки по степени антропогенной трансформированности. Еще одним важным критерием подбора ПП An+ и An– в одном блоке были близкие значения сомкнутости крон деревьев. Таким образом, условия на ПП An+ и An– в одном блоке были сходными и различались только по доминанту древостоя. На ПП An– доминировали Malus baccata (L.) Borkh, Prunus padus L., Pinus sylvestris L., Quercus robur L., Salix alba L., Sorbus aucuparia L., Tilia cordata Mill., Ulmus laevis Pall.
Рис. 1.
Схема размещения пробных площадей (квадраты) внутри блоков или участков (круги); An+ – площади в зарослях A. negundo, An– – площади в зарослях других видов деревьев.
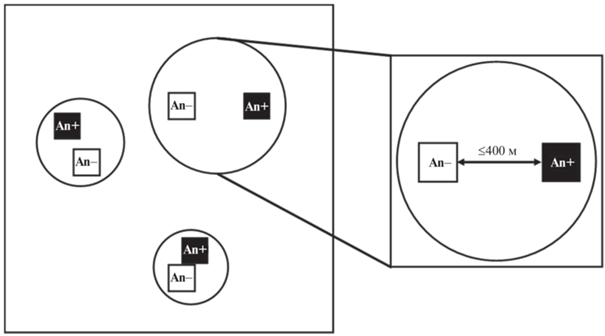
Положение ПП в рельефе характеризовали высотой над уровнем моря (м), которую измеряли в программе SASPlanet (1.0, SAS Group, 2014). Высоты изменялись от 226 до 309 м над ур. м.
Схемы измерения объемной влажности верхних 5 см почвы (SWC). Измерения SWC верхних 5 см минеральной части почвы выполняли портативным влагомером HH2 Moisturemeter (Великобритания; Delta-T Devices). Перед измерением удаляли опад и подстилку, сенсор вводили в верхний слой минеральной части почвы вертикально вниз и регистрировали SWC (в %). Измерения SWC выполняли в ходе 1–3 туров каждый вегетационный сезон (табл. 1). В одном блоке измерения SWC всегда выполняли в один день, чтобы в промежуток времени между измерениями на ПП An+ и An– не выпадало осадков. Измерения SWC проводили по двум схемам:
Таблица 1.
Даты и количество измерений объемной влажности почвы
| Дата | Количество площадей | Всего измерений | |
|---|---|---|---|
| An– | An+ | ||
| Схема I : удаление опада и подстилки непосредственно перед измерением SWC | |||
| 22 июля–5 августа 2019 г. | 11 | 11 | 550 |
| 20–22 августа 2019 г. | 11 | 11 | 550 |
| 22–26 июня 2020 г. | 9 | 9 | 450 |
| 24 июля–3 августа 2020 г. | 19 | 19 | 950 |
| 25 июля–4 августа 2021 г. | 11 | 11 | 550 |
| Схема II : удаление опада и подстилки, рыхление верхних 3–5 см почвы перед первым в вегетационном сезоне туром измерения SWC | |||
| 10 июля 2019 г. | 5 | 5 | 240 |
| 18 июля 2019 г. | 5 | 5 | 240 |
| 22 августа 2019 г. | 5 | 5 | 240 |
| 15 июля 2021 г. | 3 | 3 | 108 |
| 25 июля 2021 г. | 3 | 3 | 108 |
| 3 августа 2021 г. | 3 | 3 | 108 |
| Всего | 85 | 85 | 4094 |
Схема I – измерение SWC в 25 точках, случайно размещенных в пределах ПП 10 × 10 м или ПП 20 × 20 м с удалением опада и подстилки непосредственно перед измерением SWC. Схема I реализована в 2019–2021 гг.
Схема II – измерение SWC в 18–24 точках на ПП 10 × 10 м, приуроченных к фиксированным площадочкам размером 0.5 × 0.5 м, на которых предварительно (один раз в начале вегетационного сезона) были удалены опад и подстилка и произведено рыхление верхних 3–5 см почвы. Схема II реализована в 2019 и 2021 гг.
Сомкнутость крон. На каждой ПП в 2019 и 2020 гг. цифровым фотоаппаратом Lumix DMC-FP2 (ПЗС-датчик – 1/2.5''/10.3 млн пикселей/первичный цветовой фильтр; разрешение фото – 3648 × 2736 пикселей) в середине июля в 10 случайных точках делали по одному цветному снимку полога листьев вертикально вверх (с высоты 0.8–1.2 м) – всего 680 снимков. Снимки конвертировали в бинарные в программе Adobe Photoshop 11.0 (Adobe System Inc., 2008) таким образом, что черные пиксели соответствовали естественным преградам солнечному свету, а белые пиксели – открытому небу. Анализ сомкнутости крон произведен в программе Matlab R2018b (9.5.0.944444, The MathWorks Inc., 2018) с помощью оригинальной программы [42], которая оценивала отношение черных пикселей к общему числу пикселей на изображении и среднюю для ПП величину этого параметра, которую анализировали как сомкнутость крон.
Погодные условия вегетационных сезонов. Среднесуточные температуры воздуха и суммы осадков с 1 мая по 31 августа 2019–2021 гг. извлекли с ресурса http://www.pogodaiklimat.ru [43]. Вегетационный сезон 2019 г. был прохладным и влажным (табл. 2), сезон 2020 г. ─ средним по температурным условиям с достаточно влажным июнем и сухим июлем, а сезон 2021 г. ─ самым теплым со средним количеством осадков.
Таблица 2.
Среднесуточные температуры воздуха и суммы осадков в вегетационные сезоны 2019–2021 гг.
| Месяц | Среднесуточная температура воздуха, °C | Сумма осадков, мм | ||||
|---|---|---|---|---|---|---|
| 2019 г. | 2020 г. | 2021 г. | 2019 г. | 2020 г. | 2021 г. | |
| Май | 7.6 | 9.0 | 11.8 | 39.0 | 13.3 | 15.5 |
| Июнь | 11.1 | 9.9 | 13.1 | 49.0 | 72.9 | 51.9 |
| Июль | 14.8 | 16.8 | 14.5 | 126.1 | 20.7 | 84.5 |
| Август | 12.2 | 12.9 | 14.8 | 94.3 | 156.5 | 79.4 |
Характеристикой текущих погодных условий в период, непосредственно предшествующий измерению влажности почвы, была сумма осадков (мм), выпавших в течение 10 сут до измерения влажности почвы. Период в 10 сут выбрали на основании предварительного анализа, когда суммы осадков рассчитывали за период от 1 до 30 сут до измерения влажности почвы. Сумма осадков за 10 сут показала наиболее сильную регрессионную связь с влажностью почвы.
Анализ данных. Единицей при проведении статистического анализа было среднее значение SWC (или другого параметра) на одной ПП в конкретный тур измерений, т.е. n = 2 × 85 = 170. Для сравнения значений SWC на парах ПП An+ и An– при устраненной изменчивости, обусловленной всеми другими факторами, использовали t-критерий Стьюдента для попарно связанных переменных. Связь погодных условий и особенностей местообитаний с SWC оценивали с помощью общих (GLM) и смешанных (LMM) линейных моделей. В моделях LMM случайным фактором был блок или участок. Значения признаков, выраженных в долях (объемная влажность почвы, сомкнутость крон), анализировали после предварительного арксинус-преобразования. Значения сумм осадков за 10 сут – после предварительного логарифмирования, однако в тексте, таблицах и на рисунках использованы нетрансформированные значения признаков. Через символ ± приведена стандартная ошибка. Расчеты выполнены в пакетах JMP 10.0.0 (SAS Institute Inc., USA, 2012) и STATISTICA 10.0 (StatSoft, USA).
РЕЗУЛЬТАТЫ
Средняя арифметическая во всем массиве полученных за три вегетационных сезона 170 значений объемной влажности почвы составила 19.1 ± 0.6%, медиана – 17.9%, коэффициент вариации – 40.2%.
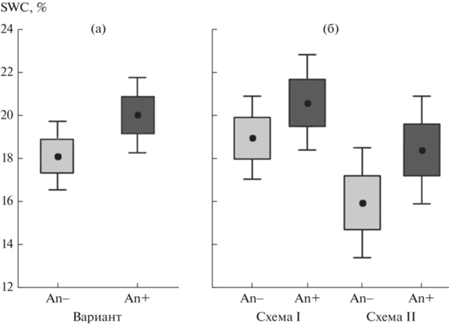
Влажность почвы в местообитаниях An– и An+. Влажность почвы зависит от большого числа факторов среды: от условий местообитаний и условий вегетационного сезона, например от режима осадков. На первом этапе, чтобы установить особенности SWC в зависимости от основного интересовавшего нас фактора – доминанта древесного яруса – выполнили анализ SWC с помощью t-критерия Стьюдента для связанных переменных. Такой подход позволил оценить различия значений SWC в вариантах местообитаний An– и An+ при отброшенной изменчивости, связанной с другими факторами, влияние которых мы предполагали или не предполагали. Средняя объемная влажность почвы в местообитаниях с доминированием A. negundo была значимо выше, чем в зарослях других видов деревьев (рис. 2а): 18.1 ± 0.8% в варианте An– и 20.0 ± 0.9% в варианте An+ (t = –1.87, dF = 85, P = 0.0001).
Рис. 2.
Объемная влажность почвы (SWC; среднее, ±SE, ±95%-ный доверительный интервал) объединенной выборки (а) и для разных схем измерения (б) в зарослях разных видов деревьев (An–; светло-серые фигуры) и зарослях Acer negundo (An+; темно-серые фигуры).
Основные факторы, влияющие на влажность почвы. Чтобы оценить вклад доминанта древостоя в общее варьирование SWC, рассчитали GLM-модель, включающую дискретные и континуальные предикторы: 1) вариант наблюдения – в древостое доминирует A. negundo или другой вид; 2) год измерения – подразумевая, что этот предиктор суммирует погодные и фенологические особенности вегетационного сезона; 3) схема измерения SWC – с удалением опада/подстилки непосредственно перед каждым измерением или с удалением опада/подстилки один раз в сезон перед первым туром измерений; 4) высота местности над уровнем моря; 5) сумма осадков за 10 сут, предшествовавших измерению SWC. Оценили все двухфакторные взаимодействия, кроме взаимодействия “год × схема измерения” (табл. 3).
Таблица 3.
Значимость влияния пяти предикторов, включая фактор “вариант”, и их взаимодействий в GLM- и LMM-моделях (со случайным фактором “участок”), объясняющих изменчивость объемной влажности почвы в урбанизированных местообитаниях на протяжении вегетационных сезонов 2019–2021 гг.
| № | Сочетание предикторов | dF | GLM | LMM | ||
|---|---|---|---|---|---|---|
| F | P | F | P | |||
| 1 | Вариант | 1 | 6.08 | 0.0148 | 8.55 | 0.0040 |
| 2 | Год | 2 | 83.99 | <0.0001 | 84.52 | <0.0001 |
| 3 | Схема измерения | 1 | 47.91 | <0.0001 | 20.29 | <0.0001 |
| 4 | Высота местности | 1 | 43.60 | <0.0001 | 11.81 | 0.0026 |
| 5 | Осадки за 10 сут | 1 | 48.33 | <0.0001 | 62.60 | <0.0001 |
| 6 | Вариант × год | 2 | 0.79 | 0.4579 | 1.03 | 0.3581 |
| 7 | Вариант × схема измерения | 1 | 0.04 | 0.8457 | 0.02 | 0.8912 |
| 8 | Вариант × высота местности | 1 | 0.35 | 0.5550 | 0.55 | 0.4602 |
| 9 | Вариант × осадки за 10 сут | 1 | 0.14 | 0.7066 | 0.18 | 0.6694 |
| 10 | Год × высота местности | 2 | 1.20 | 0.3028 | 0.43 | 0.6514 |
| 11 | Год × осадки за 10 сут | 2 | 2.82 | 0.0626 | 2.17 | 0.1176 |
| 12 | Схема измерения × высота местности | 1 | 0.52 | 0.4706 | 0.54 | 0.4667 |
| 13 | Схема измерения × осадки за 10 сут | 1 | 7.54 | 0.0068 | 10.67 | 0.0014 |
| 14 | Осадки за 10 сут × высота местности | 1 | 0.93 | 0.3376 | 1.98 | 0.1618 |
| R2 | 0.732 | 0.809 | ||||
| $R_{{{\text{Adj}}}}^{2}$ | 0.700 | 0.786 | ||||
Общее качество GLM-модели было высоким – использованная комбинация предикторов объяснила около 73% общей изменчивости SWC. Все главные эффекты были статистически значимыми. Следовательно, объемная влажность почвы различалась в разные годы, а также в зависимости от варианта наблюдения, схемы измерения, высоты местности и суммы недавно выпавших осадков. Только одно взаимодействие факторов из 9 рассчитанных было статистически значимым. При дополнительном учете в LMM-модели в качестве случайного фактора участка, на котором размещалась пара площадей An– и An+, качество объяснения изменчивости SWC улучшилось. При этом все эффекты, значимые в GLM, значимы и в LMM.
Средние значения SWC в разные годы составили: 2019 г. – 24.4 ± 0.8%; 2020 г. – 14.0 ± 0.6%; 2021 г. – 16.3 ± 0.9%. При удалении опада/подстилки с участка один раз в сезон перед первым измерением (схема II) значения SWC были в среднем низкими (17.2 ± 0.9%), а при удалении опада/подстилки непосредственно перед каждым измерением (схема I) – более высокими (19.8 ± 0.7%).
Хотя взаимодействие факторов “год × схема измерения” оценить не удалось, более высокие значения SWC при удалении подстилки непосредственно перед измерением влажности хорошо заметны и в 2019, и в 2021 гг.: в 2019 г. при схеме I ─ SWC = 27.9 ± 0.9%, при схеме II – 19.3 ± ± 1.1%; в 2021 г. при схеме I ─ SWC = 18.4 ± 1.2%, при схеме II ─ 13.7 ± 1.0%. Закономерность более высоких значений SWC при удалении опада/подстилки непосредственно перед измерением влажности просматривается также при сравнении вариантов An– и An+ (рис. 2б).
С увеличением высоты местности значения SWC закономерно уменьшались независимо от древесного доминанта (рис. 3а). Различия наклонов линий регрессии зависимостей “высота местности–SWC” в вариантах An– и An+ были незначимыми (см. табл. 3). На каждые 10 м повышения местности SWC в среднем снижалась на 1.1 ± 0.3% в зарослях A. negundo и на 0.9 ± 0.3% – в зарослях других видов деревьев.
Рис. 3.
Связь объемной влажности почвы (SWC) с высотой местности (а) и суммой осадков, выпавших за 10 сут до измерений (б), в местообитаниях с доминированием Acer negundo (⚫, сплошная линия) и других видов деревьев (⚪, пунктирная линия).
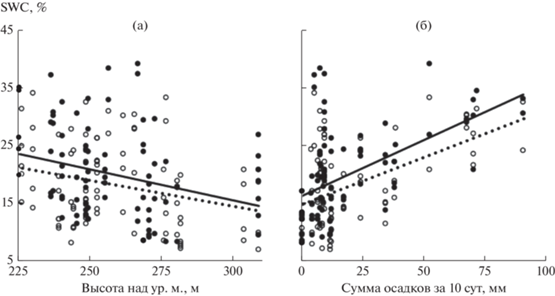
С увеличением суммы осадков, выпавших за 10 сут до измерения, значения SWC закономерно увеличивались независимо от древесного доминанта (рис. 3б). Различия наклонов линий регрессии зависимостей “осадки за 10 сут–SWC” в вариантах An– и An+ были незначимыми (см. табл. 3). На каждые 10 мм выпавших осадков SWC в среднем увеличивалась на 2.0 ± 0.3% в зарослях A. negundo и на 1.6 ± 0.3% – в зарослях других видов деревьев.
Покрытие крон как фактор влажности почвы. Высокая сомкнутость крон и обусловленное этим высокое затенение на поверхности почвы или на уровне травяного яруса – один из механизмов средопреобразующего влияния A. negundo [29, 42]. Поэтому на примере измерений SWC в сезоны 2019 и 2020 гг. мы попытались выяснить, с какой группой факторов более вероятно связана изменчивость SWC – характеристиками сомкнутости крон или наличием/отсутствием опада или подстилки. Для этого построили GLM-модель, включив предикторы: 1) год; 2) схема измерения SWC; 3) сомкнутость крон древесных растений; 4) высота местности; 4) сумма осадков за 10 сут, предшествовавших измерению SWC (табл. 4).
Таблица 4.
Значимость влияния пяти предикторов, включая фактор “сомкнутость крон”, и их взаимодействий в GLM- и LMM-моделях (со случайным фактором “участок”), объясняющих изменчивость объемной влажности почвы в урбанизированных местообитаниях на протяжении вегетационных сезонов 2019 и 2020 гг.
| № | Сочетание предикторов | dF | GLM | LMM | ||
|---|---|---|---|---|---|---|
| F | P | F | P | |||
| 1 | Год | 1 | 86.78 | <0.0001 | 91.50 | <0.0001 |
| 2 | Схема измерения | 1 | 5.28 | 0.0233 | 4.80 | 0.0305 |
| 3 | Сомкнутость крон | 1 | 0.95 | 0.3323 | 0.57 | 0.4506 |
| 4 | Высота местности | 1 | 19.84 | <0.0001 | 9.73 | 0.0048 |
| 5 | Осадки за 10 сут | 1 | 26.92 | <0.0001 | 35.17 | <0.0001 |
| 6 | Год × сомкнутость крон | 1 | 0.03 | 0.8744 | 0.01 | 0.9074 |
| 7 | Год × высота местности | 1 | 0.32 | 0.5698 | 0.30 | 0.5882 |
| 8 | Год × осадки за 10 сут | 1 | 1.36 | 0.2455 | 2.63 | 0.1075 |
| 9 | Схема измерения × сомкнутость крон | 1 | 0.06 | 0.8157 | 0.11 | 0.7398 |
| 10 | Схема измерения × высота местности | 1 | 0.22 | 0.6432 | 0.46 | 0.5016 |
| 11 | Схема измерения × осадки за 10 сут | 1 | 9.95 | 0.0020 | 10.88 | 0.0013 |
| 12 | Сомкнутость крон × высота местности | 1 | 2.86 | 0.0936 | 1.60 | 0.2105 |
| 13 | Сомкнутость крон × осадки за 10 сут | 1 | 0.59 | 0.4422 | 0.49 | 0.4844 |
| 14 | Высота местности × осадки за 10 сут | 1 | 0.50 | 0.4813 | 1.20 | 0.2760 |
| R2 | 0.735 | 0.799 | ||||
| $R_{{{\text{Adj}}}}^{2}$ | 0.703 | 0.774 | ||||
Использованная комбинация предикторов объяснила около 74% общей изменчивости значений SWC в вегетационные сезоны 2019 и 2020 гг. Эффекты, связанные с характеристиками условий на пробных площадях, т.е. с предикторами “год”, “высота местности” и “осадки за 10 сут”, воспроизвелись как статистически значимые. Из двух наиболее интересовавших нас факторов – схема измерений и сомкнутость крон – слабый эффект наблюдался только в отношении первого предиктора. Другими словами, влажность почвы в урбанизированных местообитаниях зависела от режима удаления опада или подстилки, но не зависела от густоты покрова кронами деревьев.
ОБСУЖДЕНИЕ
Объемная влажность почвы в урбанизированных местообитаниях, занятых древесными растениями, сильно различалась в разные годы, а также в зависимости от высоты местности и режима осадков. Менее выраженным было влияние на SWC схемы измерения, т.е. размера эффекта, связанного с давностью удаления подстилки. Различия, связанные с таксономическим положением доминанта древостоя, были небольшими: они ответственны за объяснение лишь небольшой части общей изменчивости оценок SWC. Однако различия, связанные с доминантом древостоя, заметны при разных способах анализа.
Наша рабочая гипотеза подтвердилась: в местообитаниях с доминированием A. negundo наблюдается более высокая объемная влажность почвы, чем в местообитаниях с аналогичным сочетанием условий, но доминированием других видов деревьев. Это заключение надежно, поскольку данные получены на большом числе пробных площадей в серии повторных регистраций влажности на протяжении трех лет. На некоторых площадях повторные измерения SWC выполнены 5 раз. Это заключение также реалистично, поскольку другие закономерности изменчивости SWC, связанные с высотой местности, количеством осадков и межгодовой изменчивостью, ожидаемы и объяснимы. В частности, хорошо понятны: 1) зависимость оценок SWC от количества осадков за предшествующий период – чем больше осадков, тем выше влажность почвы; 2) зависимость SWC от высоты местности – чем выше точка измерения SWC в рельефе, тем ниже влажность почвы; 3) более высокая SWC во влажный и прохладный сезон 2019 г. по сравнению с более теплыми и менее влагообеспеченными сезонами 2020 и 2021 гг. Поэтому можно считать, что и иные закономерности, установленные на нашем массиве оценок SWC, правдоподобны.
Результаты других исследований влажности почвы в зарослях чужеродных видов очень разнообразны: опубликованы свидетельства и снижения [13, 14, 17], и неизменности [13, 14, 20, 21], и увеличения [13, 14, 18, 19] почвенной влаги под чужеродными растениями по сравнению с сообществами аборигенных видов. Одна из причин такой гетерогенности связана с жизненной формой видов [14]. Под чужеродными травами в среднем суше, чем под местными, но под чужеродными деревьями несколько влажнее, чем под местными [14]. Поэтому наши результаты, скорее, согласуются, чем противоречат массиву опубликованных сведений.
Объяснение вероятных причин особенностей влажности почвы в зарослях A. negundo возможно с двух позиций.
Первая состоит в том, что A. negundo чаще занимает более влажные местообитания, чем прочие виды деревьев. Чтобы минимизировать вероятность таких различий, мы использовали блочный дизайн с подбором пар местообитаний, занятых A. negundo и контрольных к ним. Погодные условия и режим осадков в течение вегетационного сезона для местообитаний An– и An+ в масштабе нашего исследования одинаковы. Различаться местообитания An– и An+ могли в первую очередь положением в рельефе. Однако по этому условию ПП в парах в одном блоке были однородны: средняя высота ПП An– составила 261 ± 23 м над ур. м., ПП An+ – также 261 ± 23 м над ур. м. (тест для связанных совокупностей: t = = –0.04; dF = 17; P = 0.9648). Следовательно, наши данные не позволяют считать, что повышенная влажность почвы в зарослях A. negundo объясняется его изначальной избирательностью к условиям увлажнения местообитаний. В целом кажется маловероятным, что могут существовать причины, которые объясняли бы повышенную изначальную влажность почвы в местообитаниях с A. negundo и которые при этом действовали бы в урбанизированных экосистемах, где естественные процессы и закономерности подвержены сильным и непредсказуемым антропогенным трансформациям.
Вторая позиция – это предположение, что разность в значениях SWC между местообитаниями An– и An+ связана с экофизиологическими или экологическими особенностями самого Acer negundo. Другими словами, возможно, что повышенную влажность почвы в местообитаниях с доминированием A. negundo создает он сам. Деревья могут уменьшать влажность почвы путем перехвата осадков кронами [44, 45] и транспирации [14, 26], а повышать путем затенения почвы [25] и создания слоя опада и подстилки, которые замедляют испарение с поверхности почвы [46]. Известно, что древесные растения – и местные, и инвазивные, потребляют больше влаги, чем травянистые [13, 14, 47–49]. Поскольку у мезофильных растений с C3-фотосинтезом транспирация тесно связана с интенсивностью фотосинтеза, то у A. negundo, как у быстрорастущего вида, можно предполагать интенсивные фотосинтез и транспирацию [14, 50, 51]. Однако эти свойства приводили бы к иссушению почвы, а не росту ее влажности.
Повышенную влажность почвы в зарослях A. negundo можно объяснять его способностью создавать высокое затенение или толстую подстилку. Несмотря на наши старания выбирать сходные ПП, сомкнутость крон в варианте An+ была несколько выше (90.0 ± 0.5%), чем в варианте An– (88.5 ± 0.7%), и эти различия были значимы (тест для связанных совокупностей: t = 2.22; dF = 65; P = 0.0303). Под кронами A. negundo также было ожидаемо темнее (13 ± 2 лк × 102), чем под кронами других деревьев (25 ± 4 лк × 102) [29]. Повышенное затенение в зарослях A. negundo наблюдали и другие исследователи [27, 28, 52]. Следовательно, повышенная влажность почвы в зарослях A. negundo может объясняться его способностью формировать густой листовой полог. Но когда мы сопоставили значения SWC с условиями местообитаний (см. табл. 4), это предположение не нашло подтверждения. По нашим оценкам, влажность почвы не зависела от густоты покрова кронами деревьев, но зависела от режима удаления опада или подстилки. На участках, на которых подстилку удаляли до начала периода измерений, было суше, чем на участках, где подстилку удаляли непосредственно перед измерением SWC. Этот феномен понятен. Известно, что подстилка обладает значительной влагоемкостью [53] и препятствует испарению воды из минеральной части почвы [46]. Наблюдаемую разность SWC между местообитаниями An– и An+ можно было бы объяснить, например, если допустить, что в An+ подстилка более развита, чем в An–. Однако характеристики опада и подстилки, скорости их накопления и разложения в зарослях A. negundo не известны. В то же время известна высокая скорость разложения опада A. negundo [54], что не поддерживает предположение о слое подстилки как причине повышенной влажности в местообитаниях с его доминированием.
Таким образом, сопоставление наших результатов с опубликованными данными не позволило легко объяснить причины повышенной влажности почвы в зарослях A. negundo. Есть доводы как за, так и против, что повышенная влажность может быть связана с особенностями строения листового аппарата и крон или с накоплением и разложением опада и подстилки A. negundo, однако информации для надежного решения этого вопроса недостаточно.
ЗАКЛЮЧЕНИЕ
Результаты работы можно резюмировать в виде трех утверждений.
Объемная влажность почвы выше в местообитаниях, где доминирует чужеродное дерево Acer negundo, по сравнению с местообитаниями с другими древесными доминантами. Различие влажности почвы между местообитаниями с доминированием A. negundo и других видов небольшое, но это различие установлено надежно и устойчиво проявляется в разные вегетационные сезоны на фоне других особенностей местообитаний и на фоне влияния погодных условий. Таким образом, основная часть рабочей гипотезы, относящаяся к направлению ожидаемых различий влажности почвы между местообитаниями с доминированием A. negundo и без него, подтвердилась.
Более вероятно, что причиной повышенной влажности почвы в зарослях A. negundo являются не изначальные различия между местообитаниями, а средопреобразующее воздействие самого A. negundo. Это заключение менее надежно, чем первое, но оно подтверждает, что A. negundo – не только чужеродное дерево и инвазивный вид с расширяющимся ареалом и спектром местообитаний, но и вид-трансформер, который может изменять в сообществах-мишенях не только режим освещения [29, 42], но и режим влажности.
Предположение о значимой связи между сомкнутостью крон и влажностью почвы в урбанизированных местообитаниях не подтвердилась. Поэтому для понимания механизмов средопреобразующего воздействия A. negundo необходимы специальные исследования. При этом следует учитывать эффекты, связанные как с надземными органами и процессами, так и накоплением и разложением опада и подстилки, а также другими почвенными процессами.
Работа выполнена в рамках темы госзадания Института экологии растений и животных УрО РАН № 122021000092-9.
Авторы декларируют отсутствие конфликта интересов.
Список литературы
Richardson D.M., Pyšek P., Rejmánek M. et al. Naturalization and invasion of alien plants: Concepts and definitions // Diversity and Distributions. 2000. V. 6. P. 93–107. https://doi.org/10.1046/j.1472-4642.2000.00083.x
Catford J.A., Jansson R., Nilsson C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework // Diversity and Distributions. 2009. V. 15. P. 22–40. https://doi.org/10.1111/j.1472-4642.2008.00521.x
Reinhart K.O., Gurnee J., Tirado R., Callaway R.M. Invasion through quantitative effects: intense shade drives native decline and invasive success // Ecological Applications. 2006. V. 16. P. 1821–1831. https://doi.org/10.1890/1051-0761(2006)016(1821: ITQEIS)2.0.CO;2
Schuster M.J., Reich P.B. Amur maple (Acer ginnala): an emerging invasive plant in North America // Biological Invasions. 2018. V. 20. P. 2997–3007. https://doi.org/10.1007/s10530-018-1754-8
Callaway R.M., Ridenour W.M. Novel weapons: invasive success and the evolution of increased competitive ability // Frontiers in Ecology and the Environment. 2004. V. 2. №. 8. P. 436–443.
Stinson K.A., Campbell S.A., Powell J.R. et al. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms // PLoS Biology. 2006. V. 4. P. e140. https://doi.org/10.1371/journal.pbio.0040140
Suding K.N., Harpole W.S., Fukami T. et al. Consequences of plant-soil feedbacks in invasion // Journal of Ecology. 2013. V. 101. P. 298–308. https://doi.org/10.1111/1365-2745.12057
Mueller K.E., Eisenhauer N., Reich P.B. et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species // Soil Biology and Biochemistry. 2016. V. 92. P. 184–198. https://doi.org/10.1016/j.soilbio.2015.10.010
Veselkin D.V., Kuyantseva N.B., Chashchina O.E. et al. Levels of Leaf Damage by Phyllophages in Invasive Acer negundo and Native Betula pendula and Salix caprea // Russ. J. of Ecol. 2019. V. 50. № 6. P. 511–516. https://doi.org/10.1134/S1067413619060134
Зитте П., Вайлер Э.В., Кадерайт Й.В. и др. Ботаника: в 4 т. // Экология: Пер. с нем., под ред. Еленевского А.Г., Павлова В.Н. М.: Академия, 2007. 248 с.
Clarke N., Gundersen P., Jönsson-Belyazid U. et al. Influence of different tree-harvesting intensities on forest soil carbon stocks in boreal and northern temperate forest ecosystems // For. Ecol. Manag. 2015. № 351. P. 9–19. https://doi.org/10.1016/j.foreco.2015.04.034
Caldwell M., Dawson T.E., Richards J.H. Hydraulic lift: consequences of water efflux from the roots of plants // Oecologia. 1998. V. 113. № 2. P. 151–161. https://doi.org/10.1007/s004420050363
Calder I.R., Dye P. Hydrological impacts of invasive alien plants // Land Use and Water Resources Research. 2001. V. 1. P. 1–12.
Cavaleri M.A., Sack L. Comparative water use of native and invasive plants at multiple scales: a global meta-analysis // Ecology. 2010. V. 91. № 9. P. 2705–2715. https://doi.org/10.1890/09-0582.1
Mason N.W.H., Richardson S.J., Peltzer D.A. et al. Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity // Journal of Ecology. 2012. V. 100. P. 678–689. https://doi.org/10.1111/j.1365-2745.2012.01965.x
Guo X., Xu Z.W., Li M.Y. et al. Increased soil moisture aggravated the competitive effects of the invasive tree Rhus typhina on the native tree Cotinus coggygria // BMC Ecol. 2020. V. 20. P. 1–13. https://doi.org/10.1186/s12898-020-00284-9
Gao F.-Y., Shi F.-X., Chen H.-M. et al. Rapid Expansion of Melica przewalskyi Causes Soil Moisture Deficit and Vegetation Degradation in Subalpine Meadows // Clean – Soil, Air, Water. 2018. V. 46. P. 1700587. https://doi.org/10.1002/clen.201700587
Ruwanza S., Shackleton C.M. Effects of the invasive shrub, Lantana camara, on soil properties in the Eastern Cape, South Africa // Weed Biology and Management. 2016. V. 16. P. 67–79. https://doi.org/10.1111/wbm.12094
Baranova B., Fazekašova D., Manko P. Variations of selected soil properties in the grass fields invaded and uninvaded by invasive goldenrod (Solidago canadensis L.) // Ekologia (Bratislava). 2017. V. 36. № 2. P. 101–111.
Stefanowicz A.M., Stanek M., Nobis M., Zubek S. Few effects of invasive plants Reynoutria japonica, Rudbeckia laciniata and Solidago gigantea on soil physical and chemical properties // Sci. Total Environ. 2017. № 574. P. 938–946. https://doi.org/10.1016/j.scitotenv.2016.09.120
Ruwanza S., Tshililo K. Short term soil and vegetation recovery after Acacia mearnsii removal in Vhembe Biosphere Reserve, South Africa // Applied Ecology and Environmental Research. 2019. V. 17 № 2. P. 1705–1716. https://doi.org/10.15666/aeer/1702_17051716
Nilsson C., Engelmark O., Cory J. et al. Differences in litter cover and understory flora between stands of introduced lodgepole pine and native Scots pine in Sweden // For. Ecol. Manag. 2008. № 255. P. 1900–1905. https://doi.org/10.1016/j.foreco.2007.12.012
Cusack D.F., McCleery T.L. Patterns in understory woody diversity and soil nitrogen across native- and non-native-urban tropical forests // For. Ecol. Manag. 2014. № 318. P. 34–43. https://doi.org/10.1016/j.foreco.2013.12.036
Berg C., Drescherl A., Essl F. Using relevé-based metrics to explain invasion patterns of alien trees in temperate forests // Tuexenia. 2017. № 37. P. 127–142. https://doi.org/10.14471/2017.37.012
Sagar R., Pandey A., Singh J.S. Composition, species diversity, and biomass of the herbaceous community in dry tropical forest of northern India in relation to soil moisture and light intensity // The Environmentalist. 2012. V. 32. P. 485–493.
Leishman M.R., Haslehurst T., Ares A., Baruch Z. Leaf trait relationships of native and invasive plants: community- and global-scale comparisons // New Phytologist. 2007. V. 176. P. 635–643. https://doi.org/10.1111/j.1469-8137.2007.02189.x
Saccone P., Pagès J.P., Girel J. et al. Acer negundo invasion along a successional gradient: early direct facilitation by native pioneers and late indirect facilitation by conspecifics // New Phytol. 2010. V. 187. P. 831–842. https://doi.org/10.1111/j.1469-8137.2010.03289.x
Bottollier-Curtet M., Charcosset J., Poly F. et al. Light interception principally drives the understory response to boxelder invasion in riparian forests // Biol. Invasions. 2012. V. 14. P. 1445–1458. https://doi.org/10.1007/s10530-011-0170-0
Veselkin D.V., Dubrovin D.I., Rafikova O.S. et al. Shading and light interception in thickets of invasive Acer negundo and Sorbaria sorbifolia // Russ. J. of Biological Invasions. 2022. № 4. P. 30–42.
Fire Effects Information System (FEIS). USDA ForestService, Rocky Mountain Research Station, Fire Sciences laboratory [Online]. URL: https:// www.feis-crs.org/feis/faces/index.xhtml;jsessionid= 643C2E17A6A3897DE7A5164F7B70016B (дата обращения – 25.04.2021).
Emelyanov A.V., Frolova S.V. Ash-leaf maple (Acer negundo L.) in coastal phytocenoses of the Vorona River // Rus. J. Biol. Invasions. 2011. № 2. P. 161–163.
Казанцева Т.И., Бобровская Н.И., Тищенко В.В. Особенности восстановления залежной растительности луговых степей Центрального Черноземья (Воронежская область) // Аридные экосистемы. 2010. Т. 16. №. 42. С. 76–86.
Куликов П.В., Золотарева Н.В., Подгаевская Е.Н. Эндемичные растения Урала во флоре Свердловской области // Екатеринбург: “Гощицкий”, 2013. 610 с.
Pyšek P., Lambdon P.W., Arianoutsou M. et al. Alien Vascular Plants of Europe // DAISIE handbook of alien species in Europe. Dordrecht: Springer Netherlands, 2009. P. 43–61.
Дубовик Д.В., Дмитриева С.А., Ламан В.Н. и др. Черная книга флоры Беларуси: чужеродные вредоносные растения. Минск: Беларуская навука, 2020. 407 с.
Виноградова Ю.К., Майоров С.Р., Хорун Л.В. Черная книга флоры Средней России: чужеродные виды растений в экосистемах Средней России. М.: ГЕОС. 2010, Т. 2. 512 с.
Dgebuadze Yu.Yu., Petrosyan V.G., Khlyap L.A. The most dangerous invasive species of Russia (TOP-100). Moscow: KMK Scientific Press, 2018. 688 p.
Третьякова А.С., Куликов П.В. Адвентивный компонент флоры Свердловской области: биоэкологические особенности // Вестник Удмуртского ун-та. Серия “Биология. Науки о Земле”. 2014. № 1. С. 57–67.
Veselkin D.V., Korzhinevskaya A.A., Podgayevskaya E.N. Tree species composition and abundance of alien and invasive understory shrubs and trees in urban forests of Yekaterinburg // Vestnik Tomskogo gosudarstvennogo universiteta. Biologiya. 2018. V. 42. P. 102–118.
Gusev A.P., Shpilevskaya N.S., Veselkin D.V. The influence of Acer negundo L. on progressive successions in Belarusian landscapes // Vestnik Vitebskogo Gosudarstvennogo Universiteta. 2017. V. 94. P. 47–53.
Veselkin D.V., Dubrovin D.I. Diversity of the grass layer of urbanized communities dominated by invasive Acer negundo // Russ. J. of Ecol. 2019. V. 50. P. 413–421. https://doi.org/10.1134/S1067413619050114
Veselkin D.V., Dubrovin D.I., Pustovalova L.A. High canopy cover of invasive Acer negundo L. affects ground vegetation taxonomic richness // Sci. Rep. 2021. V. 11. P. 20T758. https://doi.org/10.1038/s41598-021-00258-x
Погода и климат [Электронный ресурс]. URL: http://www.pogodaiklimat.ru (дата обращения – 01.02.2022).
Le Maitre D.C., Scott D.F., Colvin C. Review of information on interactions between vegetation and groundwater // Water SA. 1999. V. 25 № 2. P. 137–152.
Kagawa A., Sack L., Duarte K., James S. Hawaiian native forest conserves water relative to timber plantation: species and stand traits influence water use // Ecological Applications. 2009. V. 19. P. 1429–1443. https://doi.org/10.1890/08-1704.1
Magliano P.N., Giménez R., Houspanossian J. et al. Litter is more effective than forest canopy reducing soil evaporation in Dry Chaco rangelands // Ecohydrology. 2017. V. 10. 10 p. https://doi.org/10.1002/eco.1879
Enright W.D. The effect of terrestrial invasive alien plants on water scarcity in South Africa // Physics and Chemistry of the Earth. Part B: Hydrology, Oceans and Atmosphere. 2000. V. 25. P. 237–242.
Farley K.A., Jobbagy E.G., Jackson R.B. Effects of afforestation on water yield: a global synthesis with implications for policy // Glob. Change. Biol. 2005. V. 11. P. 1565–1576.
Huxman T.E., Wilcox B.P., Breshears D.D. et al. Ecohydrological implications of woody plant encroachment // Ecology. 2005. V. 86. P. 308–319.
Vertessy R.A., Watson F.G.R., O’Sullivan S.K. Factors determining relations between stand age and catchment water balance in mountain ash forests // For. Ecol. Manag. 2001. № 143. P. 13–26. https://doi.org/10.1016/S0378-1127(00)00501-6
Irvine J., Law B.E., Kurpius M.R. et al. Age-related changes in ecosystem structure and function and effects on water and carbon exchange in ponderosa pine // Tree Physiology. 2004. V. 24. № 7. P. 753–763. https://doi.org/10.1093/treephys/24.7.753
Porte A.J., Lamarque L.J., Lortie C.J. et al. Invasive Acer negundo outperforms native species in non-limiting resource environments due to its higher phenotypic plasticity // BMC Ecology. 2011. V. 11. P. 28. https://doi.org/10.1186/1472-6785-11-28
Zhou Q., Keith D.M., Zhou X. et al. Comparing the Water-holding Characteristics of Broadleaved, Coniferous, and Mixed Forest Litter Layers in a Karst Region // Mountain Research and Development. 2018. V. 38. № 3. P. 220–229. https://doi.org/10.1659/MRD-JOURNAL-D-17-00002.1
Janušauskaitė D., Straigyte L. Leaf litter decomposition differences between alien and native maple species // Baltic Forestry. 2011. V. 17. P. 189–196.
Дополнительные материалы отсутствуют.