

Журнал эволюционной биохимии и физиологии, 2020, T. 56, № 4, стр. 303-312
НЕИНВАЗИВНОЕ ЭЛЕКТРОФИЗИОЛОГИЧЕСКОЕ ИССЛЕДОВАНИЕ ТАКТИЛЬНОЙ ЧУВСТВИТЕЛЬНОСТИ У КАРПОВЫХ РЫБ (CYPRINIDAE)
Г. В. Девицина 1, *, Д. Н. Лапшин 2
1 Московский государственный университет им. М.В. Ломоносова
Москва, Россия
2 Институт проблем передачи информации им. А.А. Харкевича Российской академии наук
Москва, Россия
* E-mail: gdevicyna@mail.ru
Поступила в редакцию 17.01.20
После доработки 25.03.20
Принята к публикации 27.03.2020
Аннотация
Тактильную рецепцию карповых рыб (серебряный карась, карп обыкновенный) исследовали методом неинвазивной регистрации рецепторных ответов на поверхности кожи головы в ответ на ритмическую тактильную стимуляцию. Ответные реакции в виде электрических потенциалов, следовавших синхронно с изменением силы давления на кожу, регистрировали с помощью хлор-серебряного электрода. Показано, что рыбы, также как и наземные позвоночные, обладают тактильными рецепторами фазного и тонического типа. Зарегистрированы три основные формы тактильных реакций: тонические ответы в виде колебаний потенциала, повторяющих динамику тактильного стимула, фазные ответы в виде on-, off- и on–off потенциалов, а также медленное отклонение от потенциала покоя. Наблюдались также суммарные реакции, одновременно включающие несколько форм ответов. Латентный период ответов тонического типа составлял в среднем 6.1 мс, фазных ответов – 4.4 мс и явным образом не зависел от изменения направления (включение и выключение) стимула. Пороговый уровень тактильной чувствительности в околоротовой и гулярной зонах головы составлял 0.05–0.2 г/мм2. Показано, что распределение тактильных рецепторов на голове карася и карпа носит зональный характер, а сенсорные зоны различаются по тактильной чувствительности. Наиболее чувствительные к тактильным стимулам зоны расположены на верхней и нижней губе, в коже около губ, а также в гулярной зоне на вентральной стороне головы. Менее чувствительными к тактильным стимулам были зоны, расположенные ростральнее и вентральнее глазного яблока. Зоны, находящиеся дорсальнее и каудальнее глазного яблока, оказались нечувствительными к применяемой тактильной стимуляции. Во всех сенсорных зонах головы рыб наиболее интенсивные реакции вызывал скользящий тактильный стимул. Специфика распределения тактильных сенсорных зон соответствует их функциональному значению в пищевом поведении рыб.
ВВЕДЕНИЕ
Кожные покровы рыб содержат многочисленные сенсорные структуры различной специализации, включающей хеморецепцию, механорецепцию и электрорецепцию [1–3]. Рыбы, обитающие в условиях низкой освещенности, активно используют тактильную рецепцию в различных формах поведения. Особенно большую роль играет тактильная рецепция в системе ориентации обитателей придонных биотопов и пещерных водоемов [3–6]. У позвоночных животных информационное значение тактильной рецепции в условиях низкой освещенности сопоставимо со значением зрительной рецепции [7, 8]. Тактильная рецепция позвоночных, являясь элементом сомато-висцеральной системы, не имеет специализированных сенсорных органов. Чувство осязания или тактильная чувствительность диффузно распределяется в кожных покровах тела и головы рыб.
До настоящего времени порядок формирования тактильной информации на рецепторном уровне у рыб оставался почти неисследованным разделом сенсорной физиологии. Большинство электрофизиологических исследований было посвящено в первую очередь химической чувствительности рыб, тогда как вызванные тактильные реакции на ненормированные механические стимулы продемонстрированы авторами только в качестве феномена. Ответы на механическую стимуляцию хемосенсорных структур в коже рыб в этих работах были зарегистрированы в волокнах тройничного и лицевого нервов, а также в ядрах соответствующих проекционных зон головного мозга [9–11]. Отсутствие данных о чувствительности и других физиологических характеристиках тактильной рецепции рыб можно объяснить, в частности, ограниченным набором методик, позволяющих дозированно осуществлять адекватную стимуляцию.
В задачи данной работы входили: 1) регистрация и определение параметров электрических ответов в коже рыб на нормированную тактильную стимуляцию, 2) определение пороговой величины тактильного стимула, 3) выявление специфических типов кожных тактильных рецепторов, 4) анализ распределения тактильной чувствительности на голове рыб.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
В исследовании использовали 12 особей серебряного карася (Carassius auratus gibelio) длиной 9–12 см и 5 особей карпа (Cyprinus carpio) длиной 12–15 см. Подопытных рыб содержали в аквариуме объемом 50 л. В качестве корма использовали личинок комаров (Chironomidae) и сухой корм. Перед экспериментом рыбу закрепляли в пластмассовом боксе 27 × 9 × 5 см таким образом, что все ее тело находилось в воде за исключением участка головы, выбранного для тестирования.
Для регистрации рецепторных потенциалов необходима чистая поверхность кожи, лишенная чешуи. В связи с этим тактильную чувствительность регистрировали только на голове рыбы. Жабры подопытной рыбы постоянно перфузировали водой при 18–19°С. Иммобилизацию рыбы обеспечивали большой скоростью (150 мл/мин) водного протока. Миорелаксанты не применяли. После завершения исследования рыбы, отпущенные в аквариум, продолжали обычным образом плавать, питаться и могли быть использованы в последующих экспериментах.
Для измерения тактильной чувствительности была применена апробированная ранее методика неинвазивной регистрации электрических потенциалов на поверхности кожи головы рыбы [12, 13]. Этот метод позволял последовательно в ходе одного опыта тестировать участки кожи на дорсальной, латеральной и вентральной сторонах головы без повреждения кожной поверхности.
Тактильную стимуляцию осуществляли упругими монофиламентами (МФ) из нейлона длиной 30–50 мм, загнутыми в терминальной части. Каждый МФ имел оплавленный торец в виде небольшой сферы диаметром 1 мм. МФ крепили в зажиме электромагнитной системы (тактильный стимулятор, рис. 1), которая под управлением сигналов от цифро-аналогового преобразователя (ЦАП) последовательно четыре раза в ходе одной регистрации направляла МФ в сторону исследуемого участка кожи подопытной рыбы. Цикл стимуляции, состоящий из 4 стимулов, изображен на рис. 1. Длительность фронтов срабатывания электромагнитной системы в процессе стимуляции составляла 4 мс, время запаздывания начала движения относительно фронтов управляющего электрического импульса не превышало 1 мс.
Рис. 1.
Схема экспериментальной установки для измерения тактильной чувствительности рыб. На врезке показан график изменения давления на кожу со стороны тактильного стимулятора. Стрелки, направленные вниз, соответствуют моментам увеличения давления на кожу со стороны стимулятора, стрелки, направленные вверх, – снижению давления.
Fig. 1. A setup for measuring tactile sensitivity in fish. The stimulus is applied onto the skin by a computer-controlled tactile stimulator. The electrical response is recorded from the skin through the electrode. Inset diagram shows the sequence of a single stimulation episode. Downward arrows indicate an increase in pressure on the skin, upward arrows show a pressure relief. ADC – analog-to-digital converter; PC – personal computer; DAC – digital-to-analog converter.

Всего было использовано шесть типов МФ разной жесткости. Усилие, оказываемое на поверхность кожи в процессе стимуляции торцом наиболее мягкого МФ, составляло 0.04 г, усилие наиболее жесткого МФ – 1.3 г. С учетом площади контакта сферического торца МФ 0.8 мм2 этим усилиям соответствовало давление в области контакта 0.05 и 1.6 г/мм2. Монофиламенты, обеспечившие промежуточные ступени силы стимуляции, были отрегулированы таким образом, чтобы при переходе на очередную ступень в сторону увеличения усилие давления на кожную поверхность возрастало в два раза (0.08, 0.16, 0.32, 0.64 г). Усилие, развиваемое каждым образцом МФ при срабатывании стимулирующей системы, калибровали с помощью весов HL-100 (A&D, Япония). В процессе подготовки к экспериментам усилие, развиваемое МФ, регулировали выбором диаметра МФ (грубый подбор) и изменением его длины (точный подбор).
В качестве регистрирующего электрода использовали хлор-серебряную проволоку диаметром 0.3 мм, заключенную в стеклянный капилляр. Объем между электродом и внутренней стенкой капилляра был заполнен водой из аквариума. Оплавленный торец капилляра во время регистрации плотно, без просветов прилегал к поверхности кожи. Сигнал с электрода поступал на вход усилителя постоянного тока с входным сопротивлением 1 ГОм, частотной полосой до 2 кГц и системой автоматической компенсации нуля на выходе, программно активировавшейся в интервалах между регистрациями. Усиленный сигнал передавался на один из входов аналогово-цифрового преобразователя (АЦП Е14-440 “Л-Кард”, Россия). Одновременно по другому каналу АЦП производилась регистрация управляющего сигнала электромагнитной системы подачи МФ. После преобразования в цифровой код с частотой квантования 20 кГц эти данные фиксировали в памяти компьютера. АЦП функционировало под управлением программы Lgraph II, разработанной фирмой Л-Кард (Россия).
Последовательность цикла движений стимулирующего МФ определялась программно. Каждая регистрация длилась 5.8 с и включала в себя интервал фоновой активности (1.8 с до начала стимуляции), ответы на четырехкратную механическую стимуляцию исследуемого участка кожи (1.4 с) и вариации потенциала в последействии (2.6 с). В процессе первоначального поиска участков кожи, механическая стимуляция которых вызывала электрический ответ, использовали МФ, создававший при нажатии усилие 1.3 г. Интервал между последовательными циклами стимуляции составлял не менее 8 с.
До начала стимуляции торец МФ без нажима касался исследуемого участка кожи. Взаимное расположение точек электрической регистрации и точек приложения тактильных стимулов зависело от анатомии исследуемого участка головы. Например, при исследовании чувствительности губ электрод устанавливали на верхней или нижней губе, затем стимулировали сами губы и ближайшие к ним участки кожи. В процессе исследования других зон торец электрода чаще всего располагали в пределах исследуемой области (рис. 2).
Рис. 2.
Схематичное представление результатов тестирования латеральной стороны околоротовой области у карася, Carassius auratus gibelio (данные одного опыта). Обозначения: крест – место контакта торца регистрирующего электрода с поверхностью кожи; мелкими ромбами обозначены участки на поверхности кожи, стимуляция которых вызывала ответные реакции.
Fig. 2. Schematic representation of data from a single experiment on testing the lateral perioral area of the crucian carp, Carassius auratus gibelio. The position of the recording electrode is indicated by a cross. The points on the skin, tactile stimulation of which elicited responses, are shown with diamond marks.
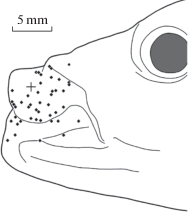
Для визуализации исследуемой области головы рыбы использовали видеокамеру CNB-ZBN-21Z27F (CNB, Китай). Точки стимуляции фиксировали в полярных координатах относительно места прилегания к коже торца отводящего электрода. Расстояние между торцом отводящего электрода и точками тактильной стимуляции варьировали в пределах от 1 до 15 мм.
Для определения тактильной чувствительности в каждой исследованной зоне измеряли пороговую силу механического давления на поверхность кожи, вызывавшую отклонение регистрируемого потенциала от базовой линии на величину не менее 0.1 мВ.
Порог реакции определяли в интервале между двумя значениями стимула при условии, что бòльший по амплитуде стимул вызывал реакцию, тогда как при действии более слабого стимула реакция в той же области кожи не наблюдалась.
Полярность электрических ответов считали “прямой” при положительном сдвиге потенциала в ответ на увеличение давления стимулятора на кожу. Если при увеличении механического усилия мы наблюдали отклонение регистрируемого потенциала в сторону отрицательных значений, то такую реакцию назвали “инверсной”. При снижении давления стимулятора на кожу “прямой” ответ выглядел как отклонение потенциала в сторону отрицательных значений, тогда как “инверсный” ответный потенциал изменялся в сторону положительных значений.
Всего было проведено 32 эксперимента, включавших в сумме 2500 регистраций. Из этого числа в 1200 случаях наблюдались реакции на тактильную стимуляцию.
Больше половины от общего числа регистраций (1300) не содержали признаков ответа на стимул. В эту группу входили контрольные регистрации потенциала при движении МФ около головы рыбы, без механического контакта с кожей и результаты стимуляции тактильно нечувствительных либо малочувствительных участков кожи.
При анализе данных определяли экспериментальную вероятность (pe) появления реакций на стимулы в пределах исследуемого участка кожи как отношение числа зарегистрированных ответов к числу предъявлений стимулов.
Измерение латентных периодов и длительности фронтов нарастания реакций в ответ на тактильные стимулы проводили после опытов с помощью программы SoundForge 10 Pro (Sony, Япония). Моментом начала ответа считали уровень 0.1 от его максимума; соответственно, момент начала спада отсчитывали от уровня 0.9 от предшествовавшего максимума электрического ответа.
Чтобы подтвердить рецепторную природу регистрируемых реакций, мы провели серию из шести контрольных опытов с применением лидокаина – блокатора потенциал-зависимых натриевых каналов [14]. Использовали либо местное нанесение 2.0% раствора лидокаина на кожу с помощью небольшого ватного тампона, либо введение подкожно в исследуемой зоне головы 20 мкл раствора лидокаина. Активность тактильных рецепторов регистрировали до аппликации лидокаина и повторяли через 4–5 мин. после аппликации. Далее, в течение следующих 20 мин. наблюдали динамику изменения параметров электрического ответа на тактильную стимуляцию до возвращения амплитуды ответной реакции к исходному уровню.
РЕЗУЛЬТАТЫ
Электрические ответы на тактильную стимуляцию
В ходе экспериментов были зарегистрированы три типа реакций: тонические ответы в виде колебаний потенциала, повторяющих динамику четырехкратного стимула (рис. 3A, B), фазные ответы в виде потенциалов on-, off- и on–off (рис. 4A, B, C), а также медленные потенциалы с общей длительностью, превышающей суммарное время цикла стимуляции. Были зарегистрированы также комплексные ответы, в которых сочетались реакции рецепторов тонического и фазного типа в сумме с медленными потенциалами (рис. 3C, D; рис. 4C). Вероятность регистрации ответов тонического и фазного типа была сходной (44% и 37% соответственно). Доля медленных ответов составляла 19% от общего числа реакций. Полярность тонических и фазных ответов могла быть прямой и инверсной (рис. 3A, B) относительно циклов надавливания МФ на кожу рыбы в процессе стимуляции.
Рис. 3.
Тонические реакции, зарегистрированные в ответ на четырехкратную тактильную стимуляцию верхней губы карася (Carassius auratus gibelio):
A – прямая форма ответа (срединная часть губы); B – инверсная форма ответа (срединная часть губы); C – нарастание амплитуды переменной составляющей ответа на фоне суммации с медленным потенциалом (латеральная часть губы);
D – постепенное уменьшение амплитуды переменной составляющей ответа на фоне суммации с медленным потенциалом (там же); E – временнáя схема включения и выключения тактильного стимулятора в ходе одной регистрации. Черными прямоугольниками обозначены интервалы действия тактильных стимулов длительностью 0.2 с каждый.
Fig. 3. DC (direct current) responses to quadruple alternating tactile stimulation of the crucian carp (Carassius auratus gibelio) upper lip. A – direct form of response (middle part of the lip); B – inverted form of response (middle part of the lip); C – summation of the sustained component of the response with gradually increasing alternating component (lateral part of the lip); D – the same, but with decreasing alternating component. E – sequence of stimulating episodes: black bars indicate the individual tactile stimuli, each of 0.2 s in duration.
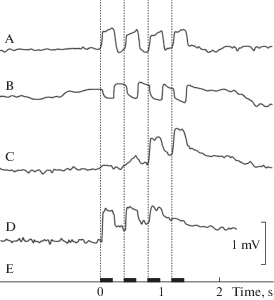
Рис. 4.
Примеры фазных ответов на тактильную стимуляцию вентральной поверхности головы карася (Carassius auratus gibelio):
A – реакции on–off ; B – реакции (on) только на передние фронты тактильных стимулов (на увеличение механического давления на кожу) с признаками суммации с тонической составляющей на уровне четвертого такта; C – последовательность однополярных ответов на все фронты переключения механического стимула в сумме с медленно нараставшим потенциалом; D – негативные on- реакции в сочетании с возрастающими по амплитуде off- реакциями на спады усилия тактильного раздражителя; E – аналогично рис. 3.
Fig. 4. Examples of phasic (AC) responses to tactile stimulation of the ventral side of a crucian carp (Carassius auratus gibelio) head. A – on-off responses; B – on-responses only to the moments of pressure increase, with some signs of summation with the DC component of the response at the fourth stimulus level; C – unipolar on- and off-responses summed with slowly rising sustained response potential; D – negative on-response together with increasing off-responses (to the moments of pressure relief); E – stimulation, see Fig. 3.

В ответах тонического типа амплитуда потенциалов, следовавших синхронно с тактами стимуляции, чаще всего была постоянной, но в ряде опытов наблюдались как постепенное возрастание амплитуды ответа (рис. 3C), так и плавное уменьшение амплитуды ответной реакции (рис. 3D).
Фазные ответы наблюдались в нескольких вариантах и сочетаниях типов реакций: выбросы потенциала на увеличение и на последующие спады механического давления на кожу (рис. 4A); реакции только на передние фронты тактильных стимулов, иногда с признаками суммации с тонической составляющей ответа (рис. 4B); реакции в виде однополярных положительных ответов на все фронты переключения механического стимула, независимо от их направления и в сумме с медленным потенциалом (рис. 4C); фазные реакции негативной полярности на фронты нарастания и спада усилия тактильного раздражителя (рис. 4D).
Аналогично процессам, сопровождавшим тонические реакции, амплитуда коротких потенциалов в фазных ответах могла меняться в процессе стимуляции – возрастать, либо уменьшаться. Многократная стимуляция одной тактильно чувствительной точки одинаковыми дозированными стимулами, как правило, приводила к снижению амплитуды ответа и выпадению отдельных его компонентов.
Особенно сильные реакции тонических и фазных типов возникали при скользящем движении торца стимулирующего МФ по поверхности кожи. Однако это явление можно отметить только в качестве феномена, так как нормировать физические параметры такой стимуляции в условиях применяемой методики не представлялось возможным.
Латентный период электрических реакций на тактильную стимуляцию
Латентный период фронта нарастания тонических ответов относительно стимулов составлял в среднем 6.1 мс (среднеквадратичное отклонение σ = 3.3 мс), задержка фронтов спада соответственно 9.8 мс (σ = 6.0 мс). Какой-либо зависимости величины латентных периодов от номера отдельного стимула в четырехкратной последовательности не выявлено. Латентный период реакций фазных рецепторов на передние фронты стимулов в среднем составлял 4.4 мс (σ = 2.7 мс), а на задние фронты 4.1 мс (σ = 3.1 мс).
Влияние лидокаина на тактильные ответы
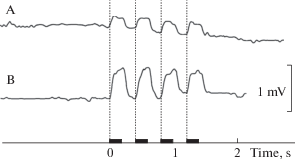
Аппликация лидокаина на поверхность исследуемого участка кожи с помощью ватного тампона или подкожное введение лидокаина вызывали сходную картину изменений в электрических ответах на тактильную стимуляцию во всех шести опытах этой серии. Наблюдалось резкое увеличение в несколько раз амплитуды регистрируемых ответов (p = 0.95 по Т-критерию Вилкоксона) через 2–3 мин после аппликации анестетика (рис. 5). Повышенная возбудимость в ответ на действие тактильных стимулов сохранялась в течение порядка 10 мин. Затем амплитуда ответов постепенно снижалась примерно за тот же интервал времени до уровня в несколько раз ниже исходного, а затем в течение 2–3 мин восстанавливалась до исходного уровня.
Рис. 5.
Увеличение амплитуды ответных потенциалов при действии лидокаина в исследуемой области кожи головы карпа (Cyprinus carpio):
A – до введения лидокаина; B – через пять минут после введения лидокаина. Область исследования и положение электрода аналогичны рис. 2. Другие обозначения аналогичны рис. 3.
Fig. 5. A lidocaine-induced increase in the amplitude of tactile responses in the test area of the carp (Cyprinus carpio) head. A – before lidocaine application; B – b5 min after lidocaine application. The area of tactile stimulation and electrode position the same as in Fig. 2. For other designations see Fig.3.
Зональное распределение тактильной чувствительности на поверхности головы рыб
Анализ распределения тактильной чувствительности на поверхности головы рыбы позволил выделить несколько сенсорных зон, в которых были зарегистрированы реакции на тактильный стимул. На латеральной стороне головы выделены зоны – околоротовая, назальная и глазничная (рис. 6A), на дорсальной стороне – околоротовая, назальная и межглазничная (рис. 6B), на вентральной стороне – околоротовая, гулярная и межжаберная (рис. 6C). Эти зоны различались по уровню чувствительности к тактильной стимуляции и по вероятности появления реакций при тестировании соседних участков кожи. Характер ответов на тактильную стимуляцию в каждой зоне головы рыбы имел свою специфику сочетаний реакций тонического, фазного и медленного типа.
Рис. 6.
Сенсорные зоны головы рыбы, в которых изучали тактильную рецепцию:
A – на латеральной стороне: околоротовая (1), назальная (2), глазничная (3);
B – на дорсальной стороне: околоротовая (1), назальная (2), межглазничная (3);
C – на вентральной стороне: околоротовая (1), гулярная (2), межжаберная (3).
На врезках для каждой стороны приведены гистограммы, иллюстрирующие экспериментальную вероятность (pe) регистрации реакций в пределах отдельных зон. Рядом с гистограммами указано общее количество предъявлений тактильных стимулов при тестировании одной из сторон (Carassius auratus gibelio).
Fig. 6. Sensory areas on the fish head (Carassius auratus gibelio), where the tactile reception was studied. A – on the lateral side: near-mouth (1), nasal (2), orbital (3); B – on the dorsal side: perioral (1), nasal (2), interorbital (3); C – on the ventral side: perioral (1), gular (2), inter-gill (3). Bar charts in the insets for each side show the probability of responses (pe) in each area. Numbers near the bar charts stand for the total number of tactile stimulation tests applied to each side.
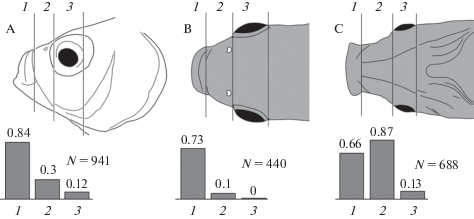
Высокая тактильная чувствительность и высокая вероятность реакций (pe = 0.84) была характерна для околоротовых зон, которые включают верхнюю, нижнюю губы и поверхность кожи около губ (рис. 2; рис. 6, зона 1 на каждой из сторон). В околоротовой зоне в латеральной части верхней губы регистрировались ответы фазного и тонического типа. В околоротовой зоне на дорсальной поверхности головы в центральной части верхней губы были зарегистрированы преимущественно медленные потенциалы, а также ответы тонического типа. Тактильная стимуляция верхней губы иногда вызывала двигательную реакцию тела и хвоста рыбы. Кожа около верхней губы проявляла также высокую тактильную чувствительность. Здесь регистрировались реакции на тактильные стимулы 0.04 г (0.05 г/мм2), которые проявлялись в форме тонического ответа или медленного колебания потенциала.
Латеральные участки нижней и верхней губы, контактирующие друг с другом, характеризовались особенно низкими порогами тактильной чувствительности. Здесь были зарегистрированы реакции тактильных рецепторов на давление 0.05 г/мм2. На вентральной стороне головы в коже около и каудальнее нижней губы также отмечена высокая чувствительность тактильных рецепторов тонического и фазного типа.
Распределение различных типов ответных реакций в околоротовых зонах (рис. 6, зона 2) на латеральной, дорсальной и вентральной стороне головы был сходным. В этих зонах ответы тонического и фазного типов представлены почти в равном соотношении, составляя 36%–41%. Встречались также ответы смешанного типа (фазно-тонические) и ответы в сочетании с медленным колебанием потенциала. Медленную форму ответа наблюдали преимущественно на дорсальной стороне головы, как в околоротовой (23%), так и в назальной (18%) зонах. На латеральной и вентральной стороне головы в околоротовых зонах медленная форма ответа проявлялась лишь в 10% регистраций. В назальной зоне на латеральной стороне головы соотношение тонического и фазного типов ответа было сходным с околоротовыми зонами: 46% представляли ответы тонического типа и 36% – фазного.
На латеральной стороне головы зоны назальная и глазничная (рис. 6A, зоны 2, 3) были значительно менее чувствительны к тактильным стимулам, чем зоны околоротовой локализации (рис. 6A, зона 1). Так, в назальной и глазничной зонах на латеральной стороне головы пороговые реакции в виде синхронных со стимулами сдвигов потенциала амплитудой более 0.1 мВ возникали в диапазоне давлений на кожу 0.2–1.6 г/мм2, тогда как в околоротовых зонах были зарегистрированы ответы при давлении от 0.05 до 0.2 г/мм2. В назальной зоне дорсальной стороны головы (рис. 6B, зона 2) тактильные реакции удалось зарегистрировать только при еще большем давлении на кожу – не менее 1.6 г/мм2. В межглазничной зоне на дорсальной стороне головы (рис. 6B, зона 3) тактильные реакции не выявлены.
На вентральной стороне головы каудальнее околоротовой зоны центральное положение занимает гулярная зона (рис. 6С, зона 2). Для рострального участка этой зоны характерна особенно высокая вероятность возникновения реакций на тактильную стимуляцию (pe = 0.87). В этой зоне рядом расположенные точки на поверхности кожи различались по пороговой чувствительности на одну–две градации. Судя по степени выраженности ответных реакций на минимальное стимулирующее воздействие, пороги чувствительности в некоторых точках этой зоны были ниже минимального уровня тактильного стимула 0.04 г (0.05 г/мм2).
В центральной части гулярной зоны наблюдались признаки одновременной активации тактильных рецепторов фазного и тонического типов. В отличие от глазничных зон на дорсальной и латеральной сторонах, в гулярной зоне преобладали ответы фазного типа. Ответы в разных точках гулярной зоны различались не только по форме, но и по амплитуде и длительности. В ростральной части гулярной зоны встречались участки, где были зарегистрированы только фазные ответы, иногда типа on–off. В каудальной части гулярной зоны регистрировались ответы тонических рецепторов чаще (51%), чем фазных (39%).
На поверхности наиболее латеральных участков гулярной зоны вероятность появления тактильных реакций (pe = 0.3) была примерно в три раза ниже по сравнению с ее центральной частью (pe = 0.87). Пороги реакций в этих областях также оказались в 2–4 раза выше по сравнению с центральной частью гулярной зоны. В коже межжаберной зоны (рис. 6С, зона 3) вероятность регистрации тактильных реакций низкая (pe = 0.13).
Таким образом, характер распределения параметров тактильной системы на поверхности головы рыб (карася и карпа) показывает тенденцию к снижению уровня чувствительности и вероятности возникновения электрофизиологических ответов на механические стимулы в ростро-каудальном и в вентро-дорсальном направлении.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
При анализе полученных результатов важно было решить вопрос о природе ответных потенциалов. Два фактора указывают на то, что зарегистрированная нами активность тонических и фазных рецепторов является отражением первичных электрических процессов (т.е. рецепторных потенциалов) в кожных рецепторах при действии механических стимулов: 1 – малые задержки ответов (в среднем 6.1 мс для тонических реакций и 4.1 мс для фазных) и 2 – характерное влияние на амплитуду вызванных потенциалов аппликации лидокаина, который является блокатором потенциал-зависимых натриевых каналов [14]. Увеличение амплитуды вызванных потенциалов при действии лидокаина означает, что источники этих потенциалов расположены дистально, т.е. до афферентных волокон, и источником потенциалов являются рецепторы. Зарегистрированные в наших опытах эффекты суммации фазных и тонических ответов можно объяснить особенно высокой плотностью распределения различных типов тактильных рецепторов в определенных участках кожи головы рыбы. Это подтверждают и морфологические исследования, которые отмечают чрезвычайно высокую плотность распределения нервных окончаний в поверхностных слоях кожного покрова, и особенно в сенсорных зонах околоротовой и вентральной локализации [15–17].
Такие особенности иннервации кожи головы рыб можно рассматривать как основу разнообразия форм регистрируемых нами ответных реакций на тактильные стимулы. Разнообразие форм ответных потенциалов характерно для различных типов механо-чувствительных клеток [18]. Реакции на увеличение механического давления либо реакции на снятие этого давления отражают функционирование, как минимум, двух подтипов (on– и off–) в каждой из групп тонических и фазных рецепторов. Комплексы рецепторов, противофазно реагирующих на механический стимул, давно уже известны в системе боковой линии рыб и земноводных [19, 20]. Такие аналогии в функциональном составе механо-рецепторных систем, по-видимому, отражают общие принципы обработки тактильной сенсорной информации у различных видов позвоночных.
Опыты показали, что наиболее интенсивные тактильные реакции во всех сенсорных зонах поверхности головы вызывали скользящие стимулы. По-видимому, такой стимул является наиболее адекватным, сигнализирующим о характере движения и ориентации рыбы в водной среде. Основной поток тактильных раздражений на поверхности головы рыб при плавании является прямым следствием, как внешних воздействий, так и движений самой рыбы. Эффективная обработка внешней информации возможна при выполнении двух условий: 1 – в сенсорный канал поступает информация о моторной программе движения животного, так называемой, моторной копии [21] и 2 – компенсаторная стабилизация положения динамического диапазона рецепторных элементов в соответствии с моторной активностью животного [22]. Выполнение обоих условий подразумевает активное влияние эфферентной регуляции как в центральных отделах сенсорного канала, так и непосредственно на рецепторном уровне. Именно таким влиянием можно объяснить различные формы модуляции тактильных ответов во времени и модификацию ответов в процессе повторных сеансов стимуляции одних и тех же точек на коже головы. Этим же обстоятельством, по-видимому, обусловлено временное увеличение амплитуды ответов на тактильное раздражение при действии лидокаина. Можно предположить, что лидокаин блокировал проведение спайковой активности по эфферентным нервным волокнам и тем самым способствовал снижению центрального тормозного влияния на тактильные рецепторы.
Теоретическое исследование процессов выделения внешних сигналов на фоне сенсорного потока, вызванного собственным движением животного, показало, что эффективность обнаружения слабых тактильных внешних сигналов в таких условиях существенно зависит от способа представления сенсорной информации на выходах рецепторного аппарата. В частности, повышению эффективности обнаружения источника сигнала способствует рецепция не только самого исходного сигнала, но и его производных во времени [21, 22]. Этим можно объяснить изобилие зарегистрированных нами фазных тактильных рецепторов, имевших различные временные характеристики ответов.
Распределение тактильной чувствительности на голове рыб имеет не равномерный, а зональный характер. Локализация сенсорных зон видоспецифична и связана с особенностями биологии вида [6, 7]. В наших опытах у карася и карпа наиболее высокая тактильная чувствительность зарегистрирована в кожных покровах головы околоротовой и вентральной локализации, а наиболее низкая – в зонах дорсальной и каудальной локализации. По-видимому, такое распределение имеет вполне определенную экологическую обусловленность. Объекты настоящего исследования являются факультативными бентофагами. Для обитателей придонных биотопов особенно важную роль при поиске пищевых объектов играют рецепторы как ростральной, так и вентральной локализации.
Зональная организация тактильной рецепции составляет основу формирования тактильной сенсорной системы, где каждая зона является элементом этой системы. Характер распределения и иннервации тактильно чувствительных сенсорных зон на голове рыб совпадает с зональностью распределения вкусовой рецепции [6, 23]. Это дает основание предполагать существование их взаимодействия и формирования обобщенной тактильно-вкусовой информации [24]. Взаимодействие сенсорных систем контактного и дистантного действия составляет основу полисенсорной ориентации и формирования сложных форм поведения животных и рыб. Представители сем. Cyprinidae, к которому относятся объекты настоящего исследования, отличаются особенно высоким уровнем полисенсорности поведенческих адаптаций [2, 3].
Таким образом, с помощью методики неинвазивной регистрации кожных реакций в ответ на тактильную стимуляцию было показано, что рыбы, так же как и наземные позвоночные, обладают тактильными рецепторами фазного и тонического типов. Большую функциональную значимость тактильной рецепции обеспечивают ее физиологические особенности: высокая чувствительность и низкая латентность ответных реакций, а также ее взаимодействие с другими сенсорными системами – зрительной и вкусовой [9–11, 15].
Характер распределения тактильной чувствительности на голове исследованных видов рыб связан с особенностями их пищевого поведения. Повышенная плотность распределения тактильных рецепторов в сенсорных зонах головы карповых рыб создает оптимальные условия для ориентации в пространстве, а также при поиске и распознавании пищевых объектов.
Список литературы
Протасов В.Р. Поведение рыб. М.: Пищевая промышленность. 296 с. 1978. [Protasov V. R. Fish behavior. M.: Food industry. 296 p. 1978. (in Russ)].
Павлов Д.С., Касумян А.О. Сенсорные основы пищевого поведения рыб. Вопр. ихтиологии. 30 (5): 720–732. 1990. [Pavlov D.S., Kasumyan A.O. Sensory base of food behavior in fish. J. Ichthyology. 30 (5): 720–732. 1990].
Суворов Е.К. Основы ихтиологии Л.: Советская наука. 579 с. 1948. [Suvorov E.K. Base of Ichthyology. L.: Sovetskaya nauka. 579 p. 1948. (in Russ)].
Tan D., Patton P., Coombs S. Do blind cave fish have behavioral specializations for active flow-sensing. J. Comp. Physiol. 197A: 743–754. DOI: 011 0637 6.https://doi.org/10.1007/ s00359
Ильинский О.Б. Вопросы физиологии сенсорных систем. Механорецепторы. Л.: Наука. 82 с. 1967. [Ilinsky O.B. Problems of physiology in sensory systems. Mechanoreceptors. L.: Nauka. 82 p. 1967 (in Russ)].
Kasumyan A.O. Tactile reception and behavior of fish. J. Ichthyol. 2011. 51 (11): 1035–1103. https://doi.org/10.1134/S003294521111004X
Смит К. Биология сенсорных систем. М.: БИНОМ. 583 с. 2005. [Smith C. Biology of Sensory systems. John Willey and Sons. LTD. 515 p. 2005].
Павлов Д.С. Биологические основы управления поведением рыб в потоке воды. М.: Наука. 319 с. 1979. [Pavlov D.S. The basis of the control of fish Behavior in a water flow. M.: Nauka. 319 p. 1979. (in Russ)].
Marui T., Caprio J., Kiyohara S., Kasahara Y. Topographical organization of taste and tactile neurons in the facial lobe of the Sea Catfish, Plotosus lineatus. Brain Res. 446: 178–182. 1988. https://doi.org/10.1016/0006-8993(88)91312-1
Belousova T.A., Devitsina G.V., Malyukina G.A. Functional peculiarities of fish trigeminal system. Chemical senses. 8 (2): 121–130. 1983.
Pusdrowsky R.L. Afferent projection of the trigeminal nerve in the goldfish, Carrasius auratus. J. Morphol. 198: 131–147. 1988.
Chervova L.S., Lapshin D.N. The Effect of olfactory deprivation on the thresholds of sensitivity of skin chemoreceptors of carp Cyprinus carpio. J. Ichthyology. 50 (11): 1030–1035. 2010. https://doi.org/10.1134/S0032945210110081
Девицина Г.В., Лапшин Д.Н. Тактильная рецепция наружных покровов головы серебряного карася (Carassius auratus gibelio). ДАН. 466 (4): 493–495. 2016. [Devitsina G. V., Lapshin D. N. Tactile reception in the outer head cover of goldfish (Carassius auratus gibelio). Doklady Biological Sciences. 466: 21–23. 2016]. https://doi.org/10.1134/S0012496616010075https://doi.org/10.7868/S0869565216040253
Лекарственные препараты в России: Справочник Видаль. М: ВидальРус. 1480 с. 2015. [Medicinal preparations in Russia. Formulary. M: WidalRus. 1480 p. 2015. (in Russ)].
Девицина Г.В., Головкина Т.В. Структурная организация вкусового аппарата у харациновых рыб (Characidae, Teleostei). Вопросы ихтиологии. 58 (1): 53–66. 2018. Devitsina G.V., Golovkina T.V. Structural organization of the Taste Apparatus in Characins (Characidae, Teleostei). J. Ichthyology. 58 (1): 59–72. 2018. https://doi.org/10.1134/S0032945218010046
Appelbaum S., Riehl R. Scanning electron microscopic observation of the chemo- and mechanoreceptors of carp larvae (Cyprinus carpio) and their relationship to early behaviour. Aquat. Living Resour. 10: 1–12. 1997. https://doi.org/10.1051/alr:1997001
Whitear M. The Free Nerve Endings in Fish Epidermis. J. Zool. London. 163: 231–236. 1971.
Harris G.G., Frishkopf L.S., Flock A. Receptor potentials from hair cells of the lateral line. Science. V: 76–79. 1970.
Krouse A.B.A., van der Zalm J.M., van den Bercken J. Extracellular receptor potentials from the lateral-line organ of Xenopus laevis. J. Experimental biology. 86: 63–77. 1980.
Poulet J.F.A., Hedwig B. The cellular basis of a corollary discharge. Science. 27(311): 518–522. 2006. https://doi.org/10.1126/science.1120847
Poulet J.F.A., Hedwig B. New insights into corollary discharges mediated by identified neural Pathways. Trends in Neuroscience. 30 (1): 14–21. 2007. https://doi.org/10.1016/j.tins.2006.11.005
Лапшин Д.Н., Лукьяница А.А. Нейросетевое моделирование процедуры обнаружения внешних сигналов на фоне сенсорного потока, вызванного собственным движением животного. Сенсорные системы. 26 (1): 69–75. 2012. [Lapshin D.N., Lukyanitsa A.A. Neural network model of signal extraction from a complex sensory flow containing self-generated noise. Sensornye systemy. 26 (1): 69–75. 2012. (in Russ)].
Девицина Г.В. Адаптивная изменчивость рецепторного отдела вкусовой системы карпа (Cyprinus carpio L.) после хронической аносмии. Журн. эвол. биохим. и физиол. 42 (6): 589–594. 2006. [Devitsina G.V. Adaptive variability of the gustatory system receptor part of the carp, Cyprinus carpio (Cyprinidae, Teleostei) after chronic Anosmia. J. Evol. Biochem. and Physiol. 42 (6): 743–750. 2006]. https://doi.org/10.1134/S0022093006060111
Herrick C.J. On the centers for taste and touch in the medulla oblongata of fishes. J. Comp. Neurol. 16: 403–439. 1906.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии