

Экология, 2023, № 1, стр. 22-34
Неоднородность почвенно-растительного покрова при постагрогенной восстановительной сукцессии в средней подзоне тайги
Е. Н. Наквасина a, Т. А. Паринова a, *, А. Г. Волков a, Л. В. Голубева b
a Северный (Арктический) федеральный университет имени М.В. Ломоносова
163002 Архангельск, наб. Северной Двины, 17, Россия
b Архангельский педагогический колледж
163002 Архангельск, ул. Смольный Буян, 5, Россия
* E-mail: t.parinova@narfu.ru
Поступила в редакцию 05.03.2022
После доработки 08.09.2022
Принята к публикации 12.09.2022
- EDN: HAWHYD
- DOI: 10.31857/S0367059723010080
Аннотация
Представлены результаты исследований по выявлению особенностей неоднородности почвенно-растительного покрова на ранних стадиях самовосстановительной сукцессии, детерменированных биотическими и биокосными условиями, измененными под влиянием антропогенных факторов после длительной распашки на азональных окарбоначенных почвах в подзоне средней тайги (Россия, Архангельская обл., Каргопольский р-н). Объем анализируемого материала: 25 полнопрофильных почвенных разрезов, 500 почвенных проб, 125 площадок для учета видов деревьев и кустарников, 25 стандартных геоботанических пробных площадей, 250 площадок для учёта травянистых видов. Показано, что в первые 15 лет восстановительной сукцессии на залежном поле приоритет в фитоценотической значимости имеет травянистая растительность. В условиях остаточно карбонатных почв лесной зоны специализация экологических свойств, как приуроченность видов к определенным факторам, проявляется слабо и только у отдельных травянистых видов. Факторами формирования мозаичности растительного покрова на 15-летней залежи являются не изменчивость почвенных свойств и влияние поселившихся древесных и кустарниковых видов, а биологические свойства видов травянистых растений, их специфические потребности к экологическим факторам. Плодородие и увлажнение почв – главные факторы, обусловливающие варьирование растительности на этой стадии.
Изучению неоднородности растительного и почвенного покровов в последнее время уделяется все больше внимания. Считается, что неоднородность является основополагающей в сохранении биоразнообразия и существовании видов и поддержания экосистем [1–5], а ее снижение может привести к потере биоразнообразия [6].
Формирование неоднородности представляется закономерным природным процессом [7, 8], главными факторами которого являются биотические [5], создающие фитоценогенные микрогруппировки [9]. При этом признается роль биокосных факторов, связанных с педогенными причинами и почвообразующими породами [9–12]. Относительно земель, используемых в сельском хозяйстве, однозначно признается влияние агротехнических и агромелиоративных приемов в сочетании с естественно-генетическими факторами на формирование неоднородности почвенного покрова как в отношении морфогенетических показателей, так и изменчивости агрохимических свойств почв [4, 10, 12–15]. Уровень сельскохозяйственной нагрузки и тип землепользования определяют комбинации факторов в природно-антропогенных экосистемах, которые способствуют специализации среды обитания видов [16].
Распахивание почв природных ландшафтов при вовлечении в сельскохозяйственную деятельность влияет на формирование неоднородности почв неоднозначно. Гетерогенность почвенного покрова по отдельным параметрам может снижаться, по другим увеличиваться – эти изменения имеют региональный характер [15, 17]. Так, агрогенное воздействие выравнивает агрофизические и агрохимические показатели верхнего почвенного слоя [9, 12, 18], но в то же время при интенсивном сельскохозяйственном производстве неравномерность внесения удобрений приводит к усилению гетерогенности связанных с ними свойств [8, 19]. Кроме того, на пашнях, в верхнем гомогенном слое почв (пахотном горизонте), могут сохраняться мозаики природных почв, связанные с внутренним рельефом почвообразующих пород [9, 11] и сохраняющие свое влияние во время восстановительной сукцессии. Распахивание почв нарушает естественные процессы и сохраняет свое воздействие длительное время [12, 15], оказывая влияние на сукцессионные процессы. Мозаичность почвенно-растительного покрова при демутационных сукцессиях усиливается при расселении на открытые местообитания опушечной и луговой растительности [18].
Неоднородность растительного и почвенного покрова различных ландшафтов чаще рассматривалась представителями отдельных специальностей – геоботаниками, почвоведами, агрономами, а число комплексных работ невелико. Знание пространственной неоднородности почв востребовано для точного земледелия: для оптимального управления продуктивностью посевов с учетом плодородия почв на каждом метре поля [2, 7, 8, 15], для мониторинга и оценки уровня деградации земель [20] и сельскохозяйственного управления [21]. Мозаичность растительности в различных ландшафтах изучали для познания структуры фитоценозов, динамики смен растительности, моделирования биоразнообразия и изучения продуктивности биогеоценозов [5, 13, 22–24]. Все чаще экосистемы рассматриваются комплексно, с междисциплинарных точек зрения [21], основываясь на мозаично-цикличной концепции развития экосистем [25] и учитывая их разные элементы [4, 25, 26].
При этом важным считается изучить не только мозаичность растительного покрова и почвенной среды, но и их взаимодействие [26, 27] в системе “вид–среда обитания” [2, 28]. Мозаичность почвенного покрова обеспечивает неоднородность и разнообразие автохтонных и аллохтонных видов растений и микроорганизмов [4, 29], так как создаются экологические ниши для их поселения. Так, на черноземах именно пространственная составляющая и экологические функции почв обеспечили 43.6% разнообразия почвенной микрофлоры [30].
Особый интерес с точки зрения неоднородности почвенно-растительного покрова вызывают залежи (выведенные из оборота пашни), проходящие этапы восстановительной сукцессии при зарастании лесом. Отчуждение пашни не только влияет на рост древесной растительности, но и запускает различные механизмы процесса самозарастания [31, 32]. Поселение видов деревьев и кустарников начинает усиливать прежде всего почвенную неоднородность [20, 28]. В то же время, по данным Vellend et al. [33], в лесах, сформированных на бывших полях, биоразнообразие ниже по сравнению с нативными, что может сохраняться столетиями.
Отчуждение сельскохозяйственных земель из активного пользования, охватившее не только Россию, но практически все регионы мира, вызывает проблему экологических изменений в ландшафтах, которые предсказать невозможно [34] из-за значительных локальных и региональных особенностей влияния абиотических и биотических факторов, типов окружающих залежи сообществ, предшествующих агротехнических воздействий и применяемых технологий. Для прогнозирования реакции видов на изменение ландшафта, динамики формирования сообществ необходимо изучение процессов зарастания таких земель с учетом типологических характеристик почв, общей сукцессионной смены растительности и истории поля.
Цель наших исследований – выявить особенности неоднородности почвенно-растительного покрова на ранних стадиях самовосстановительной сукцессии, детерминированные биотическими и биокосными условиями, измененными влиянием антропогенных факторов после длительной распашки, для прогнозирования потенциала формирования лесных насаждений в подзоне средней тайги на азональных окарбоначенных почвах.
МАТЕРИАЛ И МЕТОДЫ
Для Европейского Севера России исторически характерно мелкополье. В советское время нередко проводили перепланировку полей, в частности при укрупнении совхозов, что приводило к объединению полей разных сроков использования, припахиванию вновь осваиваемых участков с разной технологией освоения. Все это влияло на мозаичность почвенного покрова в пределах большого поля при его забрасывании. Окружающий лес является источником обсеменения залежи лесными видами. Под пологом древесной растительности на землях сельскохозяйственного пользования сохраняется потенциал сорной и культурной растительности многопольного севооборота.
Объектом нашего изучения было заброшенное поле площадью 44.7 га в районе бывшей деревни Васильево (Каргопольский р-н Архангельской обл., 61°30′41.79″ с.ш., 38°47′30.92″ в.д.). Предмет изучения – неоднородность почвенно-растительного покрова заброшенного поля.
Неоднородность понимается нами как варьирование свойств в пространстве и во времени [35], а однородность соответственно как константность значений в пределах изучаемого объекта [36]. Неоднородность почвенного покрова залежного поля анализируется нами через оценку показателей, характеризующих пространственную изменчивость (мощность пахотного слоя, гранулометрический состав, содержание элементов питания), которая возникает в процессе залежеобразования. Неоднородность растительного покрова оценивается через элементы горизонтальной структуры растительности, которая проявляется в том, что на разных участках преобладают (имеют высокий индекс фитоценотической значимости) растения разных видов и/или отсутствуют растения каких-либо видов, встречающиеся в окружающих залежь фитоценозах.
Для Каргопольской суши, где проводились исследования, характерны азональные текстурно-метаморфические грубогумусированные остаточно-карбонатные почвы [37], сформированные на карбонатной морене и известковой плите. Эти почвы широко использовались в сельском хозяйстве, как правило, обладают одновременно практически несовместимыми свойствами: высокой гумусностью, высоким значением рН, с одной стороны, и оподзоленностью, с другой.
Рассматриваемое нами поле имеет сложную историю, связанную с двукратным укрупнением: в послевоенное время поле укрупняли за счет отжига и распашки лесного участка, позднее (1960-е годы) – за счет припахивания старых огородов бывшей деревни и поймы ручья. Таким образом, время сельскохозяйственного использования поля колеблется от 30 лет до более века (на полях деревенских огородов). В последние десятилетия перед забрасыванием залежное поле использовалось в многопольном севообороте. По данным СХП “Каргопольское”, в севообороте выращивали кормовые многолетние злаки, бобовые культуры (горох, клевер), зерновые культуры (овес, ячмень, озимую и яровую рожь), после уборки культур поле передавалось под осеннее пастбище для крупного рогатого скота. Поле заброшено окончательно в 1999 г., обследовано нами в 2015 г.
На территории поля вдоль его длинной стороны, примыкающей к лесу, заложили трансекту: 25 точек с интервалом 20 м, общая длина трансекты 500 м (рис. 1). Подобная методика изучения мозаичности почвенно-растительного покрова применяется как в России, так и за рубежом [13, 14, 38].
Рис. 1.
Расположение трансекты на поле в районе бывшей деревни Васильево Каргопольского р-на Архангельской области (а) и местоположение объекта исследований на Русской равнине (б).
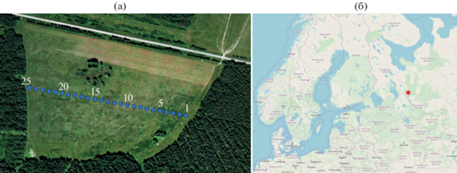
Общий рельеф поля равнинный, высота над уровнем моря – 136 м, встречаются линейные перепады 50–80 см, представленные колеями, оставленными машинами. Поле с трех сторон окружено лесом. В нулевой точке трансекты произрастает 90-летний сосняк хвощово-разнотравный (состав 8С2Е + Б): средняя высота Pinus sylvestris – 29.5 ± 0.8 м, диаметр – 33.3 ± 1.7 см. Согласно перечету на пяти площадках размером 2 × 5 м, подрост (2300 шт/га) представлен Pinus sylvestris, Picea abies × P. obovata, Populus tremula и Betula pendula × B. pubescens (состав 39С35Е17Ос9Б) в основном средней крупности (0.5–1 м); подлесок (900 шт/га) представлен Rosa acicularis, Sorbus aucuparia и Juniperus communis. Почва органо-аккумулятивная светлогумусовая глееватая остаточно-карбонатная среднесуглинистая.
В конечной точке трансекты произрастает 80-летний осинник разнотравный (снытьевый), в состав которого входят Populus tremula, Betula pendula × B. pubescens, Alnus incana и Picea abies × × P. obovata, Pinus sylvestris (8Ос2Б + Олх ед. Е, С). Диаметр Populus tremula составляет в среднем 52.2 ± ± 5.6 см, высота – 32.2 ± 1.3 м. Подрост (11 000 шт/га) представлен Alnus incana и Picea abies × P. obovata (состав 91Олх9Е) высотой не более 1.5 м, подлесок (2400 шт/га) – Padus racemosa, Salix caprea и Sorbus aucuparia. Почва текстурно-дифференцированная дерново-подзолистая остаточно-карбонатная легкосуглинистая.
Точки опробования закрепляли в натуре колышками с указанием номера. У каждой из точек закладывали почвенный разрез с описанием горизонтов и диагностикой почвы. Отбирали пробы почвы металлическим буром для определения плотности сложения по общепринятой методике с толщи пахотного горизонта (5–15 см) с интервалом 1 м на расстоянии 10 м в обе стороны от точки опробования (20 проб в каждой точке опробования, всего 500 проб). Эту же почву использовали для химических анализов, проводимых по общепринятым методикам. Состав и численность видов деревьев и кустарников описывали на 5 площадках размером 2 × 5 м, расположенных вблизи опорной точки, напочвенный покров – на 10 площадках размером 1 × 1 м, расположенных в пределах геоботанической площади 10 × 10 м, с центром в точке опробования. Всего было заложено: 25 полнопрофильных почвенных разрезов, 125 площадок для учета видов деревьев и кустарников, 25 стандартных геоботанических пробных площадей, 250 площадок для учета травянистых видов. Для каждого вида определяли проективное покрытие по равномерной шкале (0–5–10–15…95–100%). Номенклатура видов указана по сводке С.К. Черепанова [39].
Чтобы учесть проективное покрытие и встречаемость вида в напочвенном покрове, использовали фитоценотический индекс (индекс Понятовской – Сырокомской, индекс фитоценотической значимости, ИФЗ) [40]:
где I – фитоценотический индекс, ИФЗ; n – число площадок, на которых отмечен вид; N – общее число площадок; ΣP – сумма проективных покрытий вида на отмеченных площадках. Для удобства значения полученных индексов логарифмировали.Анализировали данные о соотношении на трансекте эколого-ценотических групп видов растений [41], групп по отношению к фактору увлажнения, фактору трофности почвы, соотношение групп по жизненным формам [42].
Элементный состав пахотного горизонта почв определяли на CHNS анализаторе EA 3000 в Центре коллективного пользования научным оборудованием “Арктика” Северного (Арктического) федерального университета имени М.В. Ломоносова. Подвижные формы фосфора и калия определяли после экстрагирования раствором соляной кислоты (0.2 моль/дм), с последующим количественным измерением подвижных соединений фосфора на фотоэлектроколориметре и калия – на пламенном фотометре в лаборатории САС “Архангельская”.
Для анализа флористической структуры и экологических условий в пределах трансекты использована ординация методом NMS (мера расстояния – коэффициент Съеренсена) в программном пакете PC-ORD 6.0 [43]. Интерпретация полученных осей ординации проведена с использованием индикаторных значений по экологическим шкалам Д.Н. Цыганова [44], расчет которых выполнен в программе Ecoscale 5.0 [45]. Для анализа видового списка, расчета индексов сходства/различия и ординации использовали R и пакет Vegan [46].
РЕЗУЛЬТАТЫ
Неоднородность почвенного покрова. Почва поля диагностируется как старопахотная реградированная текстурно-дифференцированная дерново-подзолистая остаточно-карбонатная. Однако на линии трансекты выделяются две почвенные разности, занимающие 60 и 40% площади поля соответственно. Отличия проявляются прежде всего в строении нижней части почвенного профиля, не затронутой вспашкой. Это не позволяет отнести почвенный покров поля к однородному в типологическом отношении.
Вскипание почвы от HCl на глубине 40–55 см подтверждает окарбоначенность почвообразующей породы. Изменчивость толщи пахотного горизонта и общей толщи почвы менее 25% (табл. 1), и показатели в пределах трансекты оцениваются как однородные [47]. Изменчивость толщи образующегося реградированного горизонта W (39%) носит ценотический (вторичный) характер, при сохранении первичных признаков почвообразования (толщи почвы до почвообразующей породы) и выравненного пахотной деструкцией агрогенного горизонта Pw.
Таблица 1.
Характеристики почвенных переменных
| Показатель | M ± m | Минимум– максимум | Стандартное отклонение | Коэффициент вариации, % | |
|---|---|---|---|---|---|
| Содержание, мг/кг | P2O5 | 151.8 ± 14.0 | 56.0–290.0 | 69.9 | 46 |
| K2O | 59.9 ± 6.3 | 28.0–136.0 | 31.6 | 53 | |
| Элементный состав, % | N | 0.09 ± 0.01 | 0.01–0.16 | 0.05 | 41 |
| C | 1.25 ± 0.05 | 0.58–1.76 | 0.26 | 21 | |
| Содержание гумуса, % | 2.2 ± 0.1 | 1.0–3.0 | 0.5 | 21 | |
| C/N | 15.8 ± 1.8 | 7.0–35.0 | 8.0 | 50 | |
| pHKCl | 5.8 ± 0.1 | 4.8–6.8 | 0.5 | 8 | |
| Плотность сложения, г/см3 | 1.25 ± 0.01 | 1.19–1.30 | 0.03 | 3 | |
| Мощность, см | W | 7.7 ± 0.6 | 3.0–12.0 | 3.0 | 39 |
| Pw | 19.8 ± 0.7 | 13.0–26.0 | 3.3 | 17 | |
| Профиль | 43.7 ± 1.5 | 31.0–62.0 | 7.5 | 17 | |
Деструктивные изменения почвы, созданные 30-летней вспашкой, сохраняются, несмотря на 15-летнее восстановление и зарастание видами трав, деревьев и кустарников. Плотность сложения почвы имеет минимальную изменчивость из всех изученных показателей (3%), что согласуется с исследованиями других авторов [12, 15].
Слабая изменчивость по протяженности трансекты в пределах поля характерна для обменной кислотности, содержания органического углерода и связанного с ним содержания гумуса (8–21%). Несмотря на низкую изменчивость показателя рНKCl, реакция среды значимо влияет на содержание подвижного калия (r = 0.73 при p ≤ 0.05) и определяет степень сформированности реградированного горизонта (r = − 0.59 при p ≤ 0.05).
В нашем случае неоднородность пахотного горизонта в пределах поля проявляется по содержанию подвижных форм фосфора и калия (рис. 2а), коэффициент изменчивости которых составляет 46–53%. Это может быть связано как с изначальной невыравненностью внесения удобрений в период сельскохозяйственного использования поля, так и носить ценотический характер и заключаться в поступлении опада различного качества при зарастании поля. На большую изменчивость содержания фосфора в пахотных горизонтах дерново-подзолистых почв указывается в ряде источников [8, 19], однако уровень изменчивости определяется литологией почв [15].
Рис. 2.
Изменчивость содержания подвижных форм фосфора и калия (а) и показателя C/N (б) в пределах трансекты залежного поля.

В пределах изученного поля изменчивость содержания азота (41%) при небольшой изменчивости количества органического углерода (21%) обеспечивает высокую изменчивость показателя C/N, который колеблется от 7 до 35 при коэффициенте изменчивости 50% (табл. 1, рис. 2б). По мнению ряда авторов [3, 26, 29], пространственная неоднородность содержания N и показателя C/N в почве может способствовать повышению разнообразия растительности, в том числе и древесной, играя активную роль в сохранении богатства видов. Есть мнение [48], что именно содержание органического углерода определяет 50% физических свойств почвы, прежде всего структурообразование, и, следовательно, совместно с азотом отражает изменения, происходящие как в органогенной, так и минеральной частях почвы под влиянием лесных пород [31].
Установлено, что в пределах трансекты залежного поля влияние содержания азота и углерода на изученные показатели и свойства почвы незначительное (табл. 2). Прослеживается лишь значимая связь содержания подвижного фосфора с углеродом (r = − 0.5 при p ≤ 0.05). Прямая значимая корреляционная связь (r = −0.5, достоверен при p ≤ 0.05) между показателем С/N и мощностью формирующегося реградированного горизонта Pw в верхней части пахотного горизонта позволяет говорить о связи величины C/N с развитием дернины при залужении поля. Этот горизонт густо насыщен корнями и обеспечивает запасы корневого опада, насыщенного азотом, при разрастании бобовых, в частности Trifolium pratense. Мощность реградированного горизонта оказалась не связана с содержанием азота и калия (r = 0.4 при p ≤ 0.05), но в определенной мере обусловлена рыхлостью пахотного горизонта (r = 0.4 при p ≤ 0.05).
Таблица 2.
Корреляционная связь почвенных показателей с факторами гумусонакопления
| Показатели | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| N, % | 0.0 | –0.2 | 1.0 | 0.4 | 0.4 | 0.1 | –0.7 | 0.4 | 0.3 | 0.3 | 0.0 |
| C, % | –0.5 | 0.1 | 0.4 | 1.0 | 1.0 | 0.2 | –0.3 | –0.1 | 0.0 | –0.2 | 0.3 |
| C/N | 0.0 | 0.4 | –0.7 | –0.3 | –0.3 | –0.01 | 1.0 | –0.5 | –0.3 | –0.3 | –0.1 |
Примечание: 1 – содержание P2O5, мг/кг; 2 – содержание K2O, мг/кг; 3 – содержание N, %; 4 – содержание C, %; 5 – содержание гумуса, %; 6 – рНKCl; 7 – показатель C/N; 8 – мощность реградированного горизонта W, см; 9 – мощность пахотного горизонта Pw, см; 10 – мощность профиля, см; 11 – плотность сложения, г/см3. Полужирным выделены достоверные значения коэффициента корреляции.
Неоднородность растительного покрова. В результате процесса восстановительной сукцессии на заброшенном поле сформировался растительный покров, характеризующийся определенными сходными чертами, с одной стороны, и неоднородностью, с другой. Общий флористический список сосудистых растений в пределах трансекты (всего флористический учет вдоль трансекты выполнен на 3750 м2: 125 площадок по 10 м2 для учета видов деревьев и кустарников; 25 стандартных геоботанических пробных площадей по 100 м2, в пределах которых 250 площадок по 1 м2 для учета травянистых видов) включает 101 вид из 75 родов и 33 семейств. Основу составляют покрытосеменные растения (отдел Magnoliophyta), среди них преобладают двудольные; отдел Equisetophyta представлен тремя видами из рода Equisetum; отдел Polypodiophyta – одним видом из рода Athyrium; отдел Lycopodiophyta – одним видов из рода Lycopodium. Кроме того, на трансекте зарегистрированы три вида из отдела Bryophyta, один напочвенный вид из Lichenophyta.
В целом наибольшее участие в сложении растительного покрова залежного сообщества (по количеству видов и их проективному покрытию) принимают травянистые короткокорневищные гемикриптофиты со средней требовательностью к почвенному питанию и влаге (рис. 3). Деревья и кустарники (исходя из численности и габитусных размеров) пока играют незначительную роль в сложении залежного сообщества (см. рис. 3), однако сами по себе, при отсутствии всех видов хозяйственной деятельности, они являются существенным перспективным ценотическим фактором, повышающим уровень неоднородности [49]. Преобладающей эколого-ценотической группой ожидаемо (при отсутствии пока большей ценотической роли деревьев и кустарников) является луговая-луговоопушечная – это виды Dactylis glomerata, Trifolium pratense, Taraxacum officinale и др.
Рис. 3.
Соотношение эколого-ценотических групп видов в пределах трансекты залежного поля: Br – бореальная; H – высокотравная; Md – луговая и лугово-опушечная; Nm – неморальная; Nt – нитрофильная; Pn – боровая; Wt – водно-болотная (гигрофильная).

В то же время считается, что заметное воздействие древесных пород на почву и нижние ярусы растительности проявляется уже в первые 20 лет [50] и заключается в постепенном изменении уровня освещенности, качества и количества опада, состава кроновых вод и прежде всего отражается на почвенной биоте [51]. В условиях Каргопольской суши деревьям и кустарникам для этого требуется больше времени.
Пул древесных пород на изучаемом поле представлен основными лесообразующими видами: Pinus sylvestris, Picea abies, Betula pubescens, B. pendula, Populus tremula, Alnus incana, а также видами подлеска (Sorbus aucuparia, Padus avium, Juniperus communis, Rosa majalis, Salix aurita, S. caprea, S. acutifolia). Средняя численность для видов деревьев – 63 ± 6.1 шт/100 м2, основных лесообразующих видов подроста – 47 ± 4.2 шт/100 м2, видов подлеска – 16 ± 3.6 шт/100 м2, причем подлесок на 86% представлен видами ив.
Окружение поля лесными массивами с трех сторон определило одновременное поселение на нем основных лесообразующих видов посредством распространения семян от стен леса. По возрастному и размерному составу пула подроста (от всходов до деревьев высотой более 1.5 м) просматривается первоочередное заселение видами хвойных: Pinus sylvestris – доля в численности составляет 67%, в меньшей степени Picea abies – 19%. Доля подроста лиственных невелика (14% от общей численности деревьев), среди них много всходов Betula pendula (до 64 шт/100 м2).
Вдоль трансекты в разных точках количество отдельных древесно-кустарниковых видов и их численность неоднородны (рис. 4). Особенно высокая изменчивость численности характерна для подлеска – от 1 до 51 шт/100 м2; для подроста коэффициент изменчивости численности в пределах трансекты составляет 45%.
Рис. 4.
Изменчивость численности видов деревьев (а) и кустарников (1), и в том числе хвойных (2), и количества видов травянистого яруса (б) в пределах трансекты залежного поля.

Видовая насыщенность на 100 м2 для травяного яруса вдоль трансекты в среднем составляет 27 видов – по отдельным участкам трансекты она варьирует от 14 до 35 видов. Проявляется влияние численности древесных и кустарниковых пород на видовое разнообразие травяного яруса (r = − 0.25 при p ≤ 0.05). Разрастание крон деревьев и кустарников изменяет уровень освещенности, меняется растительный опад, однако в молодом возрасте влияние древесных пород не оказывает кардинального влияния на состав напочвенного покрова, что согласуется с мнением ряда исследователей [50]. По их данным, признаки леса при зарастании полей начинают проявляться через 20 лет после отчуждения и начала зарастания древесными и кустарниковыми породами. На этой стадии сукцессионного развития, несмотря на поселение видов деревьев и кустарников, залежный ценоз все еще относится к луговой стадии развития, с преобладанием травянистых рыхлокустовых видов.
Наибольший индекс фитоценотической значимости обнаружен для таких видов травяного яруса, как Trifolium pratense (ИФЗ = 7.5), Hypericum perforatum (ИФЗ = 6.5), Galium mollugo (ИФЗ = 6.4), Briza media (ИФЗ = 6.4) и Leucanthemum vulgare (ИФЗ = 5.8). Показатель учитывает встречаемость вида и покрытие, но в пределах трансекты проективное покрытие видов с высоким фитоценотическим индексом меняется. Интересно отметить, что по нашим наблюдениям Elytrigia repens всегда является индикатором молодых залежей на пойменных почвах района исследований, а в условиях 15-летнего самовосстановления окультуренных окарбоначенных почв не проявляет высокую фитоценотическую значимость. Этому могло способствовать быстрое разрастание других видов, в частности из семейства бобовых, в условиях повышенного плодородия почв с благоприятной для них реакцией среды.
Характер распространения видов, поселившихся на залежном поле после его отчуждения из активного севооборота, обусловлен многими факторами: возможностью заноса зачатков, семенным и/или вегетативным расселением уже внедрившихся видов, воздействием внешних факторов (например, остановками гусеобразных на пролете) и в том числе свойствами верхнего (пахотного) горизонта почвенного покрова, вариабельность которых будет формировать фитоценотические группировки. Почвенные условия на отдельных участках вдоль трансекты создают экологические ниши для определенных видов и влияют на их локальную приуроченность, но при этом сказывается влияние комплекса самой формирующейся растительности, в частности зарождающиеся конкурентные и ценотические отношения между видами древесного и травянистого ярусов.
ОБСУЖДЕНИЕ
Связь неоднородности растительного и почвенного покровов. Агрогенное воздействие на неоднородный по генезису почвенный покров и формирование пахотного горизонта приводит к относительной выравненности свойств верхнего горизонта почвы, которая может сохраняться длительное время [12]. Однако в условиях 15-летнего самовосстановления биогеоценоза значимого влияния неоднородности свойств почв на формирование растительного покрова не выявлено. В данном случае большее влияние оказывают изменения, обусловленные заселением растительности из близлежащих растительных сообществ, а также сохранением семенных и вегетативных зачатков после действующих севооборотов. Одним из таких видов является фитоценотический лидер среди травянистых растений Trifolium pratense.
Влияние неоднородности свойств почвы проявляется специализированно, прежде всего в разрастании отдельных видов травянистого покрова. Так, для видов с высоким ИФЗ показана значимая корреляционная связь проективного покрытия с почвенными показателями в пределах трансекты (табл. 3). Большую значимость в их произрастании имеют содержание углерода и реакция среды. Соответствие экологической ниши конкретному виду растения обусловлено специфическими потребностями растений.
Таблица 3.
Корреляционная связь показателей растительного покрова с почвенными показателями в пределах трансекты залежного поля
| Показатели растительного покрова | Почвенные показатели | ||||
|---|---|---|---|---|---|
| Р2О5 | K2О | N | C | рНKCl | |
| Численность, шт/100 м2: | |||||
| древесных и кустарниковых видов | –0.19 | 0.34 | 0.25 | –0.04 | 0.22 |
| сосны | –0.25 | 0.26 | 0.37 | 0.25 | 0.27 |
| ели | –0.05 | –0.05 | –0.11 | –0.15 | –0.11 |
| ивы | –0.15 | 0.14 | 0.20 | 0.13 | 0.12 |
| Число травянистых видов, шт. | –0.51 | –0.22 | 0.05 | 0.32 | 0.16 |
| Общее количество видов, шт. | –0.52 | –0.27 | –0.15 | 0.20 | 0.11 |
| Виды растительного покрова (ИФЗ, lg): | |||||
| Leucanthemum vulgare (5.8) | – 0.26 | 0.24 | 0.28 | 0.38 | 0.26 |
| Galium mollugo (6.4) | – 0.05 | –0.43 | –0.04 | 0.08 | –0.38 |
| Briza media (6.4) | – 0.18 | 0.29 | 0.41 | 0.65 | 0.45 |
| Hypericum perforatum (6.5) | 0.03 | –0.42 | –0.08 | –0.08 | –0.52 |
| Trifolium pratense (7.5) | 0.13 | –0.14 | –0.01 | –0.43 | –0.20 |
Корреляционный анализ показал, что существует тесная прямая связь между проективным покрытием Leucanthemum vulgare и содержанием в почве углерода, проективным покрытием Briza media с кислотностью почвы и содержанием в ней азота и углерода, а тесная обратная связь – между проективным покрытием Galium mollugo и Hypericum perforatum с содержанием калия, покрытием Trifolium prаtense и содержанием углерода.
Результаты упорядочивания видов общего флористического списка для изученного участка залежного поля неметрическим многомерным шкалированием, представленные на рис. 5, демонстрируют неоднородность флористической структуры и наличие определенных градиентов. Отсутствие четко просматриваемых кластеров характерно как для древесных и кустарниковых видов, так и для травянистых. Векторы почвенных свойств, полученные с использованием экологических шкал Д.Н. Цыганова [44], наложенные на график ординации (см. рис. 5), показывают наиболее существенное влияние на неоднородность древесно-кустарниковой и травянистой растительности в условиях Каргопольской суши таких факторов, как богатство почв азотом и увлажнение почв. При этом Pinus sylvestris, Picea abies приурочены к менее богатым почвам, а Populus tremula тяготеет к участкам с повышенным содержанием азота в почве. Такое же распределение, с приуроченностью к более высокому содержанию азота в почве, характерно для видов подлеска (Sorbus aucuparia, Padus avium, Juniperus communis, Rosa majalis, Salix aurita, S. caprea, S. acutifolia).
Рис. 5.
Результаты упорядочивания видов растительного покрова залежного поля неметрическим многомерным шкалированием (красным выделены древесные и кустарниковые виды): Nt – богатство почв азотом, Hd – увлажнение почв, Tm – термоклиматическая шкала. Травянистые виды: AchiMill – Achillea millefolium L., AegoPoda – Aegopodium podagraria L., AgroTenu – Agrostis tenuis Sibth., AlchVulg – Alchemilla sp., AmorRepe – Amoria repens (L.) C. Presl, AngeSylv – Angelica sylvestris L., AnthOdor – Anthoxanthum odoratum L., AnthSylv – Anthriscus sylvestris (L.) Hoffm., AsarEuro – Asarum europaeum L., BrizMedi – Briza media L., BromIner – Bromopsis inermis (Leyss.) Holub, BuniOrie – Bunias orientalis L., CampGlom – Campanula glomerata L., CampPatu – Campanula patula L., CampPers – Campanula persicifolia L., CentScab – Centaurea scabiosa L., ChamAngu – Chamaenerion angustifolium (L.) Scop., CirsArve – Cirsium arvense (L.) Scop., CirsHete – Cirsium heterophyllum (L.) Hill, DactGlom – Dactylis glomerata L., DactInca – Dactylorhiza incarnata (L.) Soo, DescCesp – Deschampsia cespitosa (L.) P. Beauv., EquiArve – Equisetum arvense L., EquiPrat – Equisetum pratense Ehrh., EuphBrev – Euphrasia brevipila Burnat & Gremli, FestPrat – Festuca pratensis Huds., FragVesc – Fragaria vesca L., GaleSpec – Galeopsis speciosa Mill., GaliMoll – Galium mollugo L., GeraSylv – Geranium sylvaticum L., GeumRiva – Geum rivale L., HeraSibi – Heracleum sibiricum L., HeraSosn – Heracleum sosnówskyi Manden., HierPilo – Hieracium pilosella L., HierUmbe – Hieracium umbellatum L., HypePerf –Hypericum perforatum L., JuncBufo – Juncus bufonius L., KnauArve – Knautia arvensis (L.) J.M. Coult., LathPrat – Lathyrus pratensis L., LeucVulg – Leucanthemum vulgare Lam., LuzuPilo – Luzula pilosa (L.) Willd., MyosPalu – Myosotis palustris (L.) L., OrchMili – Orchis militaris L., PhlePrat – Phleum pratense L., PimpSaxi – Pimpinella saxifraga L., PlanLanc – Plantago lanceolata L., PlanMajo – Plantago major L., PlanMedi – Plantago media L., PlatBifo – Platanthera bifolia (L.) Rich., PoaPrat – Poa pratensis L., PolyComo – Polygala comosa Schkuhr, PoteAnse – Potentilla anserina L., PrunVulg –Prunella vulgaris L., RanuAcri – Ranunculus acris L., RubuSaxa – Rubus saxatilis L., RumeConf – Rumex confertus Willd., SangOffi – Sanguisorba officinalis L., SilaSila – Silaum silaus (L.) Schinz & Thell., SoliVirg – Solidago virgaurea L., SoncArve – Sonchus arvensis L., StelGram – Stellaria graminea L., TanaVulg – Tanacetum vulgare L., TaraOffi – Taraxacum officinale F.H. Wigg., TrifPrat – Trifolium pratense L., TrolEuro – Trollius europaeus L., TussFarf – Tussilago farfara L., VeroCham – Veronica chamaedrys L., VeroSerp – Veronica serpyllifolia L., ViciCrac – Vicia cracca L., ViciSepi – Vicia sepium L., ViolMira – Viola mirabilis L. Деревья и кустарники: PinuSilv – Pinus sylvestris L., PiceAbie – Picea abies (L.) H. Karst., BetuPube – Betula pubescens Ehrh., BetuPend – B. pendula Roth, PopuTrem – Populus tremula L., AlnInc – Alnus incana (L.) Moench, SorbAucu – Sorbus aucuparia L., PaduAviu – Padus avium Mill., JuniComm – Juniperus communis L., Rosa maja – Rosa majalis Herrm., SaliAuri – Salix aurita L., SaliCapr – S. caprea L., SaliAcut – S. acutifolia Willd.

Полученная графическая интерпретация упорядочивания видов флористического списка в целом отражает общую закономерность, характерную для начальных стадий формирования залежных сообществ: большинство видов на графике локализуется в пределах одной общей области. Для залежей, находящихся на луговой рыхлокустовой стадии (по соотношению жизненных форм – см. рис. 3), это ожидаемо, так как парцеллярная структура лесного фитоценоза находится еще в самой начальной стадии формирования. К такому состоянию залежь пришла постепенно в течение 15 лет в результате нескольких процессов. С одной стороны, длиннокорневищевые травы (Elytrigia repens, Bromopsis inermis и др.) с течением времени достигли развития и, постепенно заполняя корневищами почвенный слой, уплотнили почву, в результате чего затруднился доступ воздуха в почву, повысилось увлажнение, длиннокорневищевые злаки в этих условиях стали слабее развиваться и постепенно начали выпадать из травостоя. С другой стороны, длиннокорневищные растения сами по себе имеют слабую конкурентную мощность и вытесняются рыхлодерновинными злаками, короткокорневищными видами разнотравья, ползучими и куртиноообразующими бобовыми. На месте корневищных видов трав на заброшенном поле Каргопольской суши быстро появляются более конкурентноспособные и менее требовательные к воздуху и почве рыхлокустовые злаки: Dactylis glomerata, Festuca pratensis, Agrostis tenuis, Phleum pratense. Одновременно в травостое увеличилась доля видов из семейства бобовых, в первую очередь Trifolium pratense.
Свой вклад в процесс разделения экологических ниш вносят сложная история поля и неоднородность экологических условий, связанная с ландшафтными особенностями поля, в частности на ординационной диаграмме (см. рис. 5) выделяются несколько обособленных групп. Левее от основного скопления точек, символизирующих виды, отделяются виды, которые в начале трансекты (точки апробирования 1–5) характеризуются высокой встречаемостью (Geranium sylvaticum, Sanguisorba officinalis, Solidago virgaurea, Platanthera bifolia, Trollius europaeus, Equisetum arvense, Anthriscus sylvestris). Исходя из эколого-биологических особенностей данных травянистых видов, численности древесно-кустарниковых видов в этой части трансекты, близости стены леса можно заключить, что данный залежный участок в пределах поля быстрее перейдет в лесную стадию.
Одна обособленная группа видов (Aegopodium podagraria, Tanacetum vulgare, Cirsium arvense, Bunias orientalis, Veronica serpyllifolia) и вторая группа (Juncus bufonius, Polygala comosa, Euphrasia brevipila) представлены сорно-придорожными видами и видами нарушенных местообитаний: через обследуемое поле проходит несколько зарастающих дорог, встречаются кротовины и разрытые муравейники.
Развитие лесного сообщества на изучаемом нами заброшенном поле находится в начале сукцессионного ряда по отношению к формированию зональных типов растительности. Именно с этим может быть связана недостаточная четкость флористической структуры на ординационной диаграмме. В климаксовых лесных сообществах роль типа биогеоценоза и его парцеллярного строения стабилизирована [52], и парцеллы, а в их пределах и тессеры значительно связаны со свойствами почв, отражая состав растительности [51–53]. На ранних этапах сукцессии при формировании горизонтального строения ценоза, когда еще не стабилизирован древесный ярус леса, идет смена растительности открытого луга на лесную, расселение растительности и ее мозаичность могут быть связаны не только с гетерогенностью свойств почв, но и определяться опадом и комплексом микроорганизмов, прежде всего в формирующейся лесной подстилке [51]. Из почвенных элементов питания при формировании гетерогенности растительности и ее кластеризации на группировки (парцеллы, тессеры) по мнению ряда авторов большую роль играют азот и углерод [31], азот и фосфор [53]. Стратегии расселения отдельных видов в связи с гетерогенностью почв пока изучены недостаточно и определяются биолого-экологическими особенностями видов и специфической потребностью в элементах минерального питания.
ЗАКЛЮЧЕНИЕ
При сельскохозяйственном использовании в почве формируется пахотный горизонт, который нивелирует генетические свойства почв и почвообразующих пород в пределах поля. Постоянная пахота, внесение удобрений и мелиорантов, использование севооборота, поддерживающих плодородие почв в этот период, приводят к формированию горизонта с относительно однородными свойствами, которые сохраняются и после отчуждения поля и начала восстановительной сукцессии. Формирование растительного покрова на изученной залежи в лесной зоне происходит в результате расселения древесных и кустарниковых видов от стен леса, поселения видов травянистой растительности за счет распространения зачатков от ближайших ценопопуляций (на опушках, полянах, ранее заросших полях), а также за счет диаспор сорной и культурной растительности, встречающейся или используемой в севооборотах, и их постоянной конкурентной борьбы.
Широкие спектры возможностей расселения растительности формируют ее смешанный характер, однако в первые 15 лет восстановительной сукцессии, после отчуждения поля из активного сельскохозяйственного пользования, наибольшую фитоценотическую значимость имеет травянистая растительность, формируя луговые ценозы. Поселение древесных и кустарниковых видов в первые 15 лет существенно не влияет на пул травянистой растительности и не способствует образованию локальных (территориальных) группировок. Гетерогенность почвенных свойств и более высокое плодородие верхнего (пахотного) горизонта почв, по сравнению с почвами нативных лесных насаждений, способствуют формированию широкого спектра экологических ниш, не ограничивающих экотопические аспекты формирования мозаики растительности на залежах. На этом этапе, через 15 лет залежеобразования, в условиях остаточно˗ карбонатных почв лесной зоны, специализация экологических свойств, как приуроченность видов к определенным факторам, проявляется слабо и только у отдельных травянистых видов. Факторами, определяющими формирование мозаичности растительного покрова на залежи, являются не изменчивость почвенных свойств и влияние поселившихся древесных и кустарниковых пород, которые со временем должны усилить свое влияние за счет разрастания крон и распространения корней, а биологические свойства видов травянистых растений, их специфические потребности к экологическим факторам.
Главными факторами, обусловливающими варьирование растительности на этой стадии сукцессии, являются плодородие почв и увлажнение. Именно на них реагируют виды травяного покрова 15-летней залежи в условиях Каргопольской суши. Древесно-кустарниковые виды менее дифференцированы в отзывчивости на мозаичность почвенных свойств на залежи из-за более широкой экологической амплитуды и отзывчивости на повышенное плодородие почв на залежном поле. На специфику почвенного питания, в частности насыщенность азотом, отзываются виды кустарников, входящие в пул подлесочных пород.
Исследования поддержаны грантом РФФИ и правительства Архангельской области № 17-44-290111.
Авторы заявляют об отсутствии конфликта интересов.
Настоящая статья не содержит исследований с участием людей или животных в качестве объектов изучения.
Список литературы
Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / Под ред. Смирновой О.В., Шапошникова Е.С. СПб.: РБО, 1999. 549 с.
Ettema Ch.H., Wardle D.A. Spatial soil ecology // Trends in ecology & evolution. 2002. V. 17. № 4. P. 177–183. https://doi.org/10.1016/S0169-5347(02)02496-5
Zhou Z., Sun O.J., Luo Z. et al. Variation in small-scale spatial heterogeneity of soil properties and vegetation with different land use in semiarid grassland ecosystem // Plant and Soil. 2008. V. 310. № 1. P. 103–112. https://doi.org/10.1007/s11104-008-9633-1
Townsend D.E., Fuhlendorf S.D. Evaluating relationships between spatial heterogeneity and the biotic and abiotic environments // The American Midland Naturalist. 2010. V. 163. № 2. P. 351–365. https://doi.org/10.1674/0003-0031-163.2.351
Лебедева В.Х., Ипатов В.С., Тиходеева М.Ю. Неоднородность пространственной структуры живого напочвенного покрова в лесных сообществах // Вестник СПбГУ. Сер. 3. Биология. 2015. № 2. С. 32–46.
Moreira E.F., Boscolo D., Viana B. Spatial Heterogeneity Regulates Plant-Pollinator Networks across Multiple Landscape Scales // PLoS ONE. 2015. V. 10. № 4. P. 1–19. https://doi.org/10.1371/journal.pone.0123628
Медведев В.В. Неоднородность как закономерное проявление горизонтальной структуры почвенного покрова // Грунтознавство. 2010. Т. 11. № 1–2. С. 6–15.
Медведев В.В., Мельник А.И. Неоднородность агрохимических показателей почвы в пространстве и во времени // Агрохимия. 2010. № 1. С. 20–26.
Мирин Д.М. Внутрифитоценозные элементы неоднородности растительного покрова // Изв. Самарского НЦ РАН. 2012. Т. 14. № 1(5). С. 1320–1323.
Витковская С.Е. Особенности пространственной неоднородности агрохимических показателей почвы и массы растений в полевом опыте // Плодородие. 2009. № 5. С. 8–9.
Гончаров В.М., Фаустова Е.В. Новые подходы к исследованию пространственной агрофизической неоднородности почвенного покрова // Вестник Оренбургского гос. ун-та. 2011. № 12 (131). С. 181–183.
Басевич В.Ф. Почвенные сукцессии и их связь с неоднородностью подзолистых почв // Вестник Московского ун-та. Сер. 17. Почвоведение. 2011. № 3. С. 38–42.
Юркевич М.Г. Горизонтальная структура агроценозов Южной Карелии // Экология и география почв / Под ред. Красильникова П.В. Петрозаводск, 2009. С. 105–115.
Al-Rowaily S., El-Bana M., A.R. Al-Dujain F. Changes in vegetation composition and diversity in relation to morphometry, soil and grazing on a hyper-arid watershed in the central Saudi Arabia // Catena. 2012. V. 97. P. 41–49. https://doi.org/10.1016/j.catena.2012.05.004
ванов А.И., Конашенков А.А., Хомяков Ю.В. и др. Оценка параметров пространственной неоднородности показателей плодородия дерново-подзолистых почв // Агрохимия. 2014. № 2. С. 39–49
Ntasin A., Sompongchaiyakul P., Singhruck P. Soil Nutrients in heterogeneities land use in Haui-Luang Headwater Area of the Mekong River // Journal of Environmental Management and Tourism. 2017. V. 8. № 5. P. 1006–1012. https://doi.org/10.14505//jemt.v8.5(21).04
Сидорова В.А. Изменение пространственной вариабельности почвенных свойств в результате антропогенного воздействия // Экология и география почв / Под ред. Красильникова П.В. Петрозаводск, 2009. С. 30–47.
Paz-González A., Vieira S.R., Taboada Castro M.T. The effect of cultivation on the spatial variability of selected properties of an umbric horizon // Geoderma, 2000. V. 97. P. 273–292.
Литвинович А.В. Пространственная неоднородность агрохимических показателей пахотных дерново-подзолистых почв // Агрохимия. 2007. № 5. С. 89–94.
Okayasu T., Okuro T., Jamsran U., Takeuchi K. Degraded rangeland dominated by unpalatable forbs exhibits large-scale spatial heterogeneity // Plant Ecology. 2012. V. 213. № 4. P. 625–635. https://doi.org/10.1007/s11258-012-0027-3
Dechesne A., Badawi N., Aamand J., Smets B.F. Fine scale spatial variability of microbial pesticide degradation in soil: scales, controlling factors, and implications // Frontiers in Microbiology. 2014. V. 5. № 667. https://doi.org/10.3389/fmicb.2014.00667
Seabloom E.W., Bjørnstad O.N., Bolker B.M., Reichman O.J. Spatial signature of environmental heterogeneity, dispersal, and competition in successional grasslands // Ecological Monographs. 2005. V. 75. № 2. P. 199–214. https://doi.org/10.1890/03-0841
Kumar S., Tohlgren T.J., Chong G.W. Spatial heterogeneity influences native and nonnative plant species richness // Ecology. 2006. V. 87. № 12. P. 3186–3198. https://www.jstor.org/stable/20069347
DeAngelis D.L., Yurek S. Spatially explicit modeling in ecology: a review // Ecosystems. 2017. V. 20. № 2. P. 284–300. https://doi.org/10.1007/s10021-016-0066-z
Смирнова О.В., Алейников А.А., Семиколенных А.А. и др. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике // Лесоведение. 2011. № 6. С. 67–78.
Wang L., Okin G.S., D’Odorico P. et al. Ecosystem-scale spatial heterogeneity of stable isotopes of soil nitrogen in African savannas // Landscape Ecology. 2013. V. 28. № 4. P. 685–698. https://doi.org/10.1007/s10980-012-9776-6
Chen P., Chu Y., Gu F. et al. Spatial heterogeneity of vegetation and soil characteristics in oasis-desert ecotone // The Journal of Applied Ecology. 2003. V. 14. № 6. P. 904–908.
Schooley R.L. Spatial heterogeneity and characteristic scales of species-habitat relationships // BioScience. 2006. V. 56. № 6. P. 533–537. https://doi.org/10.1641/0006-3568(2006)56[533:shacso]-2.0.co;2
Davies K.F., Chesson P., Harrison S. et al. Spatial heterogeneity explains the scale dependence of the native-exotic diversity relationship // Ecology. 2005. V. 86. № 6. P. 1602–1610. https://www.jstor.org/stable/3450785
Zhukov A., Gadorozhnaya G. Spatial heterogeneity of mechanical impedance of atypical chernozem: the ecological approach // Ekológia (Bratislava). 2016. V. 35. № 3. P. 263–278. https://doi.org/10.1515/eko-2016-0021
Солодовников А.Н., Рожков В.А. Исследование влияния древесной породы на почву методом дискриминантного анализа // Бюлл. Почвенного института им. В.В. Докучаева. 2019. Вып. 96. С. 22–38. https://doi.org/10.19047/0136-1694-2019-96-221-46
Москаленко С.В., Бобровский М.В. Расселение лесных видов растений из старовозрастных дубрав на брошенные пашни в заповеднике “Калужские засеки” // Изв. Самарского НЦ РАН. 2012. Т. 14. № 1(5). С. 1332–1335.
Vellend M., Verheyen K., Kathryn M. et al. Homogenization of forest plant communities andweakening of species–environment relationships via agricultural land use // J. of Ecology. 2007. V. 95. № 3. P. 565–573. https://doi.org/10.1111/j.1365-2745.2007.01233.x
Macdonald D.V., Crabtree J.R., Wiesinger G. et al. Agricultural abandonment in mountain areas of Europe: environmental consequences and policy response // J. of Environmental Management. 2000. V. 59. № 1. P. 47–69. https://doi.org/10.1006/jema.1999.0335
Розанов Б.Г. Морфология почв: Учебное пособие для вузов по спец. “Почвоведение и агрохимия”. М.: Изд-во МГУ, 1983. 320 с.
Дмитриев Е.А. Глава из неоконченной книги “Неоднородность почвы” // Масштабные эффекты при исследовании почв. М.: Изд-во МГУ, 2001. С. 8–38.
Горячкин С.В. Почвенный покров Севера (структура, генезис, экология, эволюция). М.: ГЕОС, 2010. 414 с.
Geypens M., Vanongeval L., Vogels N., Meykens J. Spatial variability of agricultural soil fertility parameters in a gleyic podzol of Belgium // Precision Agriculture. 2004. V. 1. P. 319–326. https://doi.org/10.1023/A:1009977109430
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
Понятовская В.М., Сырокомская И.В. Опыт сравнительной оценки участия вида в строении лугового сообщества // Труды БИН АН СССР. Сер. III.Геоботаника. Л.: АН СССР, 1960. Вып. 12. С. 127–139.
Смирнова О.В., Ханина Л.Г., Смирнов В.Э. Эколого-ценотические группы в растительном покрове лесного пояса Восточной Европы // Восточно-европейские леса: история в голоцене и современность / Под ред. Смирновой О.В. СПб., 2004. Кн.1. С. 165–175.
Тиходеева М.Ю., Лебедева В.Х. Практическая геоботаника (анализ состава растительных сообществ). СПб.: Изд-во С.-Петерб. ун-та, 2015. 166 с.
McCune B., Mefford M.J. PC-ORD Multivariate Analysis of Ecological Data, Version 6. Gleneden Beach, Oregon: MjM Software Design. 2011. 28 p.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Грохлина Т.И., Ханина Л.Г. О компьютерной обработке геоботанических описаний по экологическим шкалам // Математическое моделирование в экологии: Мат-лы 4-й нац. науч. конф. с международн. участием. Отв. ред. Комаров А.С. Пущино, 2015. С. 63–64.
Oksanen J., Blanchet F., Friendly M. et al. 2017. vegan: Community Ecology Package. R package version 2.4-4.
Витковская С.Е. Методы оценки неоднородности почвенного покрова при планировании и проведении полевых опытов. СПб.: Агрофизический институт, 2011. 52 с.
Басевич В.Ф., Тетенькин В.Л. Неоднородность подзолистых почв и пестрополье // Вестник Московского ун-та. Сер. 17. Почвоведение. 2010. № 2. С. 35–42.
Лебедева В.Х., Тиходеева М.Ю. О неоднородности лесной и болотной растительности Нижне-Свирского заповедника // Бот. журн. 2017. Т. 102. № 6. С. 746–767.
Тиходеева М.Ю., Лебедева В.Х., Панфиловская К.А. Классификация типов зарастания суходольных лугов // Сборник научных трудов ГНБС. 2016. Т. 143. С. 242–248.
Орлова М.А., Лукина Н.В., Камаев И.О. и др. Мозаичность лесных биогеоценозов и продуктивность почв // Лесоведение. 2011. № 6. С. 39–48.
Роль почвы в формировании и сохранении биологического разнообразия / Отв.ред. Добровольский Г.В., Чернов И.Ю. М.: Тов-во научн. изд. КМК, 2011. 273 с.
Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги / Отв. ред. Федорец Н.Г. М.: Наука, 2006. 287 с.
Дополнительные материалы отсутствуют.