

Физиология растений, 2020, T. 67, № 4, стр. 369-377
Сравнительный анализ роста корней разных видов растений на клеточном уровне
Н. В. Жуковская a, Е. И. Быстрова a, Н. Ф. Лунькова a, В. Б. Иванов a, *
a Федеральное государственное бюджетное учреждение науки Институт физиологии растений
им. К.А. Тимирязева Российской академии наук
Москва, Россия
* E-mail: ivanov_vb@mail.ru
Поступила в редакцию 04.12.2019
После доработки 19.12.2019
Принята к публикации 23.12.2019
Аннотация
Проведен анализ параметров роста клеток корней проростков 53 видов однодольных и 78 видов двудольных растений. Измерены приросты корней за сутки во время стационарного роста, диаметры корней, длины меристем и зон растяжения, длины клеток в ряду коры меристем и их число, а также длины клеток, закончивших растяжение. На основании измеренных параметров рассчитаны следующие показатели: скорость роста корней, относительная скорость деления и растяжения клеток, скорости образования клеток и их перехода к растяжению, продолжительность клеточных циклов и время растяжения клеток. Установлено большое сходство параметров, определяющих рост корня на клеточном уровне, в корнях разных видов растений из разных семейств. Различия в скорости роста корней у изученных видов определялись, главным образом, разным числом клеток в одном ряду меристемы и в меньшей степени – длиной клеток, заканчивающих рост, и продолжительностью клеточных циклов. Длина меристемы и число меристематических клеток в одном ряду тесно коррелировали с диаметром корня, в то время как относительные скорости роста меристематических и растягивающихся клеток, а также продолжительность роста клеток растяжением мало коррелировали с диаметром корня.
ВВЕДЕНИЕ
Рост корня происходит за счет образования новых клеток и роста их растяжением. У всех корней, произрастающих в почве в нормальных условиях, растущая часть очень небольшая и состоит из меристемы, где клетки делятся, и зоны растяжения, где они достигают окончательного размера. Скорость роста клеток в зоне растяжения намного выше, чем в меристеме, поэтому скорость роста корня определяется практически ростом растягивающихся клеток. Однако растяжение клеток продолжается недолгое время, поэтому поддержание роста корня в течение длительного времени происходит за счет перехода к растяжению новых клеток, возникающих в процессе деления [1–3].
Рост клеток изучен у небольшого числа объектов: кукурузы [4, 5], тимофеевки [6, 7], конских бобов [8], гороха [9], люпина [1], пшеницы [10, 11], арабидопсиса [12], конского каштана, дуба [13] и др. [1–3, 14, 15]. Этих данных недостаточно, чтобы проанализировать зависимость между отдельными процессами роста и деления клеток и сравнить, чем отличается рост корней на клеточном уровне у растений, различающихся по систематическому положению, толщине корня и др. Известны работы по изучению зависимости скорости роста от ряда параметров, характеризующих рост клеток, но эти исследования проведены на корнях всего нескольких видов [16, 17].
Цель работы – анализ параметров деления и роста клеток и корреляций между ними в корнях проростков 53 видов однодольных и 78 видов двудольных растений для выявления различий в росте корней на клеточном уровне у представителей 19 семейств растений.
МАТЕРИАЛЫ И МЕТОДЫ
Выращивание растений. Были изучены корни проростков 131 вида травянистых растений. Семена, полученные из разных источников, без предварительной обработки проращивали в чашках Петри на смоченной водопроводной водой, проведенной через магистральный бытовой фильтр Ita–25 (Россия), фильтровальной бумаге при температуре 23 ± 2°С в темноте в термостате Binder ED 53 (Германия). Определяли прирост стационарно растущих корней проростков за 24 ч, через несколько дней после прорастания, когда корни росли с постоянной скоростью.
Фиксация материала и подготовка к микроскопированию. Кончики корней, растущих с постоянной скоростью, длиной до 1 см отрезали лезвием бритвы и фиксировали в 70% этаноле в течение 4 ч при комнатной температуре. После фиксации корни ополаскивали дистиллированной водой и переносили сначала на 15 мин в 25% глицерин, затем в 50% глицерин. Корни более 1 мм в диаметре перед фиксацией разрезали вдоль лезвием бритвы, фиксировали в 70% этаноле в течение 4 ч при комнатной температуре и осветляли по методу [18], инкубируя в течение 15 мин в 0.24 н HCl при температуре 57ºС, затем в 7% NaOH и после в 60% этаноле в течение 15 мин в каждом растворе при комнатной температуре. Затем корни последовательно помещали на 5 мин в 40-, 20-, 10- и 5% этанол, после чего переносили на 15 мин в 25% глицерин и далее в 50% глицерин. На временных неокрашенных препаратах в 50% глицерине под микроскопом Olympus CX-41 (Япония) с помощью окулярной линейки измеряли показатели клеточного роста корня.
Результаты измерений показателей, характеризующих рост клеток в корнях разных видов, представлены в Supplementary, Table S1.
Таблица 1
| а. Характеристика роста клеток в первичных корнях однодольных растений | ||||||
| Параметры | Однодольные (n = 53) | Poaceae (n = 44) |
Amaryllidaceae (n = 5) |
Остальные однодольные (n = 4) |
||
| D, мкм | 339 ± 22 | 353 ± 25 | 342 ± 22 | 193 ± 65 | ||
| Lm, мкм | 645 ± 45 | 668 ± 51 | 696 ± 48 | 385 ± 144 | ||
| Le, мкм | 1950 ± 183 | 1861 ± 184 | 3510 ± 845 | 1177 ± 309 | ||
| Le/Lm | 3 ± 0.2 | 2.8 ± 0.2 | 5 ± 0.2 | 3 ± 0.2 | ||
| lm, мкм | 9 ± 0.3 | 8 ± 0.3 | 13 ± 0.7 | 8 ± 0.6 | ||
| lmat, мкм | 154 ± 6 | 155 ± 7 | 177 ± 15 | 126 ± 22 | ||
| lmat/lm | 18 ± 1 | 18 ± 1 | 14 ± 1 | 17 ± 4 | ||
| Nm, кл | 73 ± 5 | 77 ± 5 | 56 ± 7 | 52 ± 21 | ||
| Ne, кл | 23 ± 2 | 22 ± 2 | 39 ± 11 | 18 ± 5 | ||
| Nm/Ne | 3.7 ± 0.2 | 3.9 ± 0.2 | 1.8 ± 0.4 | 3.4 ± 1.5 | ||
| V, мм/ч | 0.64 ± 0.06 | 0.72 ± 0.06 | 0.36 ± 0.04 | 0.2 ± 0.07 | ||
| Km, ч–1 | 0.05 ± 0.002 | 0.06 ± 0.002 | 0.04 ± 0.003 | 0.04 ± 0.01 | ||
| Ke, ч–1 | 0.39 ± 0.03 | 0.44 ± 0.03 | 0.12 ± 0.03 | 0.17 ± 0.03 | ||
| Ke/Km | 7.1 ± 0.5 | 7.7 ± 0.5 | 3.4 ± 0.8 | 6.2 ± 2.3 | ||
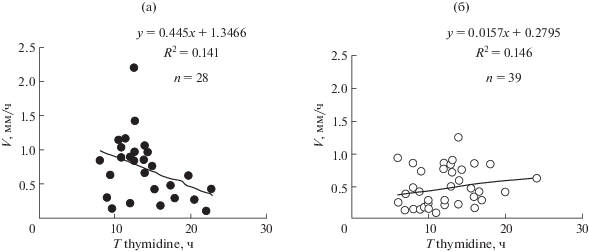
| Tthymidine, ч | 13.9 ± 0.7 (n = 28) | 12.4 ± 0.5 (n = 22) | 18.7 ± 1.3 | – | ||
| Vme, кл/ч | 4 ± 0.3 | 4 ± 0.3 | 2 ± 0.3 | 1.4 ± 0.3 | ||
| te, ч | 8 ± 1 | 5.7 ± 0.5 | 20.0 ± 4.4 | 14.7 ± 3.7 | ||
| б. Характеристика роста клеток в первичных корнях двудольных | ||||||
| Параметры | Двудольные (n = 78) | Asteraceae (n = 18) |
Brassicaceae (n = 12) |
Fabaceae (n = 19) |
Solanaceae (n = 10) |
Остальные двудольные (n = 19) |
| D, мкм | 331 ± 22 | 221 ± 25 | 245 ± 37 | 543 ± 49 | 281 ± 50 | 314 ± 27 |
| Lm, мкм | 640 ± 45 | 406 ± 62 | 497 ± 86 | 1031 ± 96 | 556 ± 108 | 606 ± 65 |
| Le, мкм | 1862 ± 155 | 1051 ± 153 | 1506 ± 278 | 2902 ± 295 | 1590 ± 437 | 1959 ± 366 |
| Le/Lm | 2.9 ± 0.2 | 2.6 ± 0.4 | 3 ± 0.4 | 2.9 ± 0.25 | 2.9 ± 0.5 | 3.2 ± 0.4 |
| lm, мкм | 9 ± 0.2 | 9 ± 0.4 | 7 ± 0.4 | 11 ± 0.5 | 9 ± 1 | 9 ± 0.4 |
| lmat, мкм | 132 ± 4 | 109 ± 6 | 130 ± 14 | 148 ± 7 | 125 ± 11 | 143 ± 9.5 |
| lmat/lm | 15 ± 0.6 | 12 ± 0.5 | 17.5 ± 2 | 14 ± 0.8 | 14 ± 1.7 | 15.62 ± 1.4 |
| Nm, кл | 67 ± 4 | 44 ± 5 | 65 ± 10 | 93 ± 6 | 62 ± 13 | 64.67 ± 7 |
| Ne, кл | 25 ± 2 | 17 ± 2 | 20 ± 3 | 36 ± 3 | 21 ± 5 | 24.68 ± 3 |
| Nm/Ne | 3 ± 0.1 | 2.7 ± 0.2 | 3.4 ± 0.3 | 2.8 ± 0.2 | 3.3 ± 0.4 | 3.00 ± 0.3 |
| V, мм/ч | 0.49 ± 0.04 | 0.31 ± 0.05 | 0.57 ± 0.1 | 0.61 ± 0.07 | 0.38 ± 0.08 | 0.55 ± 0.08 |
| Km, ч–1 | 0.05 ± 0.002 | 0.06 ± 0.002 | 0.06 ±0.01 | 0.04 ± 0.002 | 0.05 ± 0.002 | 0.06 ± 0.002 |
| Ke, ч–1 | 0.31 ± 0.02 | 0.33 ± 0.04 | 0.40 ± 0.04 | 0.22 ± 0.02 | 0.31 ± 0.04 | 0.31 ± 0.03 |
| Ke/Km | 5.6 ± 0.2 | 5.1 ± 0.5 | 6.5 ± 0.6 | 5.3 ± 0.5 | 6.1 ± 0.7 | 5.69 ± 0.5 |
| Tthymidine, ч | 12.2 ± 0.6 (n = 39) | 10 ± 0.6 (n = 13) |
– | 16.5 ± 1.5 (n = 7) | 10.1 ± 1.1 (n = 4) | 12.6 ± 1.0 (n = 14) |
| Vme, кл/ч | 3.5 ± 0.2 | 2.8 ± 0.3 | 4 ± 0.6 | 4 ± 0.4 | 2.9 ± 0.4 | 3.61 ± 0.4 |
| te, ч | 8.4 ± 0.6 | 7.9 ± 1.1 | 5.5 ± 0.5 | 10.8 ± 1 | 8.6 ± 2 | 8.15 ± 1 |
Статистическая обработка данных. Эксперименты были проведены в двух независимых повторностях. Каждая повторность включала по 8–10 корней каждого вида. Полученные данные представлены как средние значения и их стандартные ошибки. Обработку данных проводили с помощью программы MS Excel 2007 и SigmaPlot 13.
РЕЗУЛЬТАТЫ
Сравнение показателей, характеризующих рост корня на клеточном уровне в корнях разных видов
На рисунке 1 представлена схема клеточной организации роста корня. Для проведения клеточного анализа нами были измерены следующие показатели: 1) длина меристемы (Lm); 2) длина меристематических клеток (lm); 3) длина зоны растяжения (Le); 4) длина закончивших рост клеток коры (lmat); 5) диаметр корня на границе меристемы и зоны растяжения (D). Полученные данные позволили вычислить:
Рис. 1.
Схематическое изображение растущей части корня. Lm – длина меристемы; Le – длина зоны растяжения; Nm – число меристематических клеток; Ne – число растягивающихся клеток.
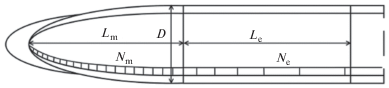
1) число меристематических клеток в продольном ряду коры (Nm):
(1)
${{N}_{{\text{m}}}} = {{{{L}_{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{L}_{{\text{m}}}}} {{{l}_{{\text{m}}}}}}} \right. \kern-0em} {{{l}_{{\text{m}}}}}}$2) число растягивающихся клеток в одном продольном ряду коры (Ne):
(2)
${{N}_{{\text{e}}}} = {{2{{L}_{{\text{e}}}}} \mathord{\left/ {\vphantom {{2{{L}_{{\text{e}}}}} {({{l}_{{\text{m}}}} + {{l}_{{{\text{mat}}}}})}}} \right. \kern-0em} {({{l}_{{\text{m}}}} + {{l}_{{{\text{mat}}}}})}}$3) скорость роста корня (V):
(3)
$V = {{\Delta L} \mathord{\left/ {\vphantom {{\Delta L} {\Delta t}}} \right. \kern-0em} {\Delta t}},$где ΔL – прирост корня за период Δt, равный в опытах 24 ч.
4) продолжительность митотического цикла (Т) [2, 19, 20]:
(4)
$T = {{\ln 2{{N}_{{\text{m}}}}{{l}_{{{\text{mat}}}}}} \mathord{\left/ {\vphantom {{\ln 2{{N}_{{\text{m}}}}{{l}_{{{\text{mat}}}}}} V}} \right. \kern-0em} V}$5) среднюю относительную скорость роста меристематических клеток:
6) среднюю относительную скорость роста растягивающихся клеток (Ke) [2]. (Эта формула применима, т. к. скорость роста много больше, чем скорость роста меристемы. В корнях нет скользящего роста, поэтому и относительная скорость роста клеток всех тканей одинакова на одном и том же расстоянии от кончика корня.):
(6)
${{K}_{{\text{e}}}} = {V \mathord{\left/ {\vphantom {V {{{L}_{{\text{e}}}}}}} \right. \kern-0em} {{{L}_{{\text{e}}}}}}$7) продолжительность роста растяжением (te) [2]:
(7)
${{t}_{{\text{e}}}} = {{2{{L}_{{\text{e}}}}} \mathord{\left/ {\vphantom {{2{{L}_{{\text{e}}}}} V}} \right. \kern-0em} V}$Анализ данных, полученных в ходе проведенных измерений и расчетов, показал большое сходство между параметрами, определяющими рост первичных корней на клеточном уровне в корнях разных видов как однодольных, так и двудольных травянистых растений, в том числе по семействам (табл. 1а, б). Среди изученных семейств у двудольных несколько отличалось сем. Fabaceae, у которого отмечали больший диаметр первичного корня и, соответственно, большую длину зон деления и растяжения, чем у представителей других семейств.
У корней как однодольных, так и двудольных были выявлены следующие общие закономерности в организации клеточного роста: длины зоны растяжения (Le) примерно в 3 раза больше, чем длины меристемы (Lm), а число растягивающихся клеток (Ne) в ряду примерно в 3 раза меньше, чем число меристематических клеток (Nm). Длина закончивших рост клеток (lmat) примерно в 15 раз больше, чем длина меристематических клеток (lm). Скорость роста корней (V) была чуть больше у проростков однодольных по сравнению с двудольными. Относительная скорость растяжения клеток (Ke) составляла в среднем 0.35 ч–1, что в 5–7 раз больше относительной скорости образования меристематических клеток (Km), которая в большинстве корней близка к 0.05 ч–1. Продолжительность митотического цикла (Т) во всех корнях составляла примерно 14 ч. Продолжительность роста растяжением (te) в корнях проростков составляла около 8 ч, что примерно в 2 раза короче, чем средняя T.
Корреляции между величинами разных показателей, характеризующих рост клеток в корне
Прежде чем проанализировать, за счет каких процессов на клеточном уровне различается скорость роста корней у разных видов, мы сопоставили корреляции между величинами отдельных параметров, характеризующих рост клеток в корне (табл. 2). Скорость роста корней у разных видов заметно различалась. Более толстые корни росли быстрее, чем более тонкие. Коэффициенты корреляции между диаметром корня и скоростью роста были 0.85 у проростков однодольных и 0.74 – у двудольных растений. При этом относительная скорость растяжения, длительность цикла и продолжительность роста растяжением не коррелировали с толщиной корня. Различия в скорости роста корней разного диаметра определялись тем, что длины меристемы и зоны растяжения возрастали по мере увеличения диаметра корня. Коэффициенты корреляции между диаметром корня и длиной меристемы составили 0.95 у корней проростков однодольных и 0.99 – у двудольных. Число меристематических клеток высоко коррелировало с диаметром в корнях (0.84 у однодольных и 0.86 у двудольных). В более толстых корнях была длиннее и зона растяжения.
Таблица 2.
Коэффициенты корреляции между отдельными показателями роста для первичных корней однодольных и двудольных растений
| Параметры | Однодольные проростки | Двудольные проростки | ||||||
|---|---|---|---|---|---|---|---|---|
| D | Lm | lmat | V | D | Lm | lmat | V | |
| D | 1 | 0.95 | 0.64 | 0.85 | 1 | 0.99 | 0.55 | 0.74 |
| Lm | 0.95 | 1 | 0.72 | 0.83 | 0.99 | 1 | 0.57 | 0.78 |
| lm | 0.34 | 0.47 | 0.46 | 0.10* | 0.67 | 0.63 | 0.33 | 0.35 |
| Le | 0.50 | 0.65 | 0.55 | 0.46 | 0.69 | 0.73 | 0.70 | 0.77 |
| lmat | 0.64 | 0.72 | 1 | 0.62 | 0.55 | 0.57 | 1 | 0.74 |
| V | 0.85 | 0.83 | 0.62 | 1 | 0.74 | 0.78 | 0.74 | 1 |
| Ke | 0.23* | 0.10* | 0.10* | 0.41 | –0.13* | –0.14* | –0.11* | 0.08* |
| Ke/Km | 0.33 | 0.21* | 0.36 | 0.37 | 0.09* | 0.08* | 0.11* | 0.06* |
| Tthymidine | –0.03* | 0.01* | 0.24* | –0.38 | 0.41 | 0.41 | 0.31 | 0.21* |
| Km | –0.10* | –0.21* | –0.42 | 0.19* | –0.29 | –0.31 | –0.27 | 0.04* |
| Nm | 0.84 | 0.90 | 0.62 | 0.87 | 0.86 | 0.90 | 0.61 | 0.81 |
| Ne | 0.34 | 0.45 | 0.26* | 0.30 | 0.65 | 0.70 | 0.48 | 0.66 |
| Vm | 0.70 | 0.67 | 0.28 | 0.89 | 0.71 | 0.75 | 0.48 | 0.92 |
| te | –0.23* | –0.10* | –0.10* | –0.38 | –0.01* | –0.001* | 0.01* | –0.16* |
Показатели относительной скорости роста растяжением клеток (Ke) в корнях были близки у однодольных и двудольных растений. При переходе клеток к растяжению относительная скорость роста клеток растяжением была в 6–7 раз выше, чем в меристеме. Относительная скорость роста растяжением (Ke) практически не зависела от размера меристемы (Nm).
Коэффициенты корреляции lmat от lm для однодольных составили 0.59, для двудольных – 0.33. Длина закончивших рост клеток не зависела от скорости растяжения (Ke) и его продолжительности (te).
Интересно отметить, что длина меристемы и число меристематических клеток коррелировали с длиной закончивших рост клеток. Коэффициенты корреляции между lmat и Nm были достоверны для однодольных и двудольных (0.53 и 0.60). Пока механизм этой корреляции непонятен.
Зависимость скорости роста корня от отдельных показателей, определяющих рост и деление клеток
Если корень растет с постоянной скоростью, как в наших опытах, то скорость его роста определяется скоростью образования клеток в меристеме (Vn), которая равна для одного ряда клеток:
(8)
${{V}_{{\text{n}}}} = {{\ln 2{{N}_{{\text{m}}}}} \mathord{\left/ {\vphantom {{\ln 2{{N}_{{\text{m}}}}} T}} \right. \kern-0em} T}$Скорость роста всего корня (V) равна:
Таким образом, скорость роста корня (V) определяется:
(10)
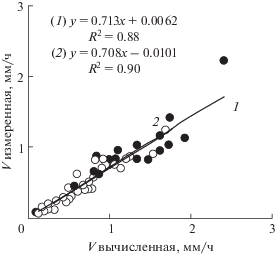
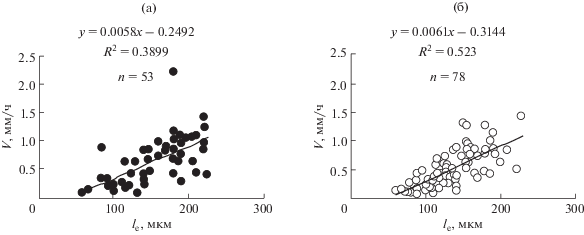
$V = {{({\text{ln}}2{{N}_{{\text{m}}}}{{l}_{{{\text{mat}}}}})} \mathord{\left/ {\vphantom {{({\text{ln}}2{{N}_{{\text{m}}}}{{l}_{{{\text{mat}}}}})} T}} \right. \kern-0em} T}$Для экспериментальной проверки формулы (10) [20] мы измерили в корнях 69 видов, для которых были опубликованы данные по Т, полученные независимым “тимидиновым” методом [21, 22], Nm и lmat и вычислили ожидаемое значение скорости роста корня, которое совпало с измеренными значениями скорости роста корней в наших опытах (рис. 2). Приложимость этой формулы для описания роста корней позволяет решать обратную задачу – вычислить Т по измерениям V, Nm и lmat. Как нами показано, рассчитанные этим методом и опубликованные значения T, полученные с помощью “тимидинового” метода, совпадают для подавляющего большинства видов растений [20].
Рис. 2.
Сравнение вычисленных значений скорости роста корня (V) (V = (ln2Nmlmat)/T) и измеренных. Nm – число меристематических клеток в ряду, Т – длительность клеточного цикла (“тимидиновый” метод), lmat – длина закончивших рост клеток; 1 – однодольные проростки (черные кружки), 2 – двудольные проростки (белые кружки) [20].
С другой стороны, скорость роста клеток в зоне растяжения значительно выше, чем в меристеме (табл. 1а, б). Поэтому V всего корня также равна:
Надо иметь в виду, что величина Le зависит от числа клеток в меристеме, скорости перехода меристематических клеток к растяжению (Vm) и времени растяжения (te). Для корня, растущего с постоянной скоростью, длина зоны растяжения обратно пропорциональна относительной скорости растяжения. Поэтому для описания роста корня может быть использована формула (9).
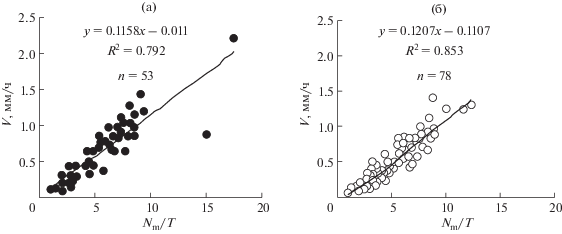
Как видно на рисунках 3–6, V возрастает с увеличением Nm, еще более резко возрастает с увеличением отношения Nm/T (скорость образования клеток), несколько слабее от длины закончивших рост клеток (lmat) и еще слабее от продолжительности митотического цикла (T). То есть наиболее сильно V коррелирует с соотношением Nm/T и несколько слабее с Nm. Слабее выражена зависимость V от lmat и еще слабее от T. Причем для двудольных корней эта зависимость не достоверна. Последний результат является удивительным, т. к. кажется очевидным, что чем быстрее делятся клетки, тем быстрее должен расти корень. С другой стороны, если посмотреть зависимость V от величин Le и Ke, то V заметно коррелирует с Le, однако у двудольных корреляция между V и Ke не достоверна. У корней, растущих с постоянной скоростью, скорость выхода клеток (Vm) из зоны меристемы и зоны растяжения равна скорости образования клеток (Vn).
Рис. 3.
Зависимость скорости роста корня (V) от числа клеток меристемы (Nm). а – однодольные проростки, б – двудольные проростки.
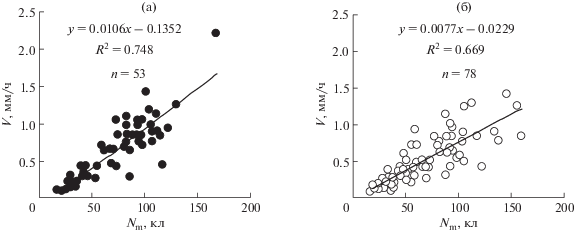
Рис. 4.
Зависимость скорости роста корня (V) от скорости образования клеток (Nm/T). а – однодольные проростки, б – двудольные проростки.
ОБСУЖДЕНИЕ
К настоящему времени опубликовано мало работ по измерениям, которые характеризуют рост корней на клеточном уровне. Однако, как показали полученные нами данные, значительное увеличение числа изученных видов не меняет уже сложившихся представлений об особенностях организации роста корня на клеточном уровне.
У всех корней, нормально растущих в почве, зона роста не превышает сантиметра, а часто даже меньше. В отличие от них, у воздушных корней длина зоны роста во много раз больше [23]. Зона роста состоит из меристемы, где клетки делятся, и зоны растяжения, в начале которой могут встречаться единичные деления клеток у некоторых корней. В растущих междоузлиях и листьях двудольных деления клеток могут происходить и на этапе роста растяжением. Однако относительная скорость роста растяжением в корнях намного выше, чем в большинстве надземных органов [24]. Растягивающиеся клетки корней увеличиваются примерно в 15–17 раз за 7–10 ч (табл. 1а, б), тогда как растягивающиеся клетки быстро растущих колеоптилей удлиняются за сутки примерно в 2 раза.
Корни проростков разных видов резко различаются по скорости роста. Полученные нами данные позволили проанализировать, за счет каких процессов на клеточном уровне скорость роста корней проростков отличается у разных видов. Ранее этот вопрос ставился в литературе, однако изучали корни лишь нескольких видов. Van’t Hof [16] на основании изучения корней 3 видов растений показал, что различия в скорости роста определяются разной скоростью пролиферации клеток в ряду и разной длиной заканчивающих рост клеток. Gazques и Beemster [17], проанализировав опубликованные данные для корней 6 видов, выращиваемых в разных условиях, также сделали вывод о том, что различия в их размерах (следовательно, и скорости роста) определяются разной скоростью образования клеток в меристемах и разной длиной зрелых клеток. При этом они отметили, что различия в скорости образования клеток определяются в большей степени различиями в числе делящихся клеток, а не различиями в продолжительности митотических циклов. Отличия же в длине закончивших рост клеток в корнях разных видов они объяснили разной продолжительностью роста растяжением.
В таблице 2 и на рисунках 3–6 представлены коэффициенты корреляций между основными параметрами, от которых зависит скорость роста корней (V) (см. формулы 8–11). Величина V высоко коррелировала со скоростью образования клеток (Vn = ln2Nm/T) и значительно слабее с длиной заканчивающих рост клеток (lmat). При этом различия в скорости образования клеток в значительно большей степени зависели от различий в числе меристематических клеток, чем продолжительностях митотических циклов (рис. 3, 6). Это подтверждает выводы выше цитированных работ. До сих пор мало известно о механизмах, контролирующих длину заканчивающих рост клеток. В наших измерениях оказалось, что lmat высоко коррелировала с длиной меристемы и числом меристематических клеток в ряду, причем механизм этой корреляции не ясен. Она также заметно коррелировала с диаметром корня и длиной меристематических клеток, но не коррелировала с относительной скоростью растяжения и продолжительностью роста растяжением.
Полученные данные показывают, что различия в скорости роста корней в существенной мере зависят от числа меристематических клеток в ряду и длины меристемы. До сих пор механизмы контроля длины меристемы остаются мало выясненными. В данной работе и ранее [25] мы показали, что в корнях есть четкая зависимость между диаметром корня (D) и длиной меристемы (Lm). При этом отношение Lm/D в корнях разных проростков близко к 2 [26]. Можно предположить, что зависимость Lm от D определяется зависимостью скорости притока необходимых для функционирования и роста меристемы веществ от диаметра корня. Число клеточных рядов в меристеме определяется в самом ее начале и зависит как от исходной величины зародышевого корня в семени, так и от делений клеток, приводящих к увеличению числа рядов, которые формируются в начале меристемы. В рамках данной статьи мы не можем обсуждать возможные механизмы гормональной регуляции этих процессов в связи с отсутствием достаточного материала. По-видимому, в начале меристемы закладываются процессы, определяющие время жизни клеток в меристеме [3] и через них ее размер. Процессы именно в этой области меристемы имеют большее значение в регуляции размера меристемы, чем сам переход клеток к растяжению в переходной зоне между меристемой и зоной растяжения, так как время жизни клеток в меристеме детерминируется уже значительно ближе к кончику корня и скорость перехода клеток к растяжению зависит от соотношения между временем жизни клеток в меристеме и скоростью их делений, пока они еще находятся в меристеме.
Число меристематических клеток на протяжении одного ряда у корней изученных видов колебалось от 10 до 170. Это означает, что клетки после отделения от первых клеток на границе покоящегося центра проходят небольшое число циклов делений (p), так как продолжительность митотических циклов постоянна на протяжении меристемы и Nm = 2p + 1. Следовательно, время жизни клеток в меристеме, равное произведению Т × p, невелико и не превышает нескольких суток [3]. Это показывает, что в меристеме клетки сменяются, и ее сохранение в относительно неизменном виде определяется аналогичными изменениями во времени новых поколений клеток, образовавшихся в результате делений инициальных клеток. Такая быстрая сменяемость клеток и их относительно короткое время жизни в меристеме является характерной особенностью клеточной организации роста корня по сравнению с надземными органами.
В данной работе были изучены корни проростков травянистых растений. Представляет большой интерес провести сравнительное изучение корней более взрослых растений и придаточных корней, а также корней деревьев и кустарников.
Авторы благодарят Н. В. Обручеву за помощь и ценные советы при обсуждении полученных результатов. Мы очень благодарны всем, кто предоставил семена, использованные в этом исследовании: Научно-исследовательский институт растениеводства им. Вавилова в Санкт-Петербурге, Россия; Главный ботанический сад им. Н. В. Цицина РАН, Москва, Россия; Ботанический институт им. В. Л. Комарова РАН, Санкт-Петербург, Россия; Королевские ботанические сады Кью, Великобритания; Ботанический сад Боннского университета, Бонн, Германия; Институт генетики растений и растениеводства Ассоциации Лейбница, Гатерслебен, Германия, и др.
Работа частично поддержана грантом Российского фонда фундаментальных исследований (№ 18-04-00918а) и Министерством науки и высшего образования РФ в рамках государственного задания (номер темы АААА-А19-119040290058-5).
Авторы Жуковская Н.В. и Иванов В.Б. внесли одинаковый вклад в эту работу.
Авторы заявляют об отсутствии конфликта интересов. Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов исследований.
Список литературы
Обручева Н.В. Определение скорости деления и растяжения клеток корня // Физиология растений. 1964. Т. 11. С. 746.
Иванов В.Б. Клеточные основы роста растений. Москва: Наука, 1974. 223 с.
Иванов В.Б. Клеточные основы роста растений. Москва: Наука, 2011. 104 с.
Erickson R.O., Sax K.B. Elemental growth rate of the primary root of Zea mays // Proc. Amer. Philos. Soc. 1956a. V. 100. P. 487.
Erickson R.O., Sax K.B. Rates of cell division and cell elongation in the growth rate of the primary root of Zea mays // Proc. Amer. Philos. Soc. 1956b. V. 100. P. 499.
Avers Ch. J. An analysis of differences in growth rate of trichoblasts and hairless cells in the root epidermis of Phleum pretense //Amer. J. Bot. 1957. V. 44. P. 686.
Goodwin R.H., Avers C.J. Studies of roots. III. An analysis of root growth in Phleum pretense using photomicrographic records // Amer. J. Bot. 1956. V. 449. P. 1053.
Gray L.H., Scholes M.E. The effects of ionizing radiations on the broad bean root. VIII. Growth rate studies and histological analysis // Brit. J. Radiol. 1951. V. 24. P. 348-352.
Rost T.L., Jones T.J. Distribution and relationship of cell division and maturation events in Pisum sativum (Fabaceae) seedling root // Amer. J. Bot. 1988. V. 42. P. 529-540.
Hejnovicz Z. Growth and cell division in the apical meristem of wheat roots // Physiol. Plant. 1959. V. 12. P. 124.
Демченко Н.П. Продолжительность митотического цикла, его периодов и митоза у клеток дерматогена и периблемы корней пшеницы // Цитология. 1976. Т. 43. С. 16.
Beemster G.T.S., Baskin T.I. STUNTED PLANT 1 mediates effects of cytokinin, but not auxin, on cell division and expansion in the root Arabidopsis // Plant Physiol. 2000. V. 124. P. 1718.
Obroucheva N.V. Physiology of growing root cells. In the development and function of roots // London: Acad. Press, L., 1975. P. 279.
Обручева Н.В. Физиология растущих клеток корня. М.: Наука, 1965. 111 с.
Иванов В.Б. Пролиферация клеток в растениях // Итоги науки и техники. Сер. “Цитология”. М.: ВИНИТИ, 1987. 220 с.
Van’t Hof J. Studies on the relationships between cell population and growth kinetics of root meristems // Exper. Cell Res. 1967. V. 46. P. 335.
Gázquez A., Beemster G.T.S. What determines organ size differences between species? A meta-analysis of the cellular basis // New Phytologist. 2017. V. 215. P. 299.
Malamy J.E., Benfey P.N. Organization and cell differentiation in lateral roots of Arabidopsis thaliana // Development. 1997. V. 124. P. 33.
Ivanov V.B., Dubrovsky J.G. Estimation of the cell-cycle duration in the root apical meristem: a model of linkage between cell-cycle duration, rate of cell production, and rate of root growth // Int. J. Pl. Sci. 1997. V. 158. P. 757.
Zhukovskaya N.V., Bystrova E.I., Dubrovsky J.G., Ivanov V.B. Global analysis of an exponential model of cell proliferation for estimation of cell cycle duration in the root apical meristem of angiosperms // Annals Bot. 2018. V. 122. P. 811.
Grif V.G., Ivanov V.B., Machs E.M. Cell cycle and its parameters in flowering plants // Cytologia. 2002. V. 44. P. 936.
Francis D., Davies M.S., Barlow P.W. A strong nucleotypic effect on the cell cycle regardless of ploidy level // Annals Bot. 2008. V. 101. P. 747.
Еськов А.К., Жуковская Н.В., Быстрова Е.И., Орлова Ю.В., Антипина В.А., Иванов В.Б. Рост воздушных корней с длинной зоной растяжения на примере полуэпифита MONSTERA DELICIOSA // Физиология растений. 2016. Т. 63. С. 834.
Silk W.K. Quantitative descriptions of development // Ann. Rev. Plant Physiol. 1984. V. 35. P. 479.
Жуковская Н.В., Быстрова Е.И., Иванов В.Б. Зависимость размеров меристематических и закончивших рост клеток от содержания ДНК в расчете на гаплоидное число хромосом // Онтогенез. 2016. Т. 47. С. 346.
Быстрова Е.И., Жуковская Н.В., Иванов В.Б. Зависимость процессов роста и деления клеток в корне от его диаметра // Онтогенез. 2018. Т. 49. С. 91.
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений