

Журнал эволюционной биохимии и физиологии, 2021, T. 57, № 3, стр. 240-249
ПРОДРОМАЛЬНЫЕ ХАРАКТЕРИСТИКИ ЭПИЛЕПСИИ У КРЫС С МАЯТНИКООБРАЗНЫМИ ДВИЖЕНИЯМИ
Т. А. Алехина 1, В. С. Плеканчук 1, 2, *, Л. В. Осадчук 1
1 Федеральный исследовательский центр Институт цитологии и генетики Сибирского отделения
Российской академии наук
Новосибирск, Россия
2 Новосибирский государственный университет
Новосибирск, Россия
* E-mail: lada9604@mail.ru
Поступила в редакцию 10.12.2020
После доработки 14.01.2021
Принята к публикации 22.02.2021
Аннотация
На селекционной линии крыс с маятникообразными движениями (МД) разрабатывается модель фокальных приступов с автоматизмами. 90% животных линии МД в дефинитивном возрасте дают припадки на аудиогенный раздражитель. В отличие от линий Крушинского–Молодкиной и GEPR-9 (“genetically epilepsy-prone rats”), в популяции которых наблюдаются генерализованные судороги, особи с МД демонстрируют клонический каскад приступов – стереотипные прыжки, достигающие в высоту 0.5 м и со скоростью 1 прыжок в секунду. Линия МД является новой моделью эпилептиформных реакций и практически не исследовалась в раннем онтогенезе. Для обнаружения продромальных признаков эпилепсии поставлена цель исследовать поведенческие и гормонально-метаболические параметры на модели МД в неонатальном периоде развития. Контролем служили крысы линии Вистар, которая являлась исходной популяцией при селекции. В результате проведенного исследования к продромальным характеристикам линии МД мы отнесли такие параметры, как замедление роста локомоторной активности и увеличения массы тела и семенников, затормаживание подъема уровня холестерина в плазме крови на 10-й и 14-й день развития. Наряду с этим показан сдвиг пика циркулярных движений у крыс МД. Выявлено повышенное проявление возбудимых реакций – вокализаций и пароксизмов. Установлена фазное изменение содержания триглицеридов в крови. Данные корреляционного анализа указывают, что у самцов линии Вистар (контрольная линия) уровень тестостерона в плазме крови достоверно отрицательно взаимосвязан с массой тела и семенников, и положительно – с уровнем три-глицеридов в крови. В то же время такие же корреляции у крыс МД были недостоверными. Эти факты говорят о дестабилизации развития морфофизиологических признаков у крыс линии МД по сравнению с контрольной линией крыс Вистар. Учитывая выявленные продромальные признаки в ходе усиления маятникообразного гиперкинеза (замедление темпов развития, сдвиг фаз циркулярного движения, содержания триглицеридов и изменение взаимосвязей коррелированных фенотипических признаков), линию крыс МД можно рассматривать как модель для дальнейшего изучения биологической основы эпилептиформной патологии.
ВВЕДЕНИЕ
В исследованиях на популяциях человека выявлены признаки “повышенной готовности” нервной системы к эпилептическим расстройствам у детей младенческого возраста. К таким признакам относят моторные реакции с клоническими, тоническими и миоклоническими приступами, диспластические процессы, замедленное развитие двигательных реакций, задержка набора в весе тела и органов [1–3]; в качестве коморбидных проявлений рассматривается дисбаланс метаболического обмена [4–6]. Все эти отклонения указывают на ранние сдвиги в нейрогормональных и метаболических процессах в организме и могут рассматриваться как маркеры генетически обусловленных эпилептических расстройств в раннем развитии.
Фокальные моторные приступы с типичными автоматизмами являются одной из форм эпилепсии. Для изучения механизмов данного синдрома и взаимосвязи с функционированием физиологических систем используются модели лабораторных животных. В.Г. Колпаковым были созданы две модели кататонического типа реагирования – линия ГК (Генетическая Кататония) [7] и линия с маятникообразными движениями (МД) [7–10]. По общей фенотипической картине две эти линии отличаются повышенным стрессорным ответом на внешние воздействия и ассоциированной с этим признаком пониженной рождаемостью потомства. Известное антагонистическое взаимодействие между надпочечниками и гонадами в ходе селекции приводит к тому, что стресс подавляет половую активность [11]. Механизмы такого явления в раннем онтогенезе на линии МД не изучены.
Для животных с МД характерны ритмичные раскачивания головы и передней части тела при отсутствии локомоций (рис. 1).
Маятникообразные движения наблюдаются не только у мышей и крыс, но и у кроликов-альбиносов и скорпионовых хомячков [цит. по 10, 12]. У крыс, селекционированных на увеличение амплитуды маятникообразных движений (МД), после 40 поколения отбора стали развиваться аудиогенные припадки в 80–90% случаев в отличие от исходной (контрольной) популяции Вистар, в которой эпилептические приступы наблюдаются только у 25–30% животных [13]. На сегодняшний день по нашим данным отмечено 10% припадков у неселекционированных крыс Вистар и WAG (Wistar Albino Glaxo) с МД при действии аудиогенного раздражителя. Факты по изменению предрасположенности к аудиогенным припадкам в разных поколениях селекции указывают на важную роль наследственной изменчивости при отборе по поведению [11]. В линии МД произошел сдвиг от кататонического типа реагирования до эпилептиформного. Соответственно произошли и отклонения в уровнях нейротрансмиттеров в мозге [13, 14]. В линии МД эти отклонения оказались сходными с таковыми у крыс Крушинского–Молодкиной и GEPR-9 (“genetically epilepsy-prone rats”) [13]. В настоящее время в пользу того, что в качестве модели фокальных моторных приступов с типичными автоматизмами можно рассматривать линейных крыс с МД, говорят следующие факты. У животных этой линии наблюдаются 50% абортивных аудиогенных припадков [13, 15], снижение уровня таурина в гиппокампе [14], найдена положительная корреляция между интенсивностью припадков и длительностью постиктальной каталепсии в отличие от контрольной линии Вистар [15]. Показано также, что в ходе отбора по усилению МД снизился вес тела у половозрелых особей, а также уровень фертильности и доля пропустовавших самок [15].
Перинатальный период онтогенеза является критическим для формирования нервной системы и развития постурально-моторных реакций. В классических работах уделяется особое внимание становлению двигательного поведения как пространственно-временного процесса [16–18]. В двигательном поведении исследователи выделяют составляющие его компоненты, анализируют их появление, частоту и выраженность. К ним относят движение головы, повороты туловища, локомоции, стойки, груминг. Основываясь на подробном анализе двигательного поведения, были выделены 4 критические точки в неонатальном развитии. Это 1-й день – движения головы и ползание, 7-й день – созревание кортикоспинального тракта, 10-й день – становление на 4 лапах и хождение, 14-й день – открытие ушей и глаз и формирование кругозора [16–18].
В неонатальный период происходит развитие репродуктивной системы организма. Выяснение физиологических механизмов преобразования репродуктивной функции в результате поведенческой селекции является ключевой проблемой, поскольку от успешного воспроизведения здорового потомства зависят существование вида и его эволюция. Сравнительный анализ морфологических и гормональных признаков развития семенников в раннем развитии позволяет установить отклонения у крыс линии МД по сравнению с контрольной линией Вистар, что может выявить предикторные особенности снижения фертильности у крыс экспериментальной линии. Тестостерон, который синтезируется в клетках Лейдига семенников, влияет на развитие и накопление костно-мышечной массы [19], регуляцию сперматогенеза и полового поведения [20] и взаимодействует с такими липидами, как холестерин и триглицериды [21, 22]. Липиды играют полифункциональную роль в онтогенезе: являются энергетическим резервом для развивающегося организма, служат предшественниками биологически активных веществ, стабилизируют работу мембран клеток, являются предшественниками в синтезе кортикостероидных и половых гормонов.
Селекционируемая линия крыс МД с частотой 95% маятникообразного гиперкинеза является новой линией. Животные этой линии в раннем онтогенезе практически не исследовались. Целью настоящей работы стали изучение развития моторных реакций, характеристика морфологической и гормональной функции гонадной системы и липидного обмена в критических точках раннего онтогенеза у крыс линии с МД. Дополнительно была поставлена задача – провести сравнительный анализ полученных данных с таковыми у крыс линии ГК, представляющих модель кататонического типа реагирования. Такое сравнение позволит определить механизмы возникновения эпилептиформных реакций у крыс МД и выявить предикторные свойства эпилепсии у крыс-маятников в дефинитивном возрасте.
МАТЕРИАЛЫ И МЕТОДИКА
Экспериментальные животные: крысы линий МД (n = 116) и Вистар (n = 126), всего n = 242. Животные содержались в стандартных условиях вивария ИЦиГ в свободном доступе к воде и корму. Все экспериментальные процедуры выполнены с соблюдением всех норм, изложенных в директивах Европейского сообщества (86/609/ЕЕС) и Хельсинкской декларации по защите позвоночных животных, используемых для лабораторных целей.
Последовательность опытов:
I часть: тесты на активность моторных подсистем и щипковую каталепсию.

II часть: взятие крови, взвешивание тела и семенников, биохимический анализ.
Для получения потомства нужного возраста проводили спаривание самцов и самок в линиях МД и Вистар. На последней неделе беременности самок отсаживали в индивидуальные клетки и день родов обозначали как первый день жизни крысят. Регистрировали количество детенышей в каждом помете при рождении. Непосредственно перед декапитацией самцов взвешивали, после декапитации собирали периферическую кровь, выделяли семенники и взвешивали.
Тест на активность моторных подсистем [16–18]
Тестирование проводилось на открытой площадке размером 20 × 30 см с подогревом до +30–35°С; температура поддерживалась на уровне температуры гнезда с крысятами. Поведение снималось на видео камеру Panasonic в течение 1 мин. Крысенка брали за кожу дорсальной части шеи, помещали на середину площадки хвостом к экспериментатору. Регистрировали следующие параметры: число двигательных реакций головы, циркулярных движений, “плавательных” движений, локомоций, время иммобильности (сек). Каждый детеныш тестировался не более 3 раз из 14 дней.
Тест на щипковую каталепсию [23]
Щипковая каталепсия тестировалась у 2-недельных крысят путем приподнимания за кожу дорсальной поверхности шеи. Регистрировались следующие параметры: время иммобильности (сек), число пароксизмов, время вокализации и время с закрытыми глазами (сек). Поведение снималось на видео камеру Panasonic в течение 2 мин.
Определение тестостерона в сыворотке крови и гомогенатах семенников. Использовали методику, описанную ранее [24, 25]. Коротко: периферическую кровь центрифугировали 20 мин при 3000 об/мин и при +4°С, сыворотку хранили при –40°С, оба семенника гомогенизировали в 500 мкл фосфатного буфера, центрифугировали и супернатант хранили при –40°С. Тестостерон в сыворотке крови и гомогенатах семенников определяли иммуноферментным методом с использованием наборов “Стероид ИФА-тестостерон-01” (Алкор Био, Санкт-Петербург) по прилагаемой инструкции. Диапазон определения концентраций 0.2–50 нмоль/л. Чувствительность 0.2 нмоль/л. Концентрацию тестостерона в сыворотке выражали в нмоль/л.
Определение показателей липидного обмена. Использовали методику, описанную ранее [25]. Уровень общего холестерина (ОХ) и триглицеридов (ТГ) определяли ферментативным колориметрическим методом на планшетной модификации стандартными наборами Вектор Бест (Новосибирск) по прилагаемой инструкции. Диапазон определения концентраций 0.2–50 ммоль/л. Чувствительность 0.2 ммоль/л.
Статистическая обработка данных. Полученные данные обрабатывались с использованием пакета программ “STATISTICA 10.0”. Данные в таблицах и рисунках представлены как средняя арифметическая и ошибка среднего (mean ± SEM). При сравнении линий МД и Вистар применяли 2-факторный анализ ANOVA (фактор генотипа и фактор возраста). По отдельным точкам в онтогенезе (1-, 7-, 10-, 14-й день) сравнивали по post hoc LSD-критерию. В качестве статистически достоверного уровня значимости считался p < 0.05. Использовали программу Correlation Matrices для подсчета коэффициентов корреляции (R) между показателями массы тела, массы семенников, уровней тестостерона, холестерина и триглицеридов.
РЕЗУЛЬТАТЫ
Зарегистрирована сниженная рождаемость в линии МД (30 пометов) по сравнению с линией Вистар (20 пометов): 7.0 ± 0.5 и 10.6 ± 0.9 крысят в помете (p < 0.001) соответственно.
Поведенческие тесты. Крысы Вистар и МД различаются по тесту на активность моторных подсистем. Основные параметры этого теста – движения головы, циркулярные, “плавательные” движения, локомоции и застывание (табл. 1; рис. 2, рис. 3). “Плавательными” движениями считали такие движения, когда крысята, не отрывая вентральной поверхности своего тела от пола площадки, “загребали” лапками, что и напоминало “плавательные” движения. Особое внимание было уделено 1-му дню жизни особей. Крысята рождаются с ограниченными возможностями принятия сенсорной информации (уши и глаза у них закрыты), и поэтому влияние фактора генотипа прослеживается наиболее четко из-за отсутствия сформированных зрительных и слуховых рецепторов при восприятии внешних стимулов. Результаты тестирования показаны в табл. 1.
Таблица 1.
Двигательные реакции у крысят Вистар и МД в первый день жизни
| Параметры | Вистар (24) | МД (18) |
|---|---|---|
| Движения головы (n) | 2.92 ± 0.45 | 1.00 ± 0.28** |
| Циркулярные движения (n) | 1.56 ± 0.24 | 1.21 ± 0.20 |
| “Плавательные” движения (n) | 0.88 ± 0.22 | 0.16 ± 0.12** |
| Время иммобильности (сек) | 41.8 ± 5.3 | 52.4 ± 2.0# |
| Общая активность (n) | 5.00 ± 0.64 | 1.72 ± 0.29*** |
Рис. 2.
Циркулярные движения у крыс Вистар и МД. Различия достоверны по влиянию фактора генотипа: F(5, 256) = 4.62; p < 0.001.
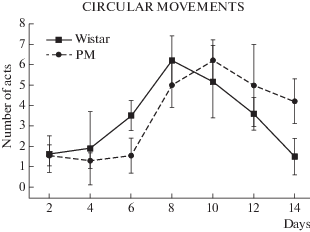
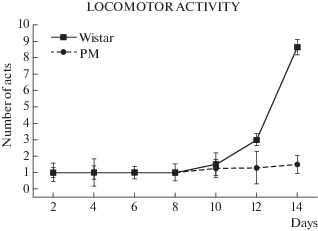
Рис. 3.
Локомоторные акты у крыс Вистар и МД. Различия достоверны по влиянию фактора генотипа: F(5, 237) = 25.45; p < 0.001.
На число актов циркулярного движения в линиях Вистар и МД оказал фактор генотипа (F(5,256) = = 4.62; p < 0.001). Крысята отличались в течение 2 нед тестирования по динамике циркулярного движения: пик у Вистар пришелся на 8-й день, пик у МД – на 10-й день.
На параметр локомоторной активности повлиял фактор генотипа (F(5, 237) = 25.45; p < 0.001 у двух линий. Это влияние выразилось в отставании темпов роста локомоций у крысят группы МД по сравнению с крысятами группы Вистар на 12-й и 14-й день.
Тест на щипковую каталепсию в 2- недельном возрасте [23].
Данные по тесту щипковая каталепсия различаются в группах Вистар и МД (табл. 1). Показатели делятся на “позитивные” и “негативные” признаки. К “позитивным” признакам, отражающим возбудимость нервной системы, относятся повышенное число пароксизмов и время вокализации. Крысы МД превосходят крыс Вистар по этим параметрам. Показатель “время с закрытыми глазами” отражает трансподобное (полусонное) состояние, характерное для каталептических реакций [26]. В общей сложности, по негативным признакам – времени иммобильности (табл. 1, табл. 2) и по длительности периода с закрытыми глазами, животные МД показывают значимые различия по сравнению с исходной линией Вистар. В целом кататонические черты – “позитивные” и “негативные”, прослеживаются в исследуемой линии МД.
Таблица 2.
Тест на щипковую каталепсию в 2-недельном возрасте у крыс Вистар и МД
| Показатели | Вистар (35) | МД (33) |
|---|---|---|
| Время иммобильности (сек) | 63.6 ± 2.4 | 72.8 ± 3.2* |
| Число пароксизмов (n) | 0.5 ± 0.1 | 1.9 ± 0.3** |
| Время вокализации (сек) | 2.0 ± 0.5 | 7.8 ± 1.3*** |
| Время с закрытыми глазами (сек) | 29.7 ± 2.7 | 47.9 ± 5.3** |
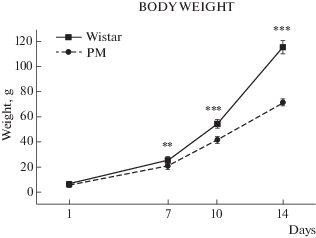
Морфологические параметры. Межлинейные показатели “массы тела” оказались под влиянием фактора генотипа (F(1, 58) = 5.0; p < 0.03) и возраста (F(3, 58) = = 104.1; p = 0.0000) (см. рис. 4). В итоге набор массы тела у крыс МД прогрессивно уменьшался в критических точках развития – на 7-й, 10-й и 14-й день.
Рис. 4.
Масса тела у крыс Вистар и МД. Различия достоверны по фактору генотипа F(1,58) = 5.0; p < 0.03 и возрасту (F(3, 58) = 104.1; p < 0.000).
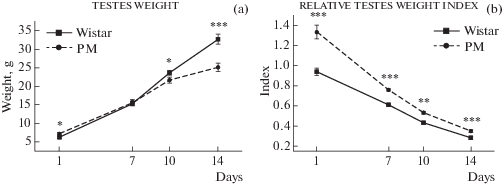
Сходным образом изменялась масса семенников. Отмечено влияние генотипа (F(1, 58) = 60.3; p < < 0.000) и возраста (F(3, 58) = 385.9; p < 0.000). На рис. 5a видно, что масса семенников достоверно отстает у крыс МД по сравнению с Вистар, начиная с 10-й дня и разница между массой семенников у крыс разных линий увеличивается к 14-й дню. Однако отношение массы семенников к массе тела было достоверно выше у крыс МД по сравнению с крысами Вистар во всех четырех точках (рис. 5b). Эти данные указывают на наличие диспластических черт и на эффект дестабилизации в ходе отбора на усиление МД у селекционируемой линии.
Рис. 5.
Масса семенников (a) и индекс относительной массы семенников (b) у крыс Вистар и МД. Различия достоверны по влиянию факторов генотипа (F(1, 58) = 60.3; p < 0.000) и возраста (F(3, 58) = 385.9; p < 0.000). Различия достоверны на уровне значимости * p < 0.05, ** p < 0.01, *** p < 0.001 по LSD-критерию.
Гормон-липидные параметры.
Показано влияние на уровень холестерина факторов генотипа (F(1, 58) = 7.37; p < 0.01) и возраста (F(3, 58) = 54.3; p < 0.000) (рис. 6a). На уровень три-глицеридов в крови не было обнаружено эффекта вышеуказанных факторов, однако при использовании дисперсионного анализа установлено значимое влияние взаимодействия факторов генотипа (F(3, 58) = 6.2; p < 0.001) и возраста (F(3, 58) = 54.3; p < < 0.000) в зависимости от критической точки неонатального периода (рис. 6b).
Рис. 6.
Уровень холестерина (a) и триглицеридов в плазме крови (b) у крыс МД и Вистар. Различия достоверны по влиянию фактора генотипа (a): F(1, 58) = 7.37; p < 0.01 и по взаимосвязи факторов генотипа и возраста (b): F(3, 58) = 6.2; p < 0.001. * p < 0.05, ** p < 0.01, *** p < 0.001 отмечены различия на уровне значимости и # p = 0.07 – тенденция по LSD-критерию.
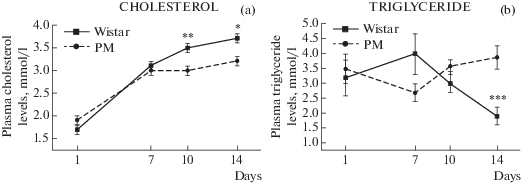
Рис. 7.
Уровень тестостерона в плазме крови (a) и тестостерона в семенниках (b) у крыс МД и Вистар. Различия недостоверны по влиянию фактора генотипа (a): F(1, 58) = 1.69; p < 0.02) и (b): F(1, 58) = 1.0; p < 0.02).
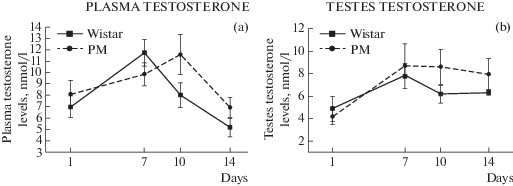
По уровню тестостерона в плазме крови и в семенниках не было найдено различий между двумя линиями: F(1, 58) = 1.6; p > 0.05 и F(1, 58) = 1.0; p > 0.05 соответственно.
Корреляционный анализ. В корреляционный анализ были включены следующие показатели: масса тела, масса семенников, уровень тестостерона в крови и в семенниках, уровни холестерина и три-глицеридов. Коэффициент корреляции обозначен как R. Самые высокие значения у двух линий R = = 0.72 и R = 0.64 установлены между уровнем холестерина и массой тела и семенников соответственно. Наибольшее количество достоверных корреляций выявлено по взаимосвязи уровня тестостерона в плазме с вышеобозначенными показателями у крыс Вистар, но не для содержания этого гормона в семенниках. К таким показателям, показавшим достоверный R с уровнем тестостерона в плазме крови, были отнесены положительные корреляции у особей Вистар: Rтестостерон в семенниках = 0.44; Rтриглицериды = 0.56 и отрицательные: Rмасса тела = = –0.38; Rмасса семенников = –0.48. Для линии МД коэффициенты корреляций между уровнем тестостерона в плазме крови с вышеобозначенными показателями имели низкие и недостоверные значения: Rхолестерин = 0.11; Rтриглицериды = 0.01; Rмасса тела = –0.06; Rмасса семенников = –0.21. Последние факты указывают на эффект дестабилизации организменных систем, к которому привела селекция на усиление гиперкинеза в линии МД.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В настоящей работе было прослежено развитие ряда фенотипических характеристик в неонатальном периоде у крыс линии МД, которые стали предикторами эпилептиформных реакций у животных в дефинитивном возрасте. Эти реакции были разделены на “отрицательные” и “положительные” проявления. К “отрицательным” признакам были отнесены те, которые развивались с задержкой в онтогенезе или проявлялись в заторможенном состоянии. Эти параметры отражают темпы наследственной изменчивости и, в итоге, могут привести к появлению патологических признаков в линии МД. К таким признакам мы отнесли замедленное развитие локомоторных реакций, повышенную иммобильность, более длительный период времени с закрытыми глазами, сдвиг в циркулярных движениях и отставание в наборе веса тела, более медленный подъем холестерина у крыс МД по сравнению с контролем. “Положительными” считали те признаки, которые имели свойство повышенной возбудимости. Это – большее число стереотипных пароксизмов и более длительный период вокализации. Обе категории признаков – и “отрицательные”, и “положительные”, наблюдаются в значительной степени у крыс линии МД и демонстрируют кататонические черты в селекционируемой линии.
Нам важно было сравнить фенотипические признаки у линий МД и ГК, поставленные в качестве дополнительной задачи в этой работе. На ранних этапах селекции реакции животных с МД исследователи относили их к кататоническому типу реагирования, поскольку этим особям было свойственно проявление маятникообразного гиперкинеза при отсутствии локомоций [27–29]. Однако после 40-го поколения отбора у крыс с МД резко выросла доля аудиогенных припадков (до 83%) [13]. Авторы селекции линии МД стали относить реакции животных к эпилептиформным. Если на первых этапах селекции линии МД была показана положительная корреляция между МД и спонтанной каталепсией, то на последних стадиях она оказалась отрицательной в форме перевернутого “U” [9]. В настоящее время у крыс линии ГК (“Генетическая” и “Кататония”) не наблюдаются МД, тогда как у крыс линии МД обнаруживаются аудиогенные припадки в 90% случаев.
Результаты сравнительного фенотипического анализа линий МД и ГК в дефинитивном возрасте, селекционированные из одной и той же популяции Вистар и содержащиеся в одинаковых условиях вивария ИЦиГ (Новосибирск), показали следующее. Обе эти модели являются предположительно моделью кататонического типа реагирования [9, 27–30]. Между моделью кататонии (линия ГК) и моделью эпилепсии (линия МД) наблюдаются разнонаправленные отклонения по фенотипическим признакам. В список противоположно направленных параметров вошли аудиогенные припадки [15, 31], количество стереотипий [9, 32, 33], рефлекс вздрагивания [9, 15, 31], уровень артериального давления [15, 33], спонтанная каталепсия [7, 15, 31]. Были найдены и сходные характеристики между животными линий ГК и МД. Они проявляются в реакциях повышенной возбудимости в разных тестовых ситуациях. Это – эпилептиформные реакции по результатам ЭЭГ у крыс ГК [34], реакций агрессии по отношению к человеку [29, 35], импульсивность, а также сниженная фертильность [36, 37], более низкий вес тела [15, 31]. Все эти вышеуказанные характеристики – и антагонистические, и сходные с ними, наблюдаются у людей-эпилептиков [38–40].
В настоящей работе изучались фенотипические характеристики в неонатальном периоде у крыс с МД, и по общей картине исследуемых признаков был выявлен ряд сходных черт с линией ГК. У крыс двух линий в этот период обнаружено повышение времени иммобильности, снижение двигательных реакций и пониженный уровень рождаемости [15, 31]. Отличительной особенностью поведения на модели ГК в раннем онтогенезе выделены такие предикторы кататонического поведения, как дискинетические движения и асимметричные позы [41], чего не было показано на модели с МД.
Если мы обратимся к сдвигам в гормонально-метаболической системе, то увидим, что эндофенотипы линий ГК и МД формировались по разным направлениям. Общий показатель холестерина в плазме крови показал разнонаправленные отклонения у крыс ГК [36] и МД (показанные в этой статье) по сравнению с контролем.
В неонатальном периоде протекает половая дифференцировка организма по мужскому типу, формируются нейросекреторные гипоталамические центры, прямые и обратные связи в системе гипоталамус-гипофиз-семенники [42, 43]. У крыс при нормальном протекании к концу этого периода наблюдается снижение уровня половых стероидных гормонов (в том числе, и тестостерона) в крови, обусловленное снижением их секреции клетками Лейдига семенников и замещением эмбрионального типа клеток на дефинитивный. Общий характер динамики тестостерона в крови оказался сниженным по окончании неонатального периода у крыс Вистар и МД. Ранее было установлено смещение пика тестостерона на более поздние сроки у самцов ГК по сравнению с самцами Вистар в раннем возрасте [36], тогда как у особей линии МД не было обнаружено различий в уровнях тестостерона ни в плазме крови, ни в клетках Лейдига семенников. Изменение уровня и продукции тестостерона в развивающемся мужском организме приводит к последующим изменениям в метаболизме, уменьшению мышечной и физической силы и снижению половой функции у взрослых особей [44–46].
Тем не менее у крыс линии МД по результатам корреляционного анализа в неонатальный период развития был выявлен дестабилизирующий эффект тестостерона. Уровень этого андрогена в крови у животных МД не связан коррелятивными отношениями с такими параметрами, как уровни холестерина, триглицеридов и весом семенников. В этом же опыте были установлены достоверные уровни корреляций у крыс Вистар по всем вышеперечисленным параметрам, которые указывали на интегрированность эндокринной системы у животных исходной линии. Факт дестабилизации морфофизиологических параметров подтвержден и у взрослых крыс линии МД по сравнению с животными из популяции Вистар [15].
В период написания статьи для выявления аудиогенных припадков были протестированы неселекционированные крысы Вистар и WAG (Wistar Albino Glaxo) со стереотипным качанием головой, и мы получили в этих группах в общей сложности 10% аудиогенных приступов. В группе неселекционируемых грызунов амплитуда раскачивания головы и плечевого пояса составляла 0.5 см в одну и в другую стороны. В то же время у крыс селекционированной группы МД амплитуда маятникообразных движений достигает до 4 см в обе стороны. В более ранних работах показано возрастание частоты приступов в ходе селекции на усиление МД в линии: от 41% (в 5-м поколении) до 91% (в 23-м поколении) [9, 13, 30, 31]. Таким образом, к 50-му поколению селекции у крыс МД сформировался новый эндофенотип с измененной гормонально-метаболической основой. К его продромальным характеристикам мы отнесли такие параметры, как замедление роста локомоций и набора массы тела и массы семенников, замедленный пик появления циркулярных движений, проявление возбудимых реакций (вокализаций и пароксизмов), снижение уровня холестерина, нарушение корреляционных связей между уровнем тестостерона в крови и морфо-липидными параметрами. Отбор животных по увеличению амплитуды МД от кататонических признаков (иммобильности и стереотипий) привел к усилению эпилептиформных реакций у взрослых особей. Выявленные продромальные отклонения в неонатальный период являются неспецифическими и представляют собой основу для появления маятникообразных движений в препубертатном и дефинитивном периодах развития. В перспективе предполагается повлиять на эти признаки, в том числе и фармакологическими методами, приводящими к снижению риска возникновения эпилептиформной патологии.
Список литературы
Sculiera C., Gasparda N. (2019) New onset refractory status epilepticus (NORSE). Seizure: European Journal of Epilepsy 68: 72–78. https://doi.org/10.1111/epi.14022
Gataullina S., Dulac O., Bulteau C. (2015) Temporal lobe epilepsy in infants and children. Rev neurologique 171: 252–253. https://doi.org/10.1016/j.neurol.2015.01.559
Camfield P., Camfield C. (2019) Regression in children with epilepsy. Neurosci Biobehav Rev 96:210–218. https://doi.org/10.1016/j.neubiorev.2018.12.008
Park J.T., Shahid A.M., Jamoul A. (2015) Common Pediatric Epilepsy Syndromes. Pediatric Annals 44 (2): e30–e35. https://doi.org/10.3928/00904481-20150203-09
Harrison V.S., Oatman O., Kerrigan J.F. (2017) Hypothalamic hamartoma with epilepsy: Review of endocrine comorbidity. Epilepsia 58 (2): 50–59. https://doi.org/10.1111/epi.13756
Kaprara A., Huhtaniemi I.T. (2018) The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 86:3–17. https://doi.org/10.1016/j.metabol.2017.11.018
Колпаков В.Г., Рицнер М.С., Корнетов Н.А., Самохвалов В.П., Залевский Г.В., Короленко Ц.П. (1985) Генетические и эволюционные проблемы психиатрии. Ред. А.С. Тимофеева. Наука: 256. [Kolpakov V.G., Richner M.S., Kornetov N.A., Samoshvalov V.P., Zalevskii G.V., Korolenko C.P. (1985) Genetic and evolutionary problems of psychiatry. Ed. Timofeeva. Nauka 256. (in Russ)].
Колпаков В.Г., Алехина Т.А., Барыкина Н.Н., Чугуй В.Ф., Попова Н.К. (2000) Некоторые физиологические проявления действия гена, контролирующего предрасположенность к маятникообразным движениям у грызунов. Рос. физиол. журн. им. И.М. Сеченова 86 (1): 33–40. [Kolpakov V.G., Alekhina T.A., Barykina N.N., Chuguĭ V.F., Popova N.K. (2000) Physiological manifestations of action of a gene regulating predisposition to a pendulum-like movements in rodents. Russ J Physiol 86(1):33–40. (in Russ)].
Колпаков В.Г., Барыкина Н.Н., Алехина Т.А., Чугуй В.Ф., Попова Н.К., Зоркольцева И.В., Аксенович Т.И. (1999) О генетических взаимоотношениях каталепсии и маятникообразных движений у крыс. Генетика. 35 (8): 1118–1123. [Kolpakov V.G., Barykina N.N., Alekhina T.A., Chuguĭ V.F., Popova N.K., Zorkol’steva I.V., Aksenovich T.I. (1999) Genetika. Genetic relationships between catalepsy and pendulum movements in rats. 35 (8): 1118–1123. (in Russ)].
Kolpakov V.G., Borodin P.M., Barykina N.N. (1977) Catatonic behaviour in the Norway rat. Behaviour 62: 190–208.
Беляев Д.К., Бородин П.М. (1982) Влияние стресса на наследственную изменчивость и его роль в эволюции. Эволюц. генетика Л 35–59. [Belyaev D.K., Borodin P.M. (1982) The impact of stress on the genetic variability and its role in evolution. Evolutionary genetics. L. 35–59. (in Russ)].
Колпаков В.Г., Барыкина Н.Н., Бородин П.М., Серова Л.И. (1976) Jactatio capitis – двигательная стереопития у грызунов. Генетика 12 (1): 71–77 [Kolpakov V.G., Barykina N.N., Borodin P.M., Serova L.I. (1976) Jactatio capitis – movement stereotypy in rodents. Genetika 12 (1): 71–77. (in Russ)].
Алехина Т.А., Прокудина О.И., Рязанова М.А., Уколова Т.Н., Барыкина Н.Н., Колпаков В.Г. (2007) Проявление типологических свойств поведения у линий крыс, селекционированных на усиление и отсутствие маятникообразных движений. Связь с моноаминами мозга. Журн. высш. нерв. деят. 57 (3): 336–343 [Alekhina T.A., Prokudina O.I., Ryazanova M.A., Ukolova T.N., Barykina N.N., Kolpakov V.G. (2007) Typological Characteristics of Behavior in Strains of Rats Bred for Enhancement and Absence of Pendulum Movements. Association with Brain Monoamines. Zh. Vyssh. Nerv. Deiat. 57 (3): 336–343 (in Russ)].
Акулов А.Е., Алехина Т.А., Мешков И.О., Петровский Е.Д., Прокудина О.И., Коптюг И.В., Савелов А.А., Мошкин М.П. (2014) Отбор на кататонический тип реагирования у крыс: исследование межлинейных различий методом магнитно-резонансной томографии. Журн. высш. нерв. деят. 64(4): 439–447 [Akulov A.E., Alekhina T.A., Meshkov I.O., Petrovskiĭ E.D., Prokudina O.I., Koptiug I.V., Savelov A.A., Moshkin M.P. (2014) Selection for catatonic reaction in rats: a study of interstrain differences by magnetic resonance imaging. Zh. Vyssh. Nerv. Deiat. 64 (4):439–447 (in Russ)].
Алехина Т.А., Кожемякина Р.В. (2019) Моделирование фокальных приступов с автоматизмами на крысах с маятникообразными движениями. Бюлл. эксп. биол. мед. 168(8): 300–303 [Alekhina T.A., Kozhemyakina R.V. (2019) Modeling of focal seizures with automatisms in rats with pendulum movements. Bull. exp. biol. med. 168 (2): 300–303. (in Russ)]. https://doi.org/10.1007/s10517-019-04-695-7
Clarac F., Vinay L., Cazalets J.R., Fady J.C., Jamon M. (1998) Role of gravity in the development of posture and locomotion in the neonatal rat. Brain. Res. Rev. 28 (1–2): 35–43https://doi.org/10.1016/S0165-0173(98)00024-1
Eilam D., Golani I. (1988) The ontogeny of exploratory behavior in the house rat (Rattus rattus): the mobility gradient. Develop. Psychobio. 21 (7): 679–710. https://doi.org/10.1002/dev.420210707
Schank C. (2008) The development of locomotor kinematics in neonatal rats: an agent-based modeling analysis in group and individual contexts. J. Theor. Biol. 254: 826–842 https://doi.org/10.1016/j.jtbi.2008.07.024
Manolagas S.C., O’Brien C.A., Almeida M. (2013) The role of estrogen and androgen receptors in bone health and disease. Nat. Rev. Endocrinol. 9 (12): 699–712. https://doi.org/10.1038/nrendo.2013.179
Smith L.B., Walker W.H. (2014) The regulation of spermatogenesis by androgens. Semin. Cell. Dev. Biol. 30:2–13. https://doi.org/10.1016/j.semcdb.2014.02.012
Saad F., Gooren L. (2009) The role of testosterone in the metabolic syndrome: a review. J Steroid Biochem Mol Biol 114 (1–2):40–43. https://doi.org/10.1016/j.jsbmb.2008.12.022
Beverlya B.E.J., Furra J.R., Lambrighta C.S., Wilsona V.S., McIntyrec B.S., Fosterc P.M.D., Travlosc G., Gray Jr. L.E. (2019) In utero exposure to simvastatin reduces postnatal survival and permanently alters reproductive tract development in the Crl:CD(SD) male rat. Toxicol. Appl. Pharmacol. 365: 112–123https://doi.org/10.1016/j.taap.2019.01.001
Korczynski R., Korda P. (1988) Immobility reflex evoked by vertical lifting of the rat. Acta. Neurobiol. Exptl. 48:145–159.
Осадчук Л.В. (2010) Тестикулярная функция у мышей инбредных линий BALB/clac, PT и CBA/Lac. Рос. физиол. журн. им. И.М. Сеченова 96 (2): 183–190 [Osadchuk L.V. (2010) Testicular function in mice of inbred strains BALB/cLac, PT, and CBA/Lac. Russ. J. Physiol. 96 (2):183–190. (in Russ)].
Осадчук Л.В., Клещев М.А., Бакланов А.В., Бажан Н.М. (2016) Тестикулярная функция и липидный обмен у самцов мышей с наследственной предрасположенностью к ожирению. Рос. физиол. журн. им. И.М. Сеченова 102 (3): 340–350 [Osadchuk L.V., Kleschev M.A., Baklanov A.V., Bazhan N.M.. Testicular function and lipid in male mice with hereditary predisposition to obesity. Russ. J. Physiol. 102 (3): 340–350. (in Russ)].
McGrow C.P., Klemm W.R. (1973) Genetic differences in susceptibility of rats to the immobility reflex (“Animal Hypnosis”). Behav. Genetics. 3 (2): 155–162. https://doi.org/10.1007/BF01067655
Kolpakov V.G., Barykina N.N., Alekhina T.A., Chepkasov I.L. (1985) Catalepsy in rats: Its inheritance and relationship to pendulum movements and audiogenic epilepsy. Behav Processes 10 (1–2): 63–76. https://doi.org/10.1016/0376-6357(85)90118-4
Kolpakov V.G., Alekhina T.A., Barykina N.N., Chugui V.F., Popova N.K. (2011) Some Physiological Manifestations of the Activity of the Gene Controlling the Predisposition to Pendulum-Like Movements in Rats. Neurosci. Behav. Physiol. 31 (3): 311–316. https://doi.org/10.1023/a:1010390719547
Барыкина Н.Н., Чугуй В.Ф., Прокудина О.И., Плюснина И.З., Колпаков В.Г. (2007) Подтверждение положительной взаимосвязи маятникообразных движений с аудиогенной эпилепсией, каталепсией и “нервностью” у крыс. Генетика 48 (7):987–993. [Barykina N.N., Chuguy V.F., Prokudina O.I., Plusnina I.P., Kolpakov V.G. (2007) Confirmation of a positive genetic relationship between pendulum movements, audiogenic epilepsy, catalepsy and “nervousness” in rats. Genetica 48 (7):987–993 (in Russ)].
Колпаков В.Г., Барыкина Н.Н., Чугуй В.Ф., Алехина Т.А. (1999) Взаимоотношения между некоторыми формами каталепсии у крыс. Попытка генетического анализа. Генетика 35 (6):807–810. [Kolpakov V.G., Barykina N.N., Chuguĭ V.F., Alekhina T.A. (1999) Relationship between certain forms of catalepsy in rats. An attempt at genetic analysis. Genetika 35 (6):807–810. (in Russ)].
Алехина Т.А., Кожемякина Р.В. (2018) Межлинейные различия по эмоциональным и весовым параметрам у крыс с кататоническим типом реагирования и Вистар. Вавилов. ж. генетики. сел. 22 (4):452–458. [Alehina T.A., Kozhemjakina R.V. (2018) Interlinear differences in emotional and weight parameters in rats with a catatonic type of response and Wistar. Vavilov Journal of Genetics and Breeding 22 (4):452–458. (in Russ)]. https://doi.org/10.18699/VJ18.382
Барыкина Н.Н., Чугуй В.Ф., Алехина Т.А., Рязанова М.А., Уколова Т.Н., Сахаров Д.Г., Колпаков В.Г. (2009) Об-учение крыс, предрасположенных к кататоническим состояниям, в водном тесте Морриса. Журн. высш. нерв. деят. 59 (6):728–735 [Barykina N.N., Chuguy V.F., Alekhina T.A., Ryazanova M.A., Ukolova T.N., Saharov D.G., Kolpakov V.G. (2009) Learning of rats predisposed to catalepsy in Morris water test. Zhurn. vish. nerv. deyat. 59 (6): 728–735. (in Russ)].
Барыкина Н.Н., Чепкасов И.Л., Алехина Т.А., Колпаков В.Г. (1983) Селекция крыс Вистар на предрасположенность к каталепсии. Генетика. 19 (12):2014–2021 [Barykina N.N., Chepkasov I.L., Alekhina T.A., Kolpakov V.G. (1983) Breeding of Wistar rats for predisposition of catalepsy. Genetica. 19 (12):2014–2021. (in Russ)].
Петрова Е.В., Лучкова Т.И., Пироженко А.В. (1992) ЭКоГ крыс с генетической каталепсией. Журн. высш. нерв. деят. 42:1009–1017 [Petrova E.V., Luchkova T.I., Pirozhenko A.V. (1992) The ECoG of rats with genetic catalepsy. Zh. Vyssh. Nerv. Deiat. Im I P Pavlova. 42 (5):1009–1017. (in Russ)].
Алехина Т.А., Пальчикова Н.А., Кожемякина Р.В., Прокудина О.И. (2016) Признаки дестабилизации при отборе на кататонию, проявляющиеся в изменении поведенческих и соматовегетативных параметров у крыс. Вавилов. ж. генетики. сел. 20 (1):74–82 [Alekhina T.A., Palchikova N.A., Kozhemyakina R.V., Prokudina O.I. (2016) Destabilization signs in behavioral and somatovegetable parametres of rats selected for catatonia. Vavilov Journal of Genetics and Breeding 20 (1):74–82. (in Russ)]. https://doi.org/10.18699/VJ16.103
Осадчук Л.В., Алехина Т.А. (2018) Гормональный и метаболический профиль в онтогенезе у самцов крыс при селекции на кататонический тип реагирования. Ж. эволюц. биохим. физиол. 54 (1): 52–59 [Osadchuk L.V., Alekhina T.A. (2018) Developmental profiles of hormonal and metabolic parameters in male rats selected for catatonic type of response. Zh. Evol. Biokhim. Fiziol. 54 (1):52–59. (in Russ)].
Клещев М.А., Алехина Т.А., Осадчук Л.В. (2018). Сперматогенная функция семенников у крыс с наследственной предрасположенностью к проявлению кататонических реакций. Вавилов. ж. генетики. сел. 22 (4):400–405 [M.A. Kleshchev, Alekhina T.A., Osadchuk L.V. (2018) The spermatogenic function testes in rats predisposed to the manifestation of catatonic reactions. Vavilov Journal of Genetics and Breeding 22 (4):400–405. (in Russ)]. https://doi.org/10.18699/VJ18.375
Lenz H. (1970) Pathologische EEG-Befunde bei epileptischen Psychosen, Depression and Schizophrenia. Archiv fur Psychiatrie und Zeitschrift f d ges Neurologie 208:52–60. 1966. https://doi.org/10.1007/BF00341696
Tadokoro Y., Oshima T., Kanemoto K. (2007) Interictal psychoses in comparison with schizophrenia – A prospective study. Epilepsy 48 (12):2345–2351. https://doi.org/10.1111/j.1528-1167.2007.01230.x
Sakakibara E., Nishida T., Sugishita K., Jinde S., Inoue Y., Kiyoto Kasai. K (2012) Acute psychosis during the postictal period in a patient with idiopathic generalized epilepsy: Postictal psychosis or aggravation of schizophrenia? A case report and review of the literature. Epilepsy Behav. 24 (3):373–376. https://doi.org/10.1016/j.yebeh.2012.04.127
Igonina T.N., Alekhina T.A., Palchikova N.A., Prokudina O.I. (2016) Prodromal signs of catatonia are associated with hereditary dysfunction of body systems in rat pups. J Experimental and Integrative Medicine 6 (3):99–108. https://doi.org/10.5455/jeim.270816.or.157
Angelopoulou R., Balla M., Lavranos G., Chalikias M., Kitsos C., Baka S., Kittas C. (2008) Sertoli cell proliferation in the fetal and neonatal rat testis: a continuous phenomenon? Acta Histochem. 110 (4):341–347. https://doi.org/10.1016/j.acthis.2007.10.009
Chen H., Ge R.S., Zirkin B.R. (2009) Leydig cells: From stem cells to aging. Mol. Cell. Endocrinol. 306 (1–2): 9–16. https://doi.org/10.1016/j.mce.2009.01.023
Gehrand A.L., Phillips J., Malott K., Raff H. (2020) Corticosterone, Adrenal, and the Pituitary-Gonadal Axis in Neonatal Rats: Effect of Maternal Separation and Hypoxia. Endocrinology 161 (7):bqaa085. https://doi.org/10.1210/endocr/bqaa085
Teerds K.J., Huhtaniemi I.T. (2015) Morphological and functional maturation of Leydig cells: from rodent models to primates. Hum Reprod Update. 21 (3):310–328. https://doi.org/10.1093/humupd/dmv008
Kilcoyne K.R., Smith L.B., Atanassova N., Macpherson S., McKinnell C., van den Driesche S., Jobling M.S., Chambers T.J., De Gendt K., Verhoeven G., O’Hara L., Platts S., Renato de Franca L., Lara N.L., Anderson R.A., Sharpe R.M. (2014) Fetal programming of adult Leydig cell function by androgenic effects on stem/progenitor cells. Proc. Natl. Acad. Sci. U.S.A. 111 (18): E1924–E1932. https://doi.org/10.1073/pnas.1320735111
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии