

Экология, 2022, № 5, стр. 345-354
Структура и запас фитомассы как показатели стадии демутации степных залежей Тувы
Н. И. Макунина a, *, А. Д. Самбуу b
a Центральный Сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
b Тувинский институт комплексного освоения природных ресурсов СО РАН
667007 Кызыл, ул. Интернациональная, 117/а, Россия
* E-mail: natali.makunina@mail.ru
Поступила в редакцию 16.03.2022
После доработки 30.03.2022
Принята к публикации 08.04.2022
- EDN: UFBMIS
- DOI: 10.31857/S0367059722050067
Аннотация
Статистически проверены следующие эмпирические выводы: каждая стадия сукцессии имеет определенную продолжительность, в ее процессе запас надземной фитомассы уменьшается, подземной – увеличивается, однако на терминальных стадиях запас подземной фитомассы существенно меньше, чем в целинных степях. Основой для работы послужили 54 образца фитомассы залежных сообществ и целинных степей; они были отобраны в период с 1994 г. по 2020 г. в четырех степных межгорных котловинах Тувы. Впервые получены количественные характеристики запасов живой надземной, мертвой надземной и подземной фитомассы на каждой стадии восстановительной сукцессии тувинских залежей. Стадии залежной демутации в Туве значимо отличаются по запасу либо надземной, либо подземной фитомассы. Значимые отличия возраста залежей получены только между начальными (стадии 1 и 2) и терминальными (стадии 3 и 4) стадиями демутации. Подтверждено, что подземная фитомасса целинных степей выше, чем сообществ терминальных стадий залежной сукцесии.
В современном мире антропогенно нарушенной растительности одной из ключевых задач является изучение процесса ее восстановления (демутации). Такие исследования, с одной стороны, региональны, с другой – имеют глобальное значение, поскольку именно они могут стать основой для разработки корректных методов эксплуатации земель в разных частях земного шара.
Один из вариантов вторичной сукцессии – процесс восстановления степной растительности на залежах. К настоящему времени о ней известно следующее: залежная демутация “однонаправлена, детерминирована, проходит определенные стадии и обусловливает появление растительного сообщества, существовавшего до распашки целины”, однако ход каждой конкретной сукцессии индивидуален и зависит от множества случайных обстоятельств; в разных регионах залежная степная сукцессия однотипна [1]. Принято выделять четыре стадии восстановления распаханных степей [2]: 1 – стадия полевых сорняков (бурьянистая), 2 – стадия корневищных растений (корневищная), 3 – стадия кустовых злаков (разнотравно-злаковая), 4 – стадия вторичной целины. Эти стадии характерны для всей степной растительности, однако флористический состав залежных сообществ и продолжительность стадий в разных степных регионах различны.
Процесс восстановления степных залежей привлекает внимание многих исследователей. В основном в работах анализируются изменения флористического состава в зависимости от возраста залежей. Подобные исследования охватывают большинство степных регионов России: Воронежская обл. [3], Саратовская обл. [4], Республика Башкирия [5], Оренбургская обл. [6], Алтайский край [7, 8], Республика Хакасия [9], Республика Тыва [10], Республика Бурятия [11].
Другое направление связано с оценкой запаса фитомассы нарушенных травяных сообществ (например, [12–18]). В России такие исследования проводятся на Южном Урале [19, 20], в Западной Сибири [21], в Хакасии [22] и Туве [1, 23], однако создать на их основе общую картину не представляется возможным. Этот факт обусловлен несколькими причинами. Обычно авторы характеризуют только продуктивность сообществ, данные о подземной фитомассе публикуют гораздо реже. Из-за трудоемкости сбора материала в большинстве работ представлена информация об единичных образцах фитомассы, что делает невозможным проведение статистического анализа. Но основная причина заключается в том, что параметры фитомассы отдельных стадий демутации залежей до сих пор не выявлены. В результате статистически достоверные данные о запасах фитомассы на залежах, находящихся на разных стадиях демутации, отсутствуют. Тем не менее для территории Тувы накоплен достаточный объем информации, позволяющий охарактеризовать запас фитомассы и ее фракций на каждой из стадий восстановления степных залежей.
Исследования продуктивности и запаса фитомассы степных залежей и целинных степей позволили сделать ряд статистически непроверенных эмпирических заключений. С нашей точки зрения, интерес представляют следующие положения [23]: запас надземной фитомассы в ходе восстановительной сукцессии на залежах уменьшается, а подземной – увеличивается; каждая из стадий демутации имеет определенную продолжительность; запас подземной фитомассы на финальных стадиях демутации меньше, чем в целинных степях.
Цель настоящей работы – статистически проверить сформулированные ниже гипотезы:
Гипотеза 1. Масса фракций фитомассы значимо отличается на каждой из четырех стадий залежной демутации.
Гипотеза 2. Структура фитомассы коррелирует с возрастом залежи, а возраст сообществ каждой из стадий значимо отличается.
Гипотеза 3. Запас подземной фитомассы сообществ, находящихся на заключительных стадиях демутации, значимо меньше, чем запас подземной фитомассы целинных степей.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в четырех степных межгорных котловинах Тувы (рис. 1). Территория Тувы расположена в двух биоклиматически отличающихся районах: семигумидном и семиаридном [24]. Две котловины (Турано-Уюкская, 800–1000 м над ур. м. и Улуг-Хемская, 500–800 м над ур. м.) относятся к семигумидному району, их фоновую растительность представляют крупнодерновинные и мелкодерновинные степи. Две другие котловины (Хемчикская, 800–1000 м над ур. м. и Убсу-Нурская, 800–1000 м над ур. м.) расположены в семиаридном биоклиматическом районе, в их растительном покрове преобладают мелкодерновинные степи.
Рис. 1.
Карта-схема Тувы. Межгорные котловины: 1 – Турано-Уюкская, 2 – Улуг-Хемская, 3 – Хемчикская, 4 – Убсу-Нурская.
Природные условия в степных котловинах мало подходят для земледелия: среднегодовая температура варьирует от –3 до –5°C, средняя температура января – от –30 до –33°C, июля – от 18 до 20°C. В семиаридных котловинах выпадает 150–250 мм осадков в год, в семигумидных – 250–350 мм; для земледелия неблагоприятны и сухая весна (дожди начинаются только в июне), и короткое лето (заморозки возможны во второй половине августа).
Мы изучали залежные сообщества разного возраста и целинные степи. Чтобы охарактеризовать распространение залежей в Туве, обратимся к истории ее земледелия. До середины XX в. распахивали небольшие участки в долинах рек Турано-Уюкской и Улуг-Хемской котловин, к 1944 г. – год вхождения Тувинской Народной Республики в состав СССР – пашни занимали 58.1 тыс. га. В процессе реализации всесоюзного проекта освоения целины площадь пашен увеличилась до 361.8 тыс. га: в Турано-Уюкской, Улуг-Хемской и Хемчикской котловинах были распаханы практически все дренированные участки долин рек и водоразделы с уклоном до 3°, пашни появились даже в Убсу-Нурской котловине. В начале 1990-х годов огромные площади пашен были заброшены: к 2008 г. в Туве распахивали только 20.8 тыс. га. Таким образом, для изучения залежной сукцессии в Туве стали доступны более 300 тыс. га залежей – шестая часть территории этой горной республики. Началом залежной сукцессии принято считать 1994 г., когда поля повсеместно перестали распахивать.
В Туве условия для изучения процессов восстановления степной растительности на залежах можно считать идеальными: известные даты повсеместного начала процесса демутации позволяют вести точную хронологию, а обширная территория залежей дает возможность получения достоверных результатов. В первое десятилетие с начала залежной сукцессии проводились лишь единичные исследования, но именно благодаря им первые стадии демутации не были упущены. В последующие два десятилетия работы стали более регулярными и охватили все основные залежные территории Тувы.
Основой для работы послужили 54 образца фитомассы, отобранные А.Д. Самбуу в период с 1994 г. по 2020 г.: 41 образец характеризует залежи разного возраста, 13 – целинные степи. Эти образцы взяты в разных частях четырех тувинских межгорных котловин: 22 образца представляют залежи и целинные степи семиаридных, 32 образца – семигумидных котловин. В разные годы на 7 ключевых участках, представляющих типичные залежи котловин, был отобран 31 образец фитомассы; в “неохваченных районах” в разные годы дополнительно были собраны еще 10 образцов; 13 образцов фитомассы целинных степей взяты в непосредственной близости от ключевых участков.
Каждый образец характеризовали 3 фракциями фитомассы: G – надземная живая, D + L – надземная мертвая (ветошь и подстилка), B + V – подземная (живая и мертвая). Сбор материала производили следующим образом. На каждом участке закладывали 8–10 площадок (10 × 10 м), на которых случайным образом выделяли 8 квадратов 50 × 50 см, на уровне почвы срезали фитомассу и собирали подстилку. В середине каждой площадки отбирали почвенные монолиты поверхностью 10 см2 и глубиной 10 см (в этом слое расположено от 50 до 90% корней), затем корни отмывали от почвы. Всю фитомассу высушивали и взвешивали [23].
Все вычисления проведены в программе PAST [25]. Для кластерного анализа применен метод Уорда (Ward’s method). При проверке гипотез сначала с помощью непараметрического статистического критерия Краскелла-Уоллеса (H) проверена значимость наблюдаемых различий. Для дальнейшего попарного сравнения использован непараметрический статистический критерий Манна-Уитни (U). Корреляция между признаками рассчитана с помощью рангового коэффициента Спирмена (rS). В тексте фракции фитомассы характеризуются медианными значениями.
Графики построены в программе PAST. Использовано стандартное изображение боксплотов: нижняя и верхняя “стенки ящика” соответствуют значениям 1-го и 3-го квартилей, “полоска в ящике” – медиане, “усы” – минимальному и максимальному значениям. Одинаковыми буквенными индексами показаны гомогенные по критерию Манна-Уитни значения.
Названия видов растений даны по сводке С.К. Черепанова [26].
РЕЗУЛЬТАТЫ
Целинные степи. Степи – фоновая растительность всех тувинских котловин. Эдификаторами крупнодерновинных степей являются злаки (Stipa capillata, Helictotrichon altaicum) и Carex pediformis; виды степного (Medicago falcata, Schizonepeta multifida, Scabiosa ochroleuca) и лугово-степного (Fragaria viridis и Phlomoides tuberosa) разнотравья не обильны. Травостой покрывает почву на 60–80%, средняя высота травостоя, соответствующая высоте прикорневых листьев злаков, 25–30 см. Травостой мелкодерновинных степей образуют злаки (Agropyron cristatum, Cleistogenes squarrosa, Koeleria cristata, Poa botryoides, Stipa krylovii) и степное разнотравье (Artemisia frigida, Potentilla acaulis). Его высота и проективное покрытие сильно зависят от метеорологических условий года, в среднем составляя соответственно 10 см и 40%.
Флористический состав залежей разного возраста. Через 5 лет сукцессии на месте пашен сформировались бурьянистые сообщества, состоящие из одно-, двулетних полыней (Artemisia scoparia, A. sieversianа) и маревых (Chenopodium album). В последующее пятилетие доля бурьяна уменьшилась, возросло участие длиннокорневищного злака Elytrigia repens, появились единичные экземпляры сегетальных (Convolvulus arvensis) и степных (Heteropappus altaicus) видов разнотравья. Через 10 лет после начала сукцессии в залежных сообществах преобладали корневищные злаки (Elytrigia repens, Poa angustifolia), появились единичные экземпляры рыхлокустовых и дерновинных видов (Leymus dasystachys, Stipa capillata). Спустя 20 лет в равной степени были представлены корневищные и дерновинные злаки; содоминирующую группу разнотравья составляли немногочисленные сорные (Artemisia scoparia) и степные (Medicago falcata, Heteropappus altaicus, Galium verum) виды. И сейчас, спустя почти 30 лет после начала сукцессии, залежные сообщества отличаются от целинных степей. Первое, что бросается в глаза, – это неравномерность травостоя: крупные пятна образуют злаки (Elytrigia repens, Poa angustifolia, Stipa capillata) и Artemisia scoparia; на их фоне выделяются более мелкие пятна степного разнотравья (Medicago falcata, Heteropappus altaicus). При более внимательном изучении обращают на себя внимание “нестепной характер” ряда злаков-доминантов (Elytrigia repens, Poa angustifolia), постоянное присутствие сорных видов и неполночленность флористического состава сообществ.
Критерии для выделения стадий восстановления залежей. Если образцы фитомассы залежных сообществ на каждой из 4 стадий демутации действительно различаются, то критерием их разделения должна быть одна или несколько характеристик фитомассы. Взяв за основу эмпирическое утверждение о том, что “доля корней в процессе демутации залежных сообществ увеличивается” [23], в качестве критерия мы выбрали структуру фитомассы. Для первичного упорядочения используемого материала построили ряд по увеличению доли их подземной фитомассы. Чтобы получить общее представление об особенностях структуры фитомассы залежных сообществ по сравнению с целинными степями, были использованы данные всех 54 образцов (залежных и целинно-степных). После проведения кластерного анализа мы выделили пять базисных кластеров, которые предположительно могли соответствовать четырем стадиям демутации и целинным степям (рис. 2) и содержали разное число образцов: первая группа – 7, вторая – 9, третья – 16, четвертая – 8, пятая – 14. Предварительно мы полагали, что образцы фитомассы целинных степей окажутся преимущественно в пятой группе, однако этого не произошло: целинно-степные образцы присутствовали в каждой из трех последних групп.
Рис. 2.
Дендрограмма структуры фитомассы залежных сообществ и целинных степей: 1–5 – базовые кластеры.

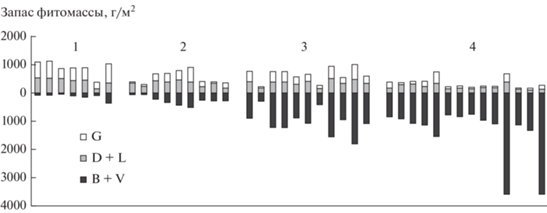
Для дальнейшего анализа мы исключили образцы целинных степей, оставив в наборе 41 образец залежей разного возраста; нумерацию образцов сохранили. После проведения кластерного анализа выделили 4 базисные группы: первая – 7, вторая – 9, третья – 11 и четвертая – 14 образцов. Затем мы сравнили расположение образцов в группах на рис. 2 и 3. Образцы фитомассы, представляющие первые три группы, не изменили своего положения, в то время как образцы фитомассы залежей из 4-й и 5-й групп на рис. 2 образовали одну группу на рис. 3. Графическая характеристика структуры фитомассы залежных сообществ представлена на рис. 4, запасов фитомассы – на рис. 5; порядок расположения образцов на рис. 4 и 5 совпадает.
Рис. 3.
Дендрограмма структуры фитомассы залежей: 1–4 – базовые кластеры залежей. Нумерация образцов на рис. 2 и 3 совпадает.
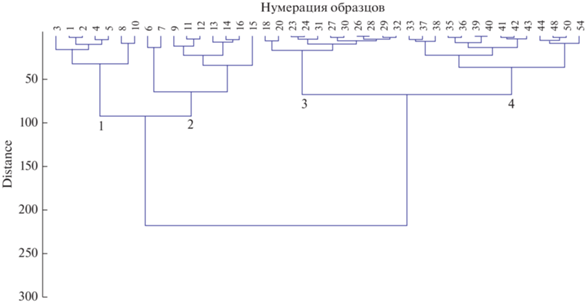
Рис. 4.
Стуктура фитомассы залежных сообществ: G – доля надземной живой фитомассы, D + L – доля надземной мортмассы, B + V – доля подземной фитомассы; 1–4 – стадии демутации.

Рис. 5.
Запас фитомассы залежных сообществ: G – надземная живая фитомасса, D + L – надземная мортмасса, B + V – подземная фитомасса; 1–4 – стадии демутации.
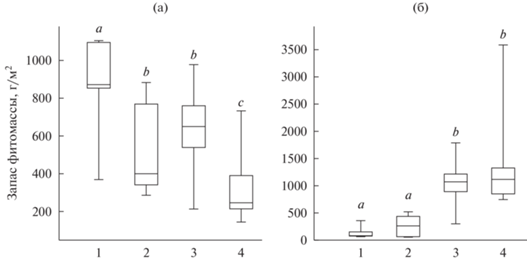
Четыре группы образцов, выделенные на рис. 3–5, значимо отличаются по доле подземной (Н = 46.98, P = 3.35E-10) и надземной (H = 47.45, P = 2.65E-10) фитомассы. Согласно нашим представлениям, они соответствуют четырем стадиям демутации залежей и характеризуются следующими параметрами (рис. 6):
Рис. 6.
Доля надземной (а) и подземной (б) фитомассы; 1–4 – стадии демутации. Здесь и на рис. 7–9 одинаковыми буквенными индексами показаны гомогенные по критерию Манна-Уитни значения.

1-я стадия: надземная фитомасса абсолютно преобладает (ее 1-й квартиль – 82%, медиана – 89%, 3-й квартиль – 93%);
2-я стадия: надземная фитомасса немного больше, чем подземная (1-й квартиль – 60%, медиана – 64%, 3-й квартиль – 79%);
3-я стадия: подземная фитомасса несколько преобладает (1-й квартиль – 62%, медиана – 62%, 3-й квартиль – 64%);
4-я стадия: подземная фитомасса заметно преобладает (1-й квартиль – 74%, медиана – 79%, 3-й квартиль – 85%).
Запас фитомассы в залежных сообществах. Надземная фитомасса максимальна на 1-й стадии (872 г/м2), на 2-й стадии она составляет 400 г/м2, на 3-й – 650 г/м2, минимальным значением характеризуется 4-я стадия (239 г/м2).
Запасы фракций надземной фитомассы изменяются следующим образом. Живая надземная фитомасса залежных сообществ уменьшается с 452 г/м2 на 1-й стадии до 195 г/м2 на 2-й, на 3-й стадии она составляет 261 г/м2 и 77 г/м2 – на 4-й стадии. Надземная мортмасса на 1-й стадии составляет 444 г/м2, на 2-й – 325 г/м2, на 3-й – 365 г/м2, на 4-й – 168 г/м2.
Подземная фитомасса в процессе залежной сукцессии увеличивается: на 1-й стадии она составляет 82 г/м2, на 2-й – 262 г/м2, на 3 – 1072 г/м2, на 4-й – 1103 г/м2
Результаты проверки гипотезы 1. Запасы надземной фитомассы начальной (1) и терминальной (4) стадий значимо отличаются от запасов фитомассы промежуточных стадий (2 и 3) (рис. 7а). Запасы подземной фитомассы на 1-й и 2-й стадиях демутации значимо отличаются от этих показателей на 3-й и 4-й стадиях (рис. 7б).
По запасам надземной живой фитомассы значимо отличается только начальная 1-я стадия (рис. 8а). Значимых отличий надземной мортмассы между стадиями не выявлено (рис. 8б).

Результаты проверки гипотезы 2. Между стадией демутации и реальным возрастом залежи выявлена заметная корреляция (rS = 0.59). Медианное значение возраста залежей 1-й стадии демутации составляет 4 года, 2-й – 7 лет, медианный возраст сообществ 3-й и 4-й стадий не отличается и равен 27 годам; стадии 1 и 2 по возрасту значимо отличаются от стадий 3 и 4.
Результаты проверки гипотезы 3. Мы сравнили запасы подземной фитомассы залежных сообществ терминальных стадий 3 и 4 (25 образцов) и целинных степей (13 образцов). Запасы подземной фитомассы залежей составили 1073 г/м2, целинных степей – 2360 г/м2; отличие значимо (U = 34, P = = 8.19E-5).
Медианные значения надземной живой фитомассы на залежах, находящихся на терминальных стадиях демутации, и в целинных степях совпадают и составляют 195 г/м2 (U = 142.5, P = 0.55). Медианные значения надземной мортмассы равны 264 г/м2 в залежных сообществах и 268 г/м2 – в целинных степях; показатели значимо не отличаются (U = 135, P = 0.41).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Статья посвящена одной из самых длительно разрабатываемых тем экологии – вторичной сукцессии. Мы представляем свой взгляд на одно из ее проявлений – восстановительную сукцессию степных залежей. На эту тему опубликовано немало работ, в которых стадии демутации обычно разграничивают по возрасту и/или по преобладающим жизненным формам. Нередко дополнительно используют характеристику флористического состава сообществ и/или их параметры (видовая насыщенность, проективное покрытие), количественные характеристики запасов фитомассы упоминают в единичных случаях; статистический анализ результатов часто не проводится.
Мы реализовали принципиально отличающийся подход и использовали как критерий разделения на стадии залежной демутации структуру фитомассы. Для этого был необходим материал, собранный в одном регионе на залежах разного возраста по единой методике в минимально достаточном количестве. Выбранный нами критерий разделения оправдал себя: 4 основных кластера сообществ, интерпретированные как 4 стадии демутации, значимо отличаются друг от друга по доле надземной и подземной фитомассы (рис. 9). В терминах нашей концепции описание стадий будет следующим: 1-я стадия – надземная фитомасса абсолютно преобладает; 2-я стадия – надземная фитомасса немного больше подземной; 3-я стадия – подземная фитомасса несколько преобладает; 4-я стадия – подземная фитомасса заметно преобладает.
Рис. 9.
Запас надземной живой (G), надземной мертвой (D + L) и подземной (B + V) фитомассы на терминальных стадиях залежной демутации (1) и целинных степях (2).
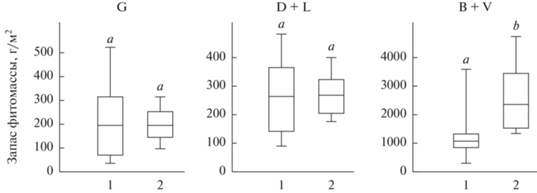
Полученные результаты позволили впервые охарактеризовать запасы фитомассы залежных сообществ и ее фракций на каждой из четырех стадий демутации в Туве и подтвердили правомерность использования непараметрических методов: распределение количественных значений фракций фитомассы на каждой из четырех стадий заметно отличается от нормального. Этот факт подчеркивает необходимость применения для количественных характеристик запасов фитомассы медианных, а не средних значений.
При характеристике фитомассы травяных сообществ часто используется лишь один ее параметр – продуктивность (запас надземной живой фитомассы). Наш анализ позволил определить эффективность его применения: оказалось, что только сообщества начальной 1-й стадии демутации имеют значимо более высокую продуктивность; сообщества более продвинутых стадий по этому параметру значимо не отличаются.
Полученные количественные характеристики фитомассы на каждой из стадий залежной демутации позволили проверить ряд эмпирических заключений, высказанных А.А. Титляновой и А.Д. Самбуу [23].
Первое заключение гласит, что запас надземной фитомассы на каждой последующей стадии демутации уменьшается, а подземной увеличивается. Полного статистического подтверждения эта гипотеза не получила. Надземная фитомасса значимо отличается на 1-й и 2-й, а также на 3-й и 4-й стадиях, тогда как 2-я и 3-я стадии не имеют значимых различий. Для подземной фитомассы ситуация противоположна: подземная фитомасса значимо отличается на 2-й и 3-й стадиях, а 1-я и 2-я также, как 3-я и 4-я стадии, по этому параметру значимо не отличаются. Таким образом, для тувинских степных залежей мы можем переформулировать гипотезу следующим образом: стадии залежной демутации значимо отличаются или по запасу надземной, или по запасу подземной фитомассы.
Мы проверили эту гипотезу на примере хакасских степей, исходные данные для которых взяты из работы [22]. Результаты несколько отличаются от тувинских: тренда уменьшения надземной фитомассы в процессе залежной демутации не выявлено, однако тенденция увеличения подземной фитомассы отмечается. Эта ситуация может быть объяснена недостаточным объемом материала и отличием критериев выделения стадий демутации (в цитируемой работе стадии выделены по преобладающим жизненным формам и возрасту залежи).
Вторая гипотеза гласила, что структура фитомассы коррелирует с возрастом залежи. Действительно, между этими параметрами наблюдается заметная корреляция, однако только 2-я и 3-я стадии значимо отличаются по возрасту; две начальные, как и две терминальные, стадии демутации значимых различий не имеют. С учетом принятых нами критериев этот факт позволяет утверждать, что залежные сообщества, характеризующиеся преобладанием надземной фитомассы, по возрасту значимо отличаются от залежных сообществ с преобладанием подземной фитомассы.
Третья гипотеза заключалась в том, что запас подземной фитомассы сообществ, находящихся на заключительных стадиях демутации, меньше, чем запас подземной фитомассы целинных степей; наблюдение статистически подтверждено. Значимых отличий в запасе живой фитомассы и ее фракций не обнаружено.
Таким образом, большая часть рассмотренных нами эмпирических выводов полностью или частично подтверждается статистически.
ЗАКЛЮЧЕНИЕ
Наша работа открывает новый этап изучения залежных сообществ Тувы. От построения конкретных рядов демутации и характеристики фитомассы на отдельных ключевых участках мы перешли к обобщению результатов для всей территории Тувы. Показано, что надежным показателем стадии восстановления степных залежей является структура фитомассы. Впервые количественно охарактеризован запас фитомассы залежных сообществ на каждой стадии демутации степей. Статистически проверен ряд эмпирических предположений. Оказалось, что стадии залежной демутации тувинских степей значимо отличаются либо по запасу надземной, либо по запасу подземной фитомассы. Значимые отличия возраста залежей получены только между 1–2-й начальными и 3–4-й терминальными стадиями демутации. Подтверждено утверждение о том, что подземная фитомасса целинных степей выше, чем сообществ терминальных стадий залежной сукцессии.
Работа выполнена в рамках государственного задания Центрального Сибирского ботанического сада СО РАН АААА-А21-121011290026-9 при финансовой поддержке РФФИ (проект № 19-29-05208/19 мк).
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Титлянова А.А., Самбуу А.Д. Детерминированность и синхронность залежной сукцессии в степях Тувы // Изв. РАН. Серия биологич. 2014. Т. 6. С. 621– 621.
Семенова-Тянь-Шанская А.М. Восстановление растительности на степных залежах в связи с вопросом о “происхождении” видов // Бот. журн. 1953. № 6. С. 862–873.
Панкратова Л.А., Ганнибал Б.К. Восстановительные сукцессии травяных сообществ в ландшафтах южной лесостепи (Воронежская область, музей-заповедник “Дивногорье”) // Вестник Санкт-Петербургского ун-та. Науки о Земле. 2009. Т. 2. С. 92–95.
Шевченко Е.Н., Сергеева И.В., Пономарева А.Л., Зябирова М.М. Способы определения возраста залежных земель на примере южной части Приволжской возвышенности Саратовской области // Аграрный научн. журн. 2016. Т. 4. С. 49–53.
Хусаинова С.А., Халикова А.А., Хусаинов А.Ф. Анализ восстановительных сукцессий на залежных землях Башкирского Предуралья // Ботаника в современном мире: Труды XIV съезда Русского ботанич. общ-ва и конференции. 2018. Т. 2. С. 145–147.
Ледовский Н.В., Абаимов В.Ф., Ходячих И.Н. Стадийность развития растительности залежей в ходе сукцессионного процесса // Изв. Оренбургского гос. аграрного ун-та. 2016. Т. 3. № 59. С. 176–179.
Овчарова Н.В., Ямалов С.М. Синтаксономический и ординационный анализы восстановительных сукцессий травяной растительности правобережья реки Оби (Алтайский край) // Изв. Самарского научного центра РАН. 2013. Т. 15. № 3–1. С. 388–394.
Овчарова Н.В. Флора и растительность залежей правобережья р. Оби (Алтайский край). Барнаул: Изд-во Алт. ун-та, 2015. 250 с.
Егунова Н.А. Экологическое состояние залежных экосистем в период вторичных сукцессий в сухостепной зоне Республики Хакасия // Вестник Тувинского гос. ун-та. Естественные и сельскохозяйственные науки. 2011. Т. 2. С. 89–93.
Дубровский Н.Г., Намзалов Б.Б., Ооржак А.В., Куулар М.М. Флористико-геоботанические и биоэкологические исследования залежной растительности Тувы // Вестник Бурятского гос. ун-та. Биология. География. 2018. Т. 1. С. 27–43.
Холбоева С.А. Флора и растительность залежей долины р. Чикой (Западное Забайкалье) // Проблемы изучения растительного покрова Сибири: Мат-лы VI междун. научн. конф., посвященной 100-летию со дня рождения А. В. Положий. Томск, 2017. С. 106–108.
Jain S.K. Total phytomass, net community productivity and system transfer functions in sub-humid grasslands at Sagar (MP) India // Flora. 1980. V. 170. № 3. P. 251–260.
Kutiel P., Danin A. Annual-species diversity and aboveground phytomass in relation to some soil properties in the sand dunes of the northern Sharon Plains, Israel // Vegetatio. 1987. V. 70. № 1. P. 45–49.
Ford M.A., Wilson F. Seasonal change in the above-ground phytomass of an Agrostis-with-Festuca grassland in North-east Scotland // Botanical J. of Scotland. 1993. V. 46. № 3. P. 427–432.
Joshi M. Seasonal changes in species diversity, phytomass and net primary production in a temperate grassland in Kumaun, Central Himalaya, India // Journal of Tropical Forest Science. 1995. P. 521–531.
Okamoto C., Nakamura M., Kabata K., Wang Y. Seasonal changes in the aboveground phytomass of a Stipa baicalensis community in Northeast China // Japanese J. of Grassland Science. 2000. V. 46. № 3–4. P. 217–222.
Chu Y., He W.M., Liu H.D. et al. Phytomass and plant functional diversity in early restoration of the degraded, semi-arid grasslands in northern China // Journal of Arid Environments. 2006. V. 67. № 4. P. 678–687.
Vazquez-de-Aldana B.R., Garcia-Ciudad A., Garia-Criado B. Interannual variations of above-ground biomass and nutritional quality of Mediterranean grasslands in Western Spain over a 20-year period // Crop & Pasture Science. 2008. V. 59. P. 769–779.
Ходячих И.Н. Водный режим и урожайность сухой массы на разновозрастных залежах // Изв. Оренбургского гос. аграрного ун-та. 2012. Т. 1. № 33. С. 50–52.
Абаимов В.Ф., Ледовский Н.В., Ходячих И.Н. Типы залежей степной зоны Южного Урала и их хозяйственно-биологическая оценка // Изв. Оренбургского гос. аграрного ун-та. 2013. Т. 4. № 42. С. 227–230.
Овчарова Н.В., Терехина Т.А. Продуктивность надземной фитомассы травяных сообществ Алтайского края в ходе восстановительной сукцессии // Проблемы ботаники Южной Сибири и Монголии. 2014. Т. 13. С. 148–152.
Чупрова В.В., Ковалева Ю.П. Особенности продукционно-деструкционных процессов в разновозрастных залежах Койбальской степи Минусинской котловины // Доклады по экологическому почвоведению. 2018. Т. 7. С. 1–23.
Титлянова А.А., Самбуу А.Д. Сукцессии в травяных экосистемах. Новосибирск: Изд-во СО РАН, 2016. 191 с.
Makunina N.I. Botanical and geographical characteristics of forest steppe of the Altai-Sayan mountain region // Contemporary Problems of Ecology. 2016. V. 9. № 3. P. 342–348. https://doi.org/10.1134/S1995425516030100
Hammer Ø. PAST Paleontological Statistics. Version 2.17. Reference manual. Oslo: University of Oslo, 2012. 227 c.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб., 1995. 992 с.
Дополнительные материалы отсутствуют.