

Физиология растений, 2020, T. 67, № 6, стр. 599-615
Реакции растений на кратковременные ежесуточные понижения температуры: феноменология и механизмы
А. Ф. Титов a, Т. Г. Шибаева a, *, Е. Н. Икконен a, Е. Г. Шерудило a
a Институт биологии Карельского научного центра Российской академии наук,
Федеральный исследовательский центр “Карельский научный центр РАН”
Петрозаводск, Россия
* E-mail: shibaeva@krc.karelia.ru
Поступила в редакцию 07.04.2020
После доработки 25.04.2020
Принята к публикации 25.04.2020
Аннотация
В обзоре представлен анализ и обобщение литературных данных и собственных работ, показывающих наличие принципиальных различий в реакциях растений на низкотемпературные воздействия двух разных типов: а) длительное постоянное и б) кратковременные ежесуточно повторяющиеся (дроп-воздействия). Рассмотрены имеющиеся в литературе данные об особенностях реакции растений на дроп-воздействия в зависимости от их параметров (интенсивности, продолжительности, времени действия в суточном цикле), а также влияние факторов внешней среды на реакцию растений на дроп-воздействия. Проанализированы сведения об особенностях реакции на дроп-воздействия растений, принадлежащих к разным по отношению к температуре и свету группам. Обсуждаются физиолого-биохимические механизмы реакции растений на дроп-воздействия.
ВВЕДЕНИЕ
Растения, не обладая способностью активно перемещаться в пространстве и будучи пойкилотермными организмами, характеризуются выраженной зависимостью от температурных условий среды обитания. Неслучайно роль температуры в жизни растений изучается многие десятилетия, и за это время опубликованы сотни работ, посвященных тем или иным аспектам реакции растительного организма на действие данного фактора. Чаще всего в центре внимания оказываются вопросы низкотемпературной адаптации и/или низкотемпературного повреждения растений [1, 2]. Но и в том и в другом случае, как правило, изучаются эффекты продолжительного действия (дни, недели) низкой температуры на растения, которая в ходе эксперимента сохраняется неизменной. Вместе с тем во многих регионах, и не только расположенных в условиях высоких широт, рост растений даже в период активной вегетации происходит на фоне постоянных и значительных флуктуаций температуры в суточном цикле, причем очень часто с выходом за пределы оптимума. Более того, при относительно высоких значениях среднесуточной температуры здесь довольно часто наблюдаются резкие понижения температуры в ночное время или в предутренние часы, что прослеживается даже в летние месяцы (рис. 1). В суточном цикле такое воздействие низких температур на растения оказывается непродолжительным (в среднем 2–4 ч). Затем оно сменяется достаточно продолжительным периодом действия в суточном цикле оптимальных и околооптимальных температур, поэтому их влияние не всегда очевидно и в силу этого долго не привлекало должного внимания исследователей. Однако с течением времени стали накапливаться данные, указывающие на то, что кратковременные ежесуточно повторяющиеся воздействия низких субповреждающих температур способны оказывать на растения хорошо выраженный морфогенетический эффект, который проявляется в торможении линейного роста растений [3, 4]. Эти наблюдения и соответствующие исследования привели к разработке агротехнического приема, который называется “temperature drop” в Европе и “temperature dip”, или “cool morning pulse” в США. Ежесуточные кратковременные понижения температуры (a daily short-term temperature drop) стали достаточно широко применяться для получения компактной рассады овощных культур и клумбовых растений, а также при выращивании цветочных растений в тепличных условиях. С этой целью температуру обычно снижают на 5−15°С от ее оптимальных значений на 2–3 ч в конце ночи. Во многих случаях благодаря этому удается полностью или частично избежать применения ретардантов (синтетических веществ, замедляющих рост стебля в высоту) [5] (рис. 2, табл. 1).
Рис. 1.
Максимальные и минимальные значения температуры в сутках в г. Москве (55°45′ с.ш.), Санкт-Петербурге (59°57′ с.ш.) и Петрозаводске (61°47′ с.ш.) в летние месяцы 2017 г. (условно принятого в качестве “типичного”) (метеоданные сайта https://nuipogoda.ru/).
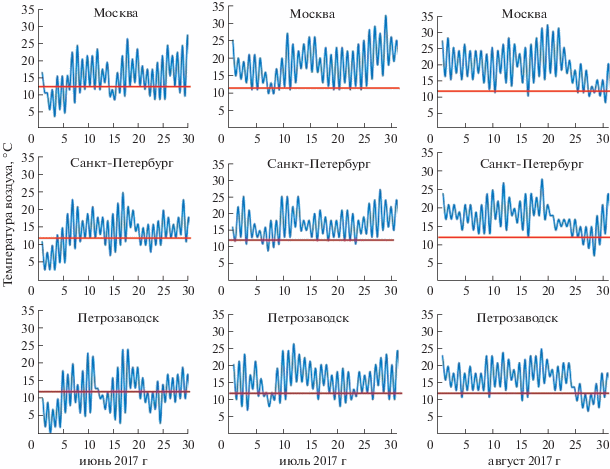
Рис. 2.
Растения молочая (Euphorbia pulcherrima, ”Starlight”), выращенные при одинаковой температуре дня и ночи 19/19°С (слева), при повышении температуры на 2 ч в начале дня до 25°С (в центре) или понижении температуры на 2 ч до 13°С (справа) [6].

Таблица 1.
Эффекты дроп-воздействий на рост и развитие растения
| Вид растения | Дроп-воздействие, °С* | Эффекты | Источник литературы |
|---|---|---|---|
| Овощные, бахчевые и масличные культуры | |||
| Brassica oleracea var. capitataf. alba L. | 23/20 → 5 20 → 6 |
Уменьшение высоты растения; отсутствие влияния на биомассу | 7, 8 |
| Brassica napus L. | 16 → 6 | Уменьшение высоты растения | 7 |
| Сapsicum annuum L. | 23/20 → 9 | Уменьшение высоты растения; отсутствие влияния на биомассу и скорость развития | 8 |
| Citrullus lanatus (Thunb.) Matsum. &Nakai | 29/25 → 2 | Уменьшение площади листьев, задержка цветения, уменьшение биомассы, урожая | 9 |
| Cucumis melo L. | 25/20 → 2 | Задержка цветения, уменьшение биомассы и урожая | 10, 11 |
| Cucumis sativus L. | 23 → 12, 23 → 9 22 → 16, 22 → 12 21 → 15, 21 → 11 |
Уменьшение высоты растения, длины черешков; отсутствие влияния на ранний урожай и качество | 12–19 |
| Pisum sativum L. | 21 → 13 20 → 12 |
Уменьшение высоты растения | 7, 20 |
| Solanum lycopersicum L. | 29 → 2, 22 → 16, 22 → 13, 21 → 12 | Уменьшение высоты растения, длины черешков; отсутствие влияния на ранний урожай и качество | 16, 21–23 |
| Горшечные цветочные культуры | |||
| Begonia x hiemalis Fortsch | 20 → 14, 19 → 16 19 → 13, 18 → 12 |
Уменьшение высоты, длины черешков, цветоносов, увеличение количества цветков | 24–28 |
| Calceolaria x herbeo-hybrida Voss | 19 → 12 | Уменьшение высоты растения, отсутствие влияния на сроки цветения | 29 |
| Campanula isophylla Moretti | 19 → 13 | Уменьшение высоты растения; отсутствие влияния на биомассу и сроки цветения | 30 |
| Chrysanthemum L. sp. | 21 → 15 18 → 10 18 → 8 | Уменьшение высоты растения; отсутствие влияния на биомассу; ускорение цветения | 31, 32 |
| Euphorbia pulcherrima Willd. ex Klotzseh | 24 → 16, 24 → 8 20 → 14, 19 → 13 18 → 10 |
Уменьшение высоты растения, длины черешков; отсутствие влияния на биомассу | 24, 28, 31, 33, 34 |
| Kalanchoe blossfeldiana Poelln. | 19 → 12 | Увеличение длины стебля и цветоносов; отсутствие влияния на сроки цветения | 29, 31 |
| Lilium longiflorum Thunb. | 20 → 12 | Уменьшение высоты растения | 3 |
| Senecio cruentus (Masson ex L′Hér.) DC. | 19 → 12 | Отсутствие влияния на высоту растения | 29 |
| Sinningia speciosa (Lodd. ex Ker Gawl) Hiern | 19 → 12 | Отсутствие влияния на высоту растения | 29 |
| Клумбовые растения | |||
| Begonia х tuberhybrida pendula | 19 → 12 | Уменьшение высоты растения, отсутствие влияния на биомассу и сроки цветения | 29 |
| Fuchsia x hybrida Hort. ex Vilm. | 21 → 15, 20 → 14 18 → 12 |
Уменьшение высоты растения; отсутствие влияния на биомассу | 28, 34 |
| Impatiens walleriana Hook.f. | 19 → 12 | Отсутствие влияния на высоту растения, биомассу и сроки цветения | 29 |
| Impatiens hawker W. Bull | 21 → 15 20 → 14 |
Уменьшение высоты растения | 28 |
| Pelargonium L′Hér ex Ait. | 19 → 12 18 → 2 |
Уменьшение высоты растения; отсутствие влияния на биомассу и сроки цветения | 29, 34, 35 |
| Petunia х hybrid Vilm. | 19 → 12, 17 → 10 16 → 10 |
Отсутствие влияния на высоту растения и биомассу; ускорение цветения | 29 |
| Salvia splendens Selloe ex Schult. | 18 → 12, 17 → 10 16 → 10 |
Отсутствие влияния на высоту растения и сроки цветения | 29 |
| Tagetes patula L. | 19 → 12 | Отсутствие влияния на высоту растения | 29 |
| Verbena elegans Kunth | 19 → 12 | Отсутствие влияния на высоту растения | 29 |
| Другие виды растений | |||
| Arabidopsis thaliana (L.) Heynh. | 16 → 6 | Уменьшение высоты растения | 7 |
| Triticum aestivum L. | 23 → 9 23 → 4 |
Отсутствие влияния на рост листьев и накопление биомассы | 12, 36 |
Цель обзора – обобщение накопленных собственных и литературных экспериментальных данных и наблюдений, касающихся различных эффектов и механизмов формирования ответной реакции растений на ежесуточные кратковременные понижения температуры (дроп-воздействия, от англ. drop – падение).
РЕАКЦИИ РАСТЕНИЙ НА ПРОДОЛЖИТЕЛЬНЫЕ И КРАТКОВРЕМЕННЫЕ НИЗКОТЕМПЕРАТУРНЫЕ ВОЗДЕЙСТВИЯ
Как отмечено во введении, при изучении устойчивости и адаптации растений к холоду чаще всего используются длительные экспозиции в условиях постоянного действия пониженной температуры. Однако следует иметь в виду, что ни в природных условиях, ни при выращивании в теплицах растения не испытывают многосуточного действия одной и той же низкой температуры хотя бы в силу существования хорошо выраженного суточного градиента температуры или, в случае выращивания растений в условиях контролируемого климата, благодаря тому, что нижний температурный предел для дневной температуры устанавливается, как правило, выше ночного. Таким образом, более естественной и часто возникающей является ситуация, когда в суточном цикле периоды с низкой температурой чередуются с более продолжительными теплыми периодами. Специфика дроп-воздействий на растения по сравнению с длительным постоянным действием низких температур определяется, прежде всего, возможностью и способностью растений компенсировать в теплый период в суточном цикле негативные последствия действия низких температур. Растения в это время ликвидируют возникшие у них нарушения и/или повреждения, метаболизируют и/или нейтрализуют токсические вещества, если они появились под влиянием холода [12]. В случае постоянного действия низкой температуры подобное восстановление и детоксикация не происходят или происходят в значительно меньшей степени и поэтому накопление различных структурно-функциональных отклонений и токсичных веществ ведет к нарушению обмена веществ. Кроме того, при длительном постоянном действии низких температур у растений происходит истощение энергетических и пластических ресурсов, необходимых для формирования повышенной устойчивости, тогда как при дроп-воздействиях отношение фотосинтез/дыхание поддерживается на определенном уровне, что позволяет восполнять запасы ассимилятов и энергии [37]. Например, перемещение молодых растений кукурузы (Zea mays L.), выращиваемых при температуре 5°С, в условия субоптимальной температуры 14°С на 1 ч в сутки значительно снижало нарушения водного режима, степень уменьшения фотохимической эффективности ФСII, фотосинтеза и снижения относительной скорости роста, а также сокращало выход электролитов из клеток листьев [38]. Поэтому при ранней высадке рассады в весенний период даже непродолжительные периоды тепла на фоне низких температур в суточном цикле позволяют теплолюбивым растениям компенсировать неблагоприятные последствия холода без ущерба для будущего урожая. Кроме того, в отличие от длительных низкотемпературных воздействий, ингибирующих рост и фотосинтез, дроп-воздействия (при определенных их параметрах − интенсивности, продолжительности, скорости снижения температуры, времени в суточном цикле) могут способствовать более полному проявлению адаптивного потенциала растений, в частности, обеспечивая более высокую скорость фотосинтеза при низкой температуре [12, 13, 37, 39, 40].
Таким образом, специфика дроп-воздействий прежде всего обусловлена тем, что во время них периоды, когда охлаждение запускает в растениях программу адаптационных изменений (сопряженную с торможением роста и развития), чередуются в суточном цикле с гораздо более продолжительными периодами действия оптимальной (или близкой к оптимальной) температуры, при которой происходит возобновление роста и реализация онтогенетической программы. В результате растения становятся более компактными, не снижая при этом скорости фотосинтеза, не замедляя существенно скорости развития и накопления биомассы и приобретая более высокую холодоустойчивость [12–15, 37, 40, 41].
ОСОБЕННОСТИ РЕАКЦИИ РАСТЕНИЙ НА ДРОП-ВОЗДЕЙСТВИЯ В ЗАВИСИМОСТИ ОТ ИХ ПАРАМЕТРОВ
Интенсивность и продолжительность дроп-воздействий. Влияние дроп-воздействий на морфологические и физиолого-биохимические показатели растений зависит от значений, до которых опускается температура. Как правило, морфогенетический эффект, фиксируемый по изменению линейных ростовых показателей (высота растений, длина междоузлий и черешков листьев, размеры листьев), усиливается с понижением температуры, используемой для дроп-воздействий [3, 4, 12, 13, 33, 34, 42, 43]. В то же время реакция растений, оцениваемая по другим показателям (накопление биомассы, содержание хлорофилла, фотосинтетическая активность, относительное содержание воды, относительный выход электролитов, интенсивность перекисного окисления липидов, холодоустойчивость), в большей степени зависит от того, опускается ли температура ниже критического уровня и вызывает ли она развитие стресс-реакции [12, 13, 43]. Наиболее выраженные эффекты дроп-воздействий (торможение роста и накопления биомассы, повышение холодоустойчивости, снижение скорости фотосинтеза и усиление дыхания растений) обычно наблюдаются при использовании низких закаливающих температур, близких к повреждающим, или даже относящихся к повреждающим при их непрерывном и достаточно продолжительном действии на растения. Важно иметь в виду, что при дроп-воздействиях ответная реакция растения в большей степени зависит от абсолютного значения, до которого понижается температура, и продолжительности холодового воздействия, чем от градиента температур, так как именно этим определяется, запускается ли в данном случае комплекс изменений, характерных для стресс-реакции [12].
Предложенные ранее “температурные карты”, указывающие местонахождение на температурной шкале границ разных температурных зон (фоновой зоны, зон холодового закаливания и холодового повреждения), установленные в опытах с постоянным продолжительным (несколько суток) действием низких температур [44–46], не являются абсолютными, а зависят от фазы развития растений, их физиологического состояния и параметров внешней среды. Под влиянием дроп-воздействий эти границы могут существенно смещаться в область более низких температур [37, 43], что необходимо учитывать при выборе значений температуры в экспериментах, направленных на решение тех или иных научных задач. Важно также иметь в виду, что при дроп-воздействиях диапазон, в котором растения способны поддерживать соотношение дыхание/фотосинтез на определенном уровне и накапливать в достаточных количествах фотоассимиляты, оказывается значительно шире, чем в случае постоянного длительного действия на них низких температур [37, 43].
Интересно, что при одинаковой “суммарной дозе” дроп-воздействий более короткие, но интенсивные ежесуточные воздействия (например, 8°С в течение 2 ч) оказывают более сильный эффект, чем более продолжительные умеренные (например, 16°С в течение 8 ч) [33]. Это связано с тем, что снижение температуры до значений ниже критического уровня вызывает дополнительные эффекты, связанные с продолжительным последействием на растения охлаждения, которые не могут быть быстро компенсированы в теплых условиях [43]. Кроме того, в большинстве случаев различия в реакции растений на дроп-воздействия разной продолжительности не определяются обычной зависимостью “доза−эффект”, так как помимо продолжительности в этом случае большую роль играет сам характер температурного воздействия (длительное или кратковременное, постоянное однократное или периодически повторяющееся) [13].
Продолжительность дроп-воздействий приобретает особенно важную роль при снижении температуры до значений, выходящих за пределы оптимума для конкретного процесса. При этом зоны температурного оптимума для разных процессов/показателей (например, рост растений в высоту и время до начала цветения) могут существенно различаться. Последнее определяет эффективность влияния дроп-воздействий на те или иные показатели. Наконец, следует иметь в виду, что эффективность применения дроп-воздействий с заданными параметрами в практических целях (в растениеводстве и цветоводстве) во многом зависит и от видовой принадлежности растений (т.е. от их биологических особенностей). Это предполагает проведение специальных исследований, направленных на выявление оптимальных параметров дроп-воздействий применительно к каждому конкретному объекту.
Помимо ситуации с применением дроп-воздействий с заданными параметрами, которые используются в практике тепличного растениеводства, изучение интенсивности и длительности ежесуточных понижений температуры актуально и для многих теплолюбивых растений, выращиваемых в открытом грунте в умеренной зоне, которые в начале и конце вегетационного сезона могут многократно испытывать кратковременный холодовой стресс. К примеру, это происходит при ранней высадке рассады томатов (Solanum lycopersicum L.) [21], дыни (Cucumis melo L.), арбузов (Citrullus lanatus (Thunb.) Matsum. & Nakai) [9–11] для получения ранней продукции этих культур. Если учесть, что для растений семейства Cucurbitaceae, таких как арбуз и дыня, температурный оптимум роста находится в диапазоне температур от 20 до 32°С с минимумом и максимумом, соответственно, 18 и 35°С, то вполне очевидно, что при ранней высадке растения многократно подвергаются действию субоптимальных температур с последующим их повышением до оптимального уровня. Увеличение же количества часов с температурой ниже оптимальной приводит к снижению урожая [9–11]. Для хлопчатника (Gossypium hirsutum L.) фоновыми являются температуры в диапазоне от 23 до 32°С. Поэтому растения не могут избежать повторяющегося холодового стресса в начале или конце сезона, поскольку в ряде мест традиционного выращивания этой культуры посадка обычно производится в середине мая, а технической зрелости необходимо достичь до первых заморозков, которые наблюдаются уже в конце октября [47]. Очевидно, что сроки посадки зависят от того, какой продолжительности понижения температуры растения способны переносить в начале и конце вегетационного сезона без ущерба для урожая. Этому вопросу посвящен целый ряд исследований, проведенных на разных культурах [9–11, 48, 49], хотя, по мнению агрономов [47], использование результатов, полученных в контролируемых условиях, имеет определенные ограничения для прогнозирования реакции растений на холод в полевых условиях.
Время дроп-воздействий в суточном цикле. Время снижения температуры в суточном цикле может оказывать существенное влияние на реакцию растений на дроп-воздействия. Так, уже в первых работах на лилии (Lilium longiflorum Wiebe), бегонии (Begonia х сhiemaтanta Everett) и томате было показано, что наиболее эффективным для торможения линейного роста растений является снижение температуры в начале светового периода [3]. Ранее считалось, что это связано с тем, что рост стебля в высоту у многих растений наиболее активно происходит именно в ранние утренние часы. Но снижать температуру на 2–3 ч утром в тепличных условиях технически весьма непросто, особенно если утром включаются лампы досветки, излучающие тепло. Необходимо применять сильную принудительную вентиляцию с последующим нагревом, что тоже ведет к дополнительным затратам энергии. Позднее было установлено, что многие растения реагируют на снижение температуры в конце ночи почти также, а некоторые даже сильнее, чем в утренние часы [16, 24–27, 50, 51]. В практическом плане оказалось эффективнее снижать температуру на 5−15°С в конце ночи за счет проветривания. Затем утром, после закрытия форточек и включения ламп досветки, воздух нагревается до нужной температуры. До недавнего времени считалось, что дроп-воздействия не оказывают тормозящего действия на рост стебля, если применяются в другое время суток, кроме как в конце ночи или утром [5, 24, 25, 50]. Однако на разных культурах показано, что дроп-воздействия в любое время суток способны вызывать морфогенетический эффект [17, 22, 52] (рис. 3). При этом наличие или отсутствие света в период понижения температуры оказывает намного более сильное влияние на ответную реакцию растений, чем суточная ритмика чувствительности растений к охлаждению. Добавим к этому, что уже в ранних работах по влиянию переменных температур на рост растений указывалось на то, что реакция растений видоспецифична [31]. Поэтому неудивительно, что данные, полученные на разных видах, могут существенно различаться.
Рис. 3.
Внешний вид контрольных растений Solanum lycopersicum и растений, подвергавшихся дроп-воздействиям (10°С, 2 ч) в начале, середине и конце дня и ночи [22].

Результаты, демонстрирующие хорошо выраженный морфогенетический эффект при дроп-воздействиях в дневное время [3, 16, 17, 22, 35, 52], указывают на то, что рост стебля в длину, как известно, происходит не с постоянной скоростью в суточном цикле, а имеет свои пики, преимущественно в ночное время [32, 53, 54]. Тем не менее, оптимальное время для дроп-воздействий с целью получения компактных растений может не совпадать с периодами наибольшей скорости роста стебля, в частности, у томата [16, 55]. Также в формальном противоречии с мнением о зависимости оптимального времени для дроп-воздействий от суточной ритмики роста стебля или черешков находятся данные о том, что с увеличением длительности дроп-воздействий с 2 до 4 ч [16, 56] или с 1.5 до 3 ч [29] их эффективность снижается или остается прежней, хотя логичней было бы ожидать ее увеличения. Объяснений этому факту пока не найдено, но в этой связи можно отметить интересные данные о стимулировании дроп-воздействиями удлинения стебля растений мари (Chenopodium rubrum L.) в течение 10 ч после окончания низкотемпературного воздействия [53]. У хризантемы (Dendranthema grandiflorum Ramat.) дроп-воздействия не нарушали суточный ритм роста стебля, но значительно изменяли амплитуду колебаний скорости роста в оставшуюся часть суток. Отсюда можно предположить, что дроп-воздействия сами по себе способны изменять ритмику роста растений в суточном цикле. Возможно, с этим связаны нередко возникающие несовпадения получаемых результатов с ожидаемыми.
Скорость снижения температуры. Реакция растений на охлаждение зависит не только от интенсивности и продолжительности низкотемпературного воздействия, но и от скорости снижения температуры [57]. Например, при скорости снижения температуры более чем 1–10°С/мин в растениях наблюдаются реакции, отличные от тех, что происходят при постепенном снижении температуры. Реакции растений на быстрое охлаждение могут не проявляться, если перепад температуры невелик. Экспериментально установлено, что для этого перепад температуры должен быть в пределах от 2 до 10°С [58].
В ряде работ показана способность растений “измерять” скорость снижения температуры. Так, было обнаружено, что диапазон деполяризации мембран корня огурца (Cucumis sativus L.) положительно коррелирует со скоростью охлаждения [59], предполагая роль Са2+ в восприятии растением скорости охлаждения, что и подтвердилось прямым измерением кинетики Са2+ [60]. Скорость изменения температуры может быть определяющей, когда температурные изменения имеют кратковременный характер [58, 59] или когда изучают быстро изменяющиеся показатели [61], например, такие как содержание внутриклеточного кальция. При длительных экспозициях более важным фактором является абсолютное значение температуры [62].
В наших экспериментах с 2-часовыми дроп-воздействиями на растениях огурца резкое снижение температуры (от 20 до 9°С) оказывало более сильное негативное воздействие на состояние фотосинтетического аппарата растений и накопление биомассы по сравнению с постепенным (0.4°С/мин) [63]. В то же время линейный рост и развитие растений, а также холодоустойчивость листьев изменялись под влиянием дроп-воздействий независимо от скорости снижения температуры. Следовательно, в лабораторных экспериментах, когда при изучении влияния температурного фактора на растения происходит резкая смена температуры (в результате быстрой перестановки растений из одних температурных условий в другие), необходимо учитывать, что при кратковременном низкотемпературном воздействии скорость снижения температуры может быть более сильным, определяющим реакцию растения фактором по сравнению с ее абсолютным значением, которого она достигает в результате снижения.
ВЛИЯНИЕ ФАКТОРОВ ВНЕШНЕЙ СРЕДЫ НА РЕАКЦИЮ РАСТЕНИЙ НА ДРОП-ВОЗДЕЙСТВИЯ
Свет. Свет способен оказывать значительное модифицирующее действие на реакцию растений на дроп-воздействия. Прежде всего, эффекты дроп-воздействий на растения различаются в зависимости от того, осуществляются они в присутствии света или в темноте. Дроп-воздействия на свету оказывают больший морфогенетический эффект [17, 64, 65]. В определенной степени это связано со снижением активности гиббереллинов в результате быстрого увеличения экспрессии одного из генов (PsGA2ox2), что приводит к деактивации гиббереллинов [20]. Экспрессия этого гена происходит лишь при действии низкой температуры на свету. Однако, надо отметить, что механизм, связанный с активностью гиббереллинов, участвует лишь в изменениях морфологии растений и не влияет на их фотосинтетическую и дыхательную способность [66]. В то же время свет может усиливать негативное влияние холода на растения, вызывая в условиях низких температур более значимые функциональные нарушения в фотосинтетическом аппарате (ФСА) [17, 64, 65], которые могут приводить к снижению скорости накопления биомассы. Даже относительно слабый свет при холодовом воздействии может приводить к поглощению растениями неиспользуемой в фотохимических реакциях световой энергии и вызывать фотоингибирование, проявляющееся в снижении скорости фотосинтеза, фотоокислении пигментов и разрушении структуры хлоропластов.
Помимо этого, реакция растений на охлаждение может зависеть и от световых условий, предшествовавших действию холода. Более высокий уровень освещенности, используемый при выращивании растений, способствует поддержанию растениями, подвергающихся дроп-воздействиям в темноте, высоких скоростей фотосинтеза (на уровне контрольных растений) при оптимальной температуре и повышению устойчивости фотосинтеза к низкой температуре [65]. При низкой освещенности влияние дроп-воздействий может быть менее выраженным из-за дефицита ассимилятов, что само по себе ограничивает рост растений.
Результаты экспериментов, проведенных с более чем 20 различными видами и сортами растений, отчетливо указывают на то, что эффективность дроп-воздействий может также зависеть от фотопериода [42, 67]. Наибольший эффект дроп-воздействий на высоту растений проявляется в условиях короткого фотопериода. С увеличением фотопериода эффект ослабевает или отсутствует. Для проявления эффекта дроп-воздействий в условиях длинных фотопериодов, как правило, требуются более продолжительные низкотемпературные воздействия. Вероятно, это связано с усилением синтеза гиббереллинов при увеличении фотопериода, что свойственно как коротко- [68], так и длиннодневным [69] растениям.
Изучение реакции растений на дроп-воздействия при разных фотопериодах показало, что в условиях круглосуточного освещения различия в реакции ФСА растений на постоянную температуру и температурный режим, включающий дроп-воздействия, выражены в наибольшей степени. Опыты в условиях круглосуточного освещения показали, что дроп-воздействия с использованием закаливающих значений температуры препятствуют световому повреждению листьев (развитию хлороза) и стабилизируют работу ФСА у чувствительных к круглосуточному освещению растений [70, 71]. Однако, следует иметь ввиду, что дроп-воздействия не обладают выраженным последействием и нивелируют или ослабляют повреждающий эффект круглосуточного освещения только в период их применения [72].
Вода. И относительная влажность воздуха, и обеспеченность водой оказывают сильное количественное и качественное влияние на реакцию растений на дроп-воздействия [73, 74]. При высокой относительной влажности воздуха дроп-воздействия вызывают значительный морфогенетический эффект, не снижая или даже увеличивая биомассу растения, что приводит к увеличению компактности растений. В условиях низкой относительной влажности воздуха и дефицита воды эффекты дроп-воздействий на компактность растений нивелируются вследствие сильного торможения линейного роста и накопления биомассы растений в результате недостатка влаги.
При совместном длительном действии низких положительных температур и водного стресса негативные эффекты усиливаются, что приводит к снижению продуктивности растений. Совместное действие “периодической засухи” (как особого агроприема) и дроп-воздействий при высокой относительной влажности воздуха приводит к увеличению компактности растений, а также их устойчивости к низкой температуре и водному стрессу, индуцированному низкой температурой [73, 74]. Хотя морфогенетический эффект таких альтернативных применению ретардантов агроприемов как дроп-воздействия и “периодическая засуха” сопоставим в плане их влияния на линейные размеры растений, увеличение компактности растений происходит только под влиянием дроп-воздействий. При “периодической засухе” растения становятся меньше, но не компактнее, из-за значительного снижения биомассы. Это важно иметь в виду, так как современное растениеводство, ориентированное на снижение затрат на единицу получаемой продукции, требует поиска относительно недорогих и безопасных для окружающей среды способов управления высотой и компактностью растений из-за строгих маркетинговых требований к качеству рассады овощных культур и при выращивании декоративных растений [75].
РЕАКЦИИ НА ДРОП-ВОЗДЕЙСТВИЯ ПРЕДСТАВИТЕЛЕЙ РАЗНЫХ ПО ОТНОШЕНИЮ К ТЕМПЕРАТУРЕ И СВЕТУ ГРУПП РАСТЕНИЙ
Теплолюбивые и холодостойкие растения. Рассмотрению ответных реакций теплолюбивых растений на дроп-воздействия посвящено немало экспериментальных работ и обзоров [3, 4, 15, 52], но данных о реакции холодостойких растений немного [12, 76, 77]. Несмотря на значительные качественные и количественные различия в реакциях холодостойких и теплолюбивых растений на постоянное продолжительное действие и последействие пониженных температур, их реакции на дроп-воздействия в целом качественно схожи [12, 36]. И у теплолюбивых, и у холодостойких видов дроп-воздействия вызывают торможение линейного роста органов растений, а также целый спектр физиологических изменений, способствующих более полному проявлению адаптивного потенциала, в частности, обеспечивая более высокую скорость фотосинтеза при низкой температуре по сравнению с контрольными растениями (не подвергавшимися низкотемпературным воздействиям), а также рост их холодоустойчивости.
Растения разных фотопериодических групп. На основании относительно немногочисленных экспериментов, проведенных в разных условиях, более 20 лет назад был сделан вывод, что длиннодневные растения менее отзывчивы на дроп-воздействия, чем короткодневные [42]. Однако, наш анализ результатов опытов, проведенных с растениями разных фотопериодических групп (короткодневных, нейтральных и длиннодневных) в условиях разных фотопериодов [15, 33, 42, 31, 67, 78 ], позволяет заключить, что реакция растений на дроп-воздействия зависит не от их принадлежности к определенной фотопериодической группе, а прежде всего определяется условиями проведения опыта, а именно продолжительностью фотопериода. Растения реагируют на дроп-воздействия сильнее в условиях коротких фотопериодов, а с увеличением фотопериода, как правило, требуются более длительные низкотемпературные воздействия для торможения роста. Кроме того, изучение динамики роста стебля в дневное и ночное время в условиях короткого и длинного дня показало, что она может значительно различаться у разных видов растений, но это не связано с их принадлежностью к той или иной фотопериодической группе [54].
Светолюбивые и теневыносливые растения. Поскольку низкие температуры снижают скорость темновых реакций фотосинтеза, и на свету в условиях холода нарушается баланс между поглощением энергии и ее использованием, то можно предположить, что теневыносливые растения, имеющие бóльший размер светособирающего комплекса и меньшие скорости транспорта электронов, могут в бóльшей степени, чем светолюбивые, быть подвержены фотоингибированию, вызванному понижением температуры в присутствии света. Результаты наших опытов со светолюбивыми и теневыносливыми гибридами огурца показали, что требовательность растений к свету может в определенных условиях влиять на реакцию растений на дроп-воздействия [64, 65]. В частности, у разных гибридов огурца в ответной реакции на дроп-воздействия наряду с общими чертами наблюдалась и определенная специфичность, связанная с экотипическими особенностями того или иного гибрида. Наибольшие различия между светолюбивыми и теневыносливыми гибридами проявились в их способности к температурной адаптации. Отдаленный во времени эффект дроп-воздействий на свету в виде снижения ранней урожайности на 50−65% проявлялся только у теневыносливых гибридов огурца.
В целом можно заключить, что в реакциях растений на дроп-воздействия значительную роль играет видоспецифичность, не связанная напрямую с их принадлежностью к той или иной группе растений − теплолюбивых или холодостойких, светолюбивых или теневыносливых, короткодневных, длиннодневных или фотопериодически нейтральных.
ФИЗИОЛОГО-БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ РЕАКЦИИ РАСТЕНИЙ НА ДРОП-ВОЗДЕЙСТВИЯ
Линейный рост и морфогенез. В основе торможения роста растений под влиянием дроп-воздействий, который зафиксирован в отношении многих видов, прежде всего, лежат изменения в содержании активных гиббереллинов [79–82]. При этом механизмы адаптации к низким температурам на свету и в темноте существенным образом различаются. На свету в результате усиления экспрессии гена PsGA2ox2 происходит инактивация активного гиббереллина ГА1, что приводит к торможению линейного роста. В то же время отсутствие влияния дроп-воздействий в темноте на содержание ГА1 и экспрессию PsGA2ox2 на фоне снижения скорости роста стебля указывает на участие иных факторов в реакции растений на охлаждение. Известно, что температура влияет и на активность других гормонов (ауксинов, цитокининов, АБК, брассиностероидов и т.д.), которые также участвуют в регуляции роста растений, однако их роль в реакциях растений на дроп-воздействия неизвестна [75].
Наряду с фитогормонами определенную роль в реакции растений на дроп-воздействия может играть фитохром В [15, 56, 82, 83]. Возможно, он регулирует чувствительность тканей стебля к гиббереллинам и именно благодаря фитохрому В растения имеют высокое содержание гиббереллинов в условиях длинных фотопериодов [75].
Влияние дроп-воздействий на водообмен следует также рассматривать как одну из важных причин торможения роста растений [34, 74, 75]. Ограничения в поглощении воды корнями при снижении температуры и снижение скорости транспирации (хотя и не полное ее прекращение) во время дроп-воздействий могут приводить к водному стрессу, в результате чего происходит торможение линейного роста, т.к. пониженное тургорное давление непосредственно приводит к уменьшению растяжения клеток.
Необходимым условием для роста растений является наличие углеводов, а нехватка фотоассимилятов, служащих строительным материалом и субстратами для дыхания, может снижать скорость роста и тем самым нивелировать эффект дроп-воздействий, что показано в опытах с использованием относительно низкой освещенности [65]. Добавим, что в определенных случаях торможение линейного роста под влиянием дроп-воздействий может не проявляться из-за того, что они осуществляются в конце ночи или начале дня, когда возникает наибольший дефицит ассимилятов, необходимых для роста.
Таким образом, физиологические механизмы, обуславливающие морфогенетический эффект дроп-воздействий, а именно их влияние на рост растений, могут включать изменение гормонального и водного статуса растений, а также участие фитохромной системы. При этом эффекты дроп-воздействий могут быть модифицированы действием светового и водного факторов. В то же время морфогенетический эффект дроп-воздействий может снижаться или не проявляться в условиях, способствующих очень быстрому росту, а именно в условиях длинных фотопериодов (из-за высокого содержания активных гиббереллинов) и низкого соотношения красного (К) и дальнего красного (ДК) света− К/ДК (при высокой плотности посадки, использовании ламп с низким отношением К/ДК, затенении). Также эффект дроп-воздействий может нивелироваться при сильно заторможенном росте в условиях низкой освещенности растений (из-за недостатка фотоассимилятов), низкой относительной влажности воздуха и при высоком соотношении К/ДК (при использовании специальных фильтров или ламп с высоким соотношением К/ДК). Эффективность дроп-воздействий может повышаться при их сочетании с так называемой “периодической засухой” в условиях высокой относительной влажности воздуха [74], которую применяют в качестве агроприема для управления ростом растений в тепличных хозяйствах.
Вегетативное и репродуктивное развитие. Главными факторами, определяющими скорость развития растений, как известно, являются не температура дня или ночи, а среднесуточная температура [84], и сроки цветения определяются суммой эффективных температур. Поскольку дроп-воздействия в силу краткосрочности приводят лишь к незначительному их понижению, то дроп-воздействия, которые применяются в практических целях, как правило, не оказывают значимого влияния на скорость вегетативного и генеративного развития [12, 13, 26, 33, 50]. Хотя в некоторых случаях отмечено увеличение ранней урожайности томатов и сладкого перца (Capsicum annuum L.), когда растения подвергались дроп-воздействиям во время пререпродуктивного периода [71]. Однако в ситуации, когда дроп-воздействия характеризуются большой интенсивностью и/или продолжительностью, наблюдается снижение скорости появления листьев и/или задержка начала цветения и плодоношения [23, 33, 42, 64], но они несопоставимы по своим последствиям для процесса развития, которое тормозится вплоть до полной остановки при постоянном действии на растения низкой температуры.
Фотосинтез, дыхание и продуктивность. Фотосинтез является одним из наиболее чувствительных к холоду физиолого-биохимических процессов, поэтому адаптация растений к низким температурам тесно связана с адаптацией ФСА, которая проявляется в повышении термоустойчивости фотосинтеза. На ряде видов растений показано, что дроп-воздействия, в отличие от постоянного действия низкой температуры, не снижают скорость фотосинтеза в оптимальных условиях, а в условиях низкой температуры вызывают ее увеличение по сравнению с контролем [12, 37, 40]. Способность растений, подвергающихся дроп-воздействиям, поддерживать устьица в открытом состоянии в условиях низких температур способствует обеспечению высокого уровня фотосинтеза [39]. Показано также, что под влиянием дроп-воздействий у некоторых видов растений возрастает фотосинтетическая эффективность использования воды, что отражает увеличение их устойчивости к дефициту воды [18]. В то же время дроп-воздействия, особенно на свету, вызывают снижение эффективности использования световой энергии в фотосинтетических реакциях (видимого квантового выхода фотосинтеза) в результате инициации ряда защитно-приспособительных реакций растений на низкую температуру, позволяющих им избежать повреждения ФСА [85].
Анализ изменения отношения темнового дыхания (Rd) листьев к фотосинтезу (Ag) − R/A (показатель сбалансированности основных физиологических процессов) у растений, подвергнутых дроп-воздействиям, показал, что оно увеличивалось, при этом величина изменения возрастала с усилением интенсивности дроп-воздействий, то есть со снижением значения действующей на растения температуры [37]. Увеличение данного показателя под влиянием дроп-воздействий на уровне листа было обусловлено в большей степени повышением Rd и в меньшей – снижением Ag. Важно, что в случае дроп-воздействий абсолютные значения температур, приводящие к увеличению соотношения R/A, оказываются значительно ниже тех, которые при постоянном ее действии приводят к снижению скорости фотосинтеза и возрастанию скорости дыхания, а следовательно, к росту R/A. Например, при постоянном действии температуры 12°С в течение 6 суток величина R/A составляет 2.08, а при дроп-воздействиях с использованием той же температуры − всего 0.05. Это свидетельствует о том, что при дроп-воздействиях температурный диапазон, в пределах которого растения способны поддерживать на определенном уровне соотношение R/A и, следовательно, скорость накопления сухой биомассы, значительно шире, чем при постоянном длительном действии низких температур. В большинстве работ, где применялись дроп-воздействия умеренной интенсивности (снижение до 8–12°С) и небольшой продолжительности (2–3 ч), растения накапливали биомассу, сопоставимую с растениями, не подвергавшимися охлаждению, или даже большую [12, 14, 27, 29, 33, 51, 73].
Следует отметить, что стимуляция дыхания под влиянием дроп-воздействий, проявляющаяся у теплолюбивых растений огурца при низких температурах, обусловлена в основном повышением активности цитохромного, а не альтернативного пути дыхания [86, 87], а у холодостойкой пшеницы (Triticum aestivum L.) дроп-воздействия, в отличие от постоянного действия низкой температуры, не вызывают явных изменений в дыхательном метаболизме [87].
Холодоустойчивость. Дроп-воздействия способны вызывать повышение холодоустойчивости теплолюбивых растений, в основе которого лежат следующие события: изменения в экспрессии генов холодового ответа и синтезе белков [88, 89]; изменение липидного состава мембран, а именно увеличение ненасыщенности жирных кислот [90, 91]; адаптивные изменения в ФСА, приводящие к поддержанию фотосинтеза при низкой температуре и обеспечению растений фотоассимилятами [12, 37–39]. Участие и роль в механизмах повышения холодоустойчивости растений, наблюдаемого под влиянием дроп-воздействий, физиолого-биохимических изменений, связанных с накоплением сахаров и других осмотически активных веществ, а также активацией антиоксидантной системы требуют дополнительного изучения.
Вопрос о влиянии дроп-воздействий на экспрессию генов у растений почти не изучен. Известно лишь, что у растений огурца, подвергавшихся дроп-воздействиям и постоянному действию низкой (12°С) температуры в течение 6 суток, существенные различия в экспрессии генов между этими вариантами опыта отсутствовали, о чем свидетельствует показатель отношения индуцированных фрагментов к общему числу амплифицированных фрагментов кДНК [15]. Такое почти полное совпадение паттернов фрагментов кДНК в обоих вариантах низкотемпературного воздействия может говорить не только о сходстве в регуляции экспрессии генов, но и об экспрессии сходного набора генов. Предполагается, что более высокий уровень холодоустойчивости в варианте с дроп-воздействиями по сравнению с постоянным действием низкой температуры обеспечивается не за счет включения определенных генов и индуцированного синтеза белков de novo, а благодаря изменению уровня экспрессии уже функционирующих генов и/или участию иных механизмов, не связанных непосредственно с геномом [15]. Изучение экспрессии одного из СОR-генов у растений картофеля (Solanum tuberosum L.), подвергавшихся дроп-воздействиям или постоянному действию низкой закаливающей температуры (5°С) в течение 6 суток, показало, что низкотемпературные воздействия обоих типов вызывают экспрессию гена ci7 [89]. Причем, если для достижения максимального уровня экспрессии этого гена при постоянном действии низкой температуры требовалось 72 ч, то при дроп-воздействиях подобный эффект достигался уже при 12 ч суммарного действия низкой температуры (6 суток по 2 ч) [15].
В то же время изучение особенностей реакции растений огурца и пшеницы, у которых был подавлен биосинтез белков на 80S цитоплазматических рибосомах (с помощью циклогексимида) или на 70S рибосомах митохондрий и хлоропластов (с помощью хлорамфеникола), на длительное и кратковременное действие пониженных температур позволило установить зависимость процесса повышения холодоустойчивости растений при дроп-воздействиях от биосинтеза белков [36], подобно тому, как это установлено ранее в отношении реакции растений на длительное (постоянное) действие холода [44]. Исходя из этих данных, вопрос о роли генетических факторов и индуцированного синтеза белков остается открытым и требует дальнейших исследований.
ЗАКЛЮЧЕНИЕ
Рост большинства видов растений протекает в условиях частых и весьма значительных по амплитуде колебаний температуры – по годам, сезонных и суточных. Одной из главных и усиливающихся тенденций изменения климата в последние годы является увеличение его нестабильности. Поэтому вполне естественно, что в процессе длительной эволюции физиологические процессы, определяющие рост и развитие растений, оказались хорошо адаптированными к широкому спектру внешних факторов и именно это позволяет растениям успешно осуществлять свою жизнедеятельность в широком диапазоне внешних условий, включая температурные. Важно, что наличие у растений значительного адаптивного потенциала открывает большие возможности для ведения селекционно-генетической работы, а также решения практических задач, связанных с выращиванием растений в условиях контролируемого климата. Это же открывает определенные возможности для поиска и апробации новых стратегий сохранения энергии в теплицах, экономическая эффективность использования которых в значительной степени зависит от энергозатрат. Так, например, в последнее время получила признание среди специалистов концепция температурного интегрирования (“temperature integration”, TI) [84, 92]. Она предполагает, что температура может колебаться в суточном цикле между нижним значением, соответствующим “базовой” температуре (нижнему температурному пределу вегетации, при котором развитие прекращается), и верхним, совпадающим со значением оптимальной температуры (при которой скорость развития максимальна). При этом действие низких температур компенсируется последующим действием высоких. Концепция TI основана на способности растений в первую очередь реагировать на среднесуточную температуру, а не на ее изменения в суточном цикле [93, 94]. Показано, что с помощью TI можно достичь не менее 5–15% экономии энергии без ущерба для роста и развития растений [95]. Ее применение совместно с динамическим контролем климата (“dynamic climate control”), заключающимся в регуляции температуры, освещенности и концентрации CO2 на основе изменений показателей фотосинтетической активности, позволяет достичь еще больших результатов [96]. Применение дроп-воздействий, которым посвящена данная статья, органично вписывается в концепцию TI, поскольку подразумевает, что температура может снижаться в ночное время, но не ниже значения “базовой”, и что в теплое время суток возможные негативные последствия охлаждения растений будут компенсированы. У холодостойких видов значения “базовой” температуры варьируют от 0 до 4°С, у теплолюбивых – от 8 до 12°С, и для каждого культивируемого вида они точно определены [97].
Поскольку реакция на кратковременные ежесуточно повторяющиеся понижения температуры является выработанным в процессе эволюции адаптивным ответом растений, испытывающих на протяжении многих поколений не только сезонные и внутрисезонные флуктуации температуры, но и ее колебания в суточном цикле, то так же, как в случае с реакцией на действие других стресс-факторов, адаптация к дроп-воздействиям включает в себя достаточно широкий комплекс физиолого-биохимических изменений. Их набор, соотношение и удельный вес зависят от параметров дроп-воздействий, видовых особенностей растений, их физиологического состояния и сопутствующих условий внешний среды. Благодаря этим изменениям растения оказываются способными не только переносить непродолжительные понижения температуры в суточном цикле, но и поддерживать при этом нормальную жизнедеятельность, рост и развитие. На практике применение дроп-воздействий в качестве агроприема позволяет управлять ростом растений без использования ретардантов (или существенно сокращая их количество) и с меньшими затратами энергии, что ведет не только к повышению эффективности сельскохозяйственного производства, но и увеличению доли экологически чистой продукции, что отвечает на запрос со стороны потребителя.
Что касается вопросов и научных задач, требующих изучения в ближайшие годы, то наиболее важным нам представляется выявление различий между температурными воздействиями разного типа на уровне генома, транскриптома и протеома. Такого рода данные крайне интересны и важны, поэтому развитие и применение современных методов исследований позволяет надеяться, что они появятся уже в ближайшие годы. Следует также продолжить изучение способов (каналов) восприятия растением отдельных параметров низкотемпературных воздействий. Несмотря на значительные успехи в изучении путей передачи сигнала в растениях, индуцированного понижением температуры [98, 99], неясным остается вопрос относительно тонких механизмов восприятия низкотемпературного сигнала. Не исключено, что в зависимости от скорости снижения температуры, интенсивности и продолжительности ее действия роль термосенсоров могут выполнять разные структуры или вещества, что позволяет растениям реагировать на охлаждение более гибко и адекватно. Круг потенциальных кандидатов на роль сенсоров низкой температуры включает в себя как отдельные внутриклеточные структуры, так и вещества – мембраны, элементы цитоскелета, хроматин, фитохромы, ДНК, РНК, определенные белки и сахара [100]. Не до конца выяснены механизмы морфогенетического эффекта дроп-воздействий и повышения холодоустойчивости растений под их влиянием. Практически вообще не изучалась ответная реакция на дроп-воздействия у дикорастущих растений. Ответы на эти и некоторые другие вопросы помогут не только лучше понять природу данного феномена, но и оценить его фундаментальную роль в жизнедеятельности активно вегетирующих растений, открывая вместе с тем новые возможности для управления их ростом в условиях контролируемого климата.
Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований в рамках научного проекта № 1914-50502 и государственного задания КарНЦ РАН (0218-2019-0074).
Авторы заявляют об отсутствии конфликта интересов. Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов исследований.
Список литературы
Levitt J. Responses of plants to environmental stresses: chilling, freezing and high temperature stresses. New York: Academic Press, 1980. 497 p.
Theocharis A., Clement C., Barka E.A. Physiological and molecular changes in plants grown at low temperatures // Planta. 2012. V. 235. P. 1091. https://doi.org/10.1007/s00425-012-1641-y
Myster J., Moe R. Effect of diurnal temperature alternations on plant morphology in some greenhouse crops: a mini review // Sci. Hortic. 1995. V. 62. P. 205. https://doi.org/10.1016/0304-4238(95)00783-P
Moe R., Heins R.D. Thermo- and photomorphogenesis in plants // Advances in floriculture research. Report no 6/2000. Agricultural University of Norway / Ed. Strømme E. Oslo: Spekter, 2000. P.52.
Runkle E. Controlling height with temperature drops // Greenhouse Product News. 2009. V. 4. P. 50.
Moe R. Morphogenetic effects of temperature and control of plant height by day/night temperature alternations // The Scientific Basis of Poinsettia Production. Agricultural University of Norway. Advisory Service / Ed. Strømme E. Ås, 1994. P. 65.
Stavang J.A., Hansen M., Olsen J.E. Short term temperature drops do not enhance cold tolerance // Plant Growth Regul. 2008. V. 55. P. 199. https://doi.org/10.1007/s10725-008-9274-7
Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Реакция теплолюбивых и холодостойких растений на ежесуточные кратковременные понижения температуры // Материалы Всероссийской научной конференции, посвященной 125-летию Института физиологии растений им. К.А. Тимирязева РАН “Фундаментальные и прикладные проблемы современной экспериментальной биологии растений”. Москва: ИФР РАН, 2015б. С. 737.
Korkmaz A., Dufault R.J. Developmental consequences of cold temperature stress at transplanting on seedling and field growth and yield. I. Watermelon // J. Amer. Soc. Hort. Sci. 2001a. V. 126. P. 404https://doi.org/10.21273/JASHS.126.4.404
Korkmaz A., Dufault R.J. Developmental consequences of cold temperature stress at transplanting on seedling and field growth and yield. II. Muskmelon // J. Amer. Soc. Hort. Sci. 2001b. V. 126. P. 410. https://doi.org/10.21273/JASHS.126.4.410
Korkmaz A., Dufault R.J. Differential cold stress duration and frequency treatment effects on muskmelon seedling and field growth and yield // Europ. J. Hort. Sci. 2004. V. 69. P. 12.
Шибаева Т.Г., Икконен Е.Н., Шерудило Е.Г., Титов А.Ф. Влияние ежесуточных кратковременных понижений температуры на теплолюбивые и холодостойкие растения // Физиология растений. 2019. Т. 66. С. 279. https://doi.org/10.1134/S0015330319040122
Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Реакция растений Cucumis sativus L. на длительное постоянное и кратковременные ежесуточные воздействия низких температур // Физиология растений. 2018. Т. 65. С. 68.https://doi.org/10.7868/S0015330318020069
Марковская Е.Ф., Сысоева М.И., Харькина Т.Г., Шерудило Е.Г. Влияние кратковременного снижения ночной температуры на рост и холодостойкость растений огурца // Физиология растений. 2000. Т. 47. С. 511.
Марковская Е.Ф., Сысоева М.И., Шерудило Е.Г. Кратковременная гипотермия и растение. Петрозаводск: Карельский научный центр РАН, 2013. 194 с.
Grimstad S.O. Low-temperature pulse affects growth and development of young cucumber and tomato plants // J. Hortic. Sci. 1995. V. 70. P. 75. https://doi.org/10.1080/14620316.1995.11515275
Shibaeva T.G., Sherudilo E.N., Ikkonen E.N., Titov A.F. Responses of young cucumber plants to a diurnal temperature drop at different times of day and night // Act-a Agric. Slov. 2018. V. 111. P. 567. https://doi.org/10.14720/aas.2018.111.3.05
Ikkonen E.N., Shibaeva T.G., Titov A.F. Water use efficiency in Cucumis sativus L. in response to daily short-term temperature drop // J. Agricult. Sci. 2015. V. 7. P. 208. https://doi.org/10.5539/jas.v7n11p208
Sysoeva M.I., Markovskaya E.F., Kharkina T.G., Sherudilo E.G. Temperature drop, dry matter accumulation and cold resistance of young cucumber plants // Plant Growth Regul. 1999. V. 28. P. 89. https://doi.org/10.1023/A:1006243230411
Stavang J.A., Junttila O., Moe R., Olsen J. Differential temperature regulation of GA metabolism in light and darkness in pea // J. Exp. Bot. 2007. V. 58. P. 3061. https://doi.org/10.1093/jxb/erm163
Dufault R.J., Melton R.R. Cyclic cold stress before transplanting influence tomato seedling growth, but not fruit earliness, fresh-market yield, or quality // J. Amer. Soc. Hort. Sci. 1990. V. 115. P. 559. https://doi.org/10.21273/JASHS.115.4.559
Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Реакции растений на кратковременные ежесуточные понижения температуры в разное время дня и ночи // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2020. № 3. С. 60. https://doi.org/10.17076/eb1088
Grimstad S.O. The effect of a daily low temperature pulse on growth and development of greenhouse cucumber and tomato plants during propagation // Sci. Hort. 1993. V. 53. P. 53. https://doi.org/10.1016/0304-4238(93)90137-F
Moe R., Mortensen L.M. Thermomorphogenesis in pot plants // Acta Hort. 1992. V. 305. P. 19. https://doi.org/10.17660/ActaHortic.1992.305.2
Grindal G., Moe R. Effects of temperature-drop and a short dark interruption on stem elongation and flowering in Begonia x hiemalis Fotsch. // Sci. Hortic. 1994. V. 57. P. 123. https://doi.org/10.1016/0304-4238(94)90040-X
Grindal G., Moe R. Growth rhythm and temperature DROP // Acta Hort. 1995. V. 378. P. 47. https://doi.org/10.17660/ActaHortic.1995.378.6
Bakken A.K., Moe R. Height and quality control in Christmas begonia by growth-retarding temperature regimes // Acta Agric. Scand. Sect. B, Soil and Plant Sci. 1995. V. 45. P 283. https://doi.org/10.1080/09064719509413117
Vogelezang J.V.M. The timing of low temperature treatments on stem elongation as affected by lighting strategies // Acta Hort. 1997. V. 435. P. 47. https://doi.org/10.17660/ActaHortic.1997.435.3
Mortensen L.M., Moe R. Effects of various day and night temperature treatments on the morphogenesis and growth of some greenhouse and bedding plant species // Acta Hort. 1992. V. 327. P. 77. https://doi.org/10.17660/ActaHortic.1992.327.9
Ihlebekk H., Eilertsen S., Junttila O., Grindal G., Moe R. Control of plant height in Campanula isophylla by temperature alternations; involvement of GAS // Acta Hort. 1995. V. 394. P. 347. https://doi.org/10.17660/ActaHortic.1995.394.38
Cuijpers L.H.M., Vogelezang J.V.M. DIF and temperature drop for short-day pot plants // Acta Hort. 1992. V. 327. P. 25. https://doi.org/10.17660/ActaHortic.1992.327.3
Tutty J.R., Hicklenton P.R., Kristie D.N., McRae K.B. The influence of photoperiod and temperature on the kinetics of stem elongation in Dendranthema grandiflorum // J. Amer. Soc. Hortic. Sci. 1994. V. 119. P. 138. https://doi.org/10.21273/JASHS.119.2.138
Ueber E., Hendriks L. Effects of intensity, duration and the time of a temperature drop on growth and flowering of Euphorbia pulcherrima Willd. ex. Klotzsch // Acta Hortic. 1992. V. 327. P. 33. https://doi.org/10.17660/ActaHortic.1992.327.4
Ueber E., Hendriks L. Effect of a short duration temperature drop on water status and elongation growth of ornamental plants // Acta Hortic. 1997. V. 435. P. 25. https://doi.org/10.17660/ActaHortic.1997.435.1
Ueber E., Hendriks L. Intensity effects of a temperature drop on pelargoniums // Acta Hort. 1995. V. 378. P. 34. https://doi.org/10.17660/ActaHortic.1995.378.3
Шерудило Е.Г., Шибаева Т.Г., Икконен Е.Н., Титов А.Ф. Сравнительное изучение эффектов длительного постоянного и кратковременных ежесуточных воздействий низких температур на основные физиологические процессы растений в условиях подавленного биосинтеза белков // Журнал общей биологии. 2019. Т. 80. С. 57. https://doi.org/10.1134/S0044459619010056
Икконен Е.Н., Шибаева Т.Г., Титов А.Ф. Влияние кратковременных ежесуточных понижений температуры на соотношение дыхания и фотосинтеза у теплолюбивых растений // Физиология растений. 2018. Т. 65. С. 63. https://doi.org/10.7868/S0015330318010050
Koscielniak J., Biesaga-Koscielniak J. The effect of short warm breaks during chilling on water status, intensity of photosynthesis of maize seedlings and final grain yield // J. Agron. Crop Sci. 2000. V. 184. P. 1.https://doi.org/10.1046/j.1439-037x.2000.00365.x
Икконен Е.Н., Шибаева Т.Г., Сысоева М.И., Шерудило Е.Г. Устьичная проводимость Cucumis sativus L. при длительном и кратковременном действии низких температур // Физиология растений. 2012. Т. 59. С. 716.
Икконен Е.Н., Шибаева Т.Г., Титов А.Ф. Реакция фотосинтетического аппарата листа у Cucumis sativus L. на кратковременное ежесуточное понижение температуры // Физиология растений. 2015. Т. 62. С. 528. https://doi.org/10.7868/S0015330315040090
Шерудило Е.Г., Шибаева Т.Г. Влияние кратковременных ежесуточных понижений температуры на холодоустойчивость листьев разного возраста // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2018. №6. С. 115. https://doi.org/10.17076/eb821
Moe R., Willumsen K., Ihlebekk I.H., Stupa A.I., Glomsrud N.M., Mortensen L.M. DIF and temperature drop responses in SDP and LDP, a comparison // Acta Hortic. 1995. V. 378. P. 27. https://doi.org/10.17660/ActaHortic.1995.378.2
Шибаева Т.Г., Икконен Е.Н., Шерудило Е.Г., Титов А.Ф. Особенности реакции растений на ежесуточные понижения температуры в зависимости от их интенсивности и продолжительности // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2018. № 12. С. 20. https://doi.org/10.17076/eb884
Дроздов С.Н., Курец В.К., Титов А.Ф. Терморезистентность активно вегетирующих растений. Л.: Наука, 1984. 168 с.
Drozdov S.N., Titov A.F., Talanova V.V., Kritenko S.P., Sherudilo E.G., Akimova T.V. The effect of temperature on cold and heat resistance of growing plants. I. Chilling-sensitive species // J. Exp. Bot. 1984. V. 35. P. 1595. https://doi.org/10.1093/jxb/35.11.1595
Титов А.Ф., Дроздов С.Н., Акимова Т.В., Таланова В.В. Исследование реакции растений сои на действие температуры. Границы температурных зон // Физиология растений. 1987. Т. 34. С. 350.
Holaday A.S., Mahan J.R., Payton P. Molecular biology and physiology: Effects of chilling temperatures on photosynthesis // J. Cotton Sci. 2016. V. 20. P. 220.
Bradow J.M. Cotton cultivar responses to suboptimal postemergent temperatures // Crop Sci. 1991. V. 31. P. 1595. https://doi.org/10.2135/cropsci1991.0011183X003100 060043x
Sofalian O., Azimy S., Jahanbakhsh S., Khomari S., Dezhsetan S. Evaluating genetic diversity of chilling stress in cotton genotypes // Plant Breed. Seed Sci. 2013. V. 68. P. 77. https://doi.org/10.2478/v10129-011-0082-2
Moe R., Fjeld T., Mortensen L. Stem elongation and keeping quality in poinsettia (Euphorbia pulcherrima Willd.) as affected by temperature and supplementary lighting // Sci. Hort. 1992. V. 50. P. 127. https://doi.org/10.1016/S0304-4238(05)80015-9
Moe R., Glomsrud N., Bratberg I., Valso S. Control of height in poinsettia by temperature drop and graphical tracking // Acta Hort. 1992. V. 327. P. 41. https://doi.org/10/17660/ActaHortic.1992.327.5
Erwin J.E., Heins R.D. Thermomorphogenetic responses in stem and leaf development // Hort. Sci. 1995. V. 30. P. 940. https://doi.org/10.21273/HORTSCI.30.5.940
Lecharny A., Schwall M., Wagner E. Stem extension rate in light-grown plants // Plant Physiol. 1985. V. 79. P. 625. https://doi.org/10.1104/pp.79.3.625
Bertram L., Karlsen P. Patterns in stem elongation rate in chrysanthemum and tomato plants in relation to irradiance and day/night temperature // Sci. Hort. 1994. V. 58. P. 139. https://doi.org/10.1016/0304-4238(94)90134-1
Gertsson U. Influence of temperature on shoot elongation in young tomato plants // Acta Hortic. 1992. V. 327. P. 71. https://doi.org/10.17660/ActaHortic.1992.327.8
Sysoeva M.I., Patil Grindal G., Sherudilo E.G., Torre S., Markovskaya E.F., Moe R. Effect of temperature drop and photoperiod on cold resistance in young cucumber plants – involvement of phytochrome B // Plant Stress. 2008. V. 2. P. 84.
Patterson B.D., Reid M.S. Genetic and environmental influences on the expression of chilling injury // Chilling Injury of Horticultural Crops. Ed. C.Y. Wang. Boca Raton, FL: CRC Press, 1990. P. 87.
Minorsky P.V. Temperature sensing by plants: a review and hypothesis // Plant Cell Environ. 1989. V. 12. P. 119. https://doi.org/10.1111/j.1365-3040.1989.tb01924.x
Minorsky P.V., Spanswick R.M. Electrophysiological evidence for a role for calcium in temperature sensing by roots of cucumber seedlings // Plant Cell Environ. 1989. V. 12. P. 137. https://doi.org/10.1111/j.1365-3040.1989.tb01925.x
Plieth C., Hansen U.P., Knight H., Knight M.R. Temperature sensing by plants: the primary characteristics of signal perception and calcium response // Plant J. 1999. V. 18. P. 491. https: //doi.org/10 1046/j.1365-313X.1999.00471.x
Nordin Henriksson K., Trewavas A.J. The effect of short-term low-temperature treatments on gene expression in Arabidopsis correlates with changes in intracellular Ca2+ levels // Plant Cell Environ. 2003. V. 26. P. 485. https://doi.org/10.1046/j.1365-3040.2003.00979.x
Zarka D.G., Vogel J.T., Cook D., Thomashow M.F. Cold induction of Arabidopsis CBF genes involves multiple ICE (inducer of CBF expression) promoter elements and a cold-regulatory circuit that is desensitized by low temperature // Plant Physiol. 2003. V. 133. P. 910. https://doi.org/10.1104/pp.103.027169
Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Особенности реакции растений Cucumis sativus L. на ДРОП в зависимости от скорости снижения температуры // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2016а. № 7. С. 121. https://doi.org/10.17076/eb489
Шибаева Т.Г., Икконен Е.Н., Шерудило Е.Г., Титов А.Ф. Особенности реакции различных по требовательности к свету гибридов Cucumis sativus L. на ежесуточные кратковременные показатели и урожайность // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2016. № 6. С. 56. https://doi.org/10.17076/eb287
Ikkonen E.N., Shibaeva T.G., Titov A.F. The role of light in cucumber plant response to a diurnal short-term temperature drop // J. Stress Physiol. Biochem. 2017. V. 13. P. 35.
Stavang J.A., Pettersen R.I., Wendell M., Solhaug K.A., Junttila O., Moe R., Olsen J.E. Thermoperiodic growth control by gibberellins does not involve changes in photosynthetic or respiratory capacities in pea // J. Exp. Bot. 2010. V. 61. P. 1015. https://doi.org/10.1093/jxb/erp366
Спиридонова Е.А., Сысоева М.И., Шерудило Е.Г. Влияние ежесуточных кратковременных снижений температуры и фотопериода на развитие декоративных растений // Ученые записки ПетрГУ. 2010. № 4. С. 18.
Oden P.C., Heide O.M. Quantification of gibberellins and indoleacetic acid in Begonia leaves: Relationship with environment, regeneration and flowering // Physiol. Plant. 1989. V. 76. P. 500. https://doi.org/10.1111/j.1399-3054.1989.tb05469.x
Zeevaart J.A.D., Talon M., Wilson T.M. Stem growth and gibberellin metabolism in spinach in relation to photoperiod // Gibberellins. Eds. N. Takahashi., B.O. Phinney, J. MacMillan. Springer-Verlag, 1991. P. 273.
Ikkonen E.N., Shibaeva T.G., Rosenquist E., Ottosen C.-O. Daily temperature drop prevents inhibition of photosynthesis in tomato plants under continuous light // Photosynthetica. 2015. V. 53. P. 389. https://doi.org/10.1007/s11099-015-0115-4
Шибаева Т.Г., Шерудило Е.Г. Действие и последействие ДРОП при круглосуточном освещении на рост и репродуктивное развитие томата // Физиология растений. 2015. Т. 62. С. 355. https://doi.org/10.7868/S0015330315030173
Шибаева Т.Г., Марковская Е.Ф., Икконен Е.Н., Шерудило Е.Г. Оценка эффективности ДРОП-обработки растений томата для предотвращения фотоповреждения листьев в условиях круглосуточного освещения // Российская сельскохозяйственная наука. 2015. Т. 5. С. 10.
Шибаева Т.Г., Шерудило Е.Г., Икконен Е.Н., Титов А.Ф. Влияние водного фактора на реакцию растений Cucumis sativus L. на кратковременное ежесуточное понижение температуры // Физиология растений. 2019. Т. 66. С. 230. https://doi.org/10.1134/S0015330319030138
Шибаева Т.Г., Титов А.Ф. Сравнительная оценка эффективности DROP-воздействий и “периодической засухи” как приемов управления ростом растений огурца (Cucumis sativus L.) // Сельскохозяйственная биология. 2019. Т. 54. С. 528. https://doi.org/10.15389/agrobiology.2019.3.528rus
Carvalho S.M.P., Noort F., van, Postma R., Heuvelink E. Possibilities for producing compact floricultural crops. Wageningen: Wageningen UR Greenhouse Horticulture, 2008. Report 173. 68 p.
Sysoeva M.I., Sherudilo E.G., Markovskaya E.F., Obshatko L.A., Matveeva E.M. Temperature drop as a tool for cold tolerance increment in plants // Plant Growth Regul. 2005. V. 46. P. 189. https://doi.org/10.1007/s10725-005-7357-2
Марковская Е.Ф., Сысоева М.И., Шерудило Е.Г. Феномен ежесуточного кратковременного влияния низких закаливающих температур на жизнедеятельность растения // Онтогенез. 2008. Т. 39. С. 323.
Hendriks L., Ludolph D., Menne A. Influence of different heating strategies on morphogenesis and flowering of ornamentals // Acta Hort. 1992. V. 305. P. 9. https://doi.org/10.17660/ActaHortic.1992.305.1
Jensen E., Eilertsen S., Ernstsen A., Junttila O., Moe R. Thermoperiodic control of stem elongation and endogenous gibberellins in Campanula isophylla // J. Plant Growth Regul. 1996. V. 15. P. 167. https://doi.org/10.1007/BF0019058
Grindal G., Ernstsen A., Reid J.B., Junttila O., Lindgard B., Moe R. Endogenous gibberellin A1 levels control thermoperiodic stem elongation in Pisum sativum // Physiol. plant. 1998. V. 102. P. 523. https://doi.org/10.1034/j.1399-3054.1998.1020406.x
Grindal G., Junttila O., Reid J.B., Moe R. The response to gibberellin in Pisum sativum grown under alternating day and night temperature // J. Plant Growth Regul. 1998. V. 17. P. 161. https://doi.org/10.1007/PL00007030
Moe R., Grindal G. Control of stem elongation: phytochrome and gibberellins involvement // J. Kor. Soc. Hort. Sci. 2000. V. 41. P. 662.
Patil G.G., Alm V., Moe R., Junttila O. Interaction between phytochrome B and gibberellins in thermoperiodic responses of cucumber // J. Am. Soc. Hortic. Sci. 2003. V. 128. P. 642. https://doi.org/10.21273/JASHS.128.5.0642
Runkle R., Blanchard M. Temperature integration // Greenhouse Product News. 2018. V. 2. P.
Икконен Е.Н., Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Влияние ДРОП-воздействий на эффективность использования световой энергии в процессе фотосинтеза у растений огурца // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2016. № 6. С. 49. https://doi.org/10.17076/eb319
Икконен Е.Н., Грабельных О.И., Шерудило Е.Г., Шибаева Т.Г. Устойчивое и чувствительное к салицилгидроксамовой кислоте дыхание теплолюбивых растений в условиях кратковременных ежесуточных понижений температуры // Физиология растений. 2020. Т. 67. С. 67. https://doi.org/10.1134/S0015330319050063
Икконен Е.Н., Шибаева Т.Г., Шерудило Е.Г., Титов А.Ф. Реакция дыхания проростков озимой пшеницы на продолжительное и кратковременное ежесуточное понижение температуры // Физиология растений. 2020. Т. 67. С. 312. https://doi.org/10.31857/S0015330320020062
Марковская Е.Ф., Сысоева М.И., Шерудило Е.Г., Топчиева Л.В. Дифференциальная экспрессия генов в растении огурца в ответ на многократные кратковременные низкотемпературные воздействия // Физиология растений. 2007. Т. 54. С. 686.
Лаврова В.В., Сысоева М.И., Шерудило Е.Г., Топчиева Л.В., Матвеева Е.М. Экспрессия гена ci7 в листьях картофеля при действии кратковременных ежесуточных снижений температуры // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2011. № 3. С. 73.
Марковская Е.Ф., Шерудило Е.Г., Рипатти П.О., Сысоева М.И. Роль липидов в устойчивости семядольных листьев огурца к постоянному и кратковременному периодическому действию низкой закаливающей температуры // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2009. № 3. С. 67.
Лаврова В.В., Сысоева М.И., Матвеева Е.М. Жирнокислотный состав липидов листьев картофеля в условиях периодической и длительной гипотермии // Труды КарНЦ РАН. Серия “Экспериментальная биология”. 2012. № 2. С. 91.
Bergstrand K.-J.I. Methods for growth regulation of greenhouse produced ornamental pot- and bedding plants – a current review // Folia Hort. 2017. V. 29. P. 63. https://doi.org/10/1515/fhort-2017-0007
Körner O., Challa H. Energy saving climate control regime for cut chrysanthemum // Acta Hort. 2004. V. 633. P. 489. https://doi.org/10.17660/ActaHortic.2004.633.61
Rijsdijk A.A., Vogelezang J.V.M. Temperature integration on a 24-hour base: a more efficient climate control strategy // Acta Hort. 2000. V. 519. P. 163. https://doi.org/10.17660/ActaHortic.2000.519.16
Elings A., Kempkes F.L.K., Kaarsemaker R.C., Ruijs M.N.A., Van de Braak N.J., Dueck T.A. The energy balance and energy-saving measures in greenhouse tomato cultivation // Acta Hort. 2005. V. 691. P. 67. https://doi.org/10.17660/ActaHortic.2005.691.5
Ottosen C.-O., Rosenqvist E., Aaslyng J.M., Jakobsen L. Dynamic climate control in combination with average temperature control saves energy in ornamentals // Acta Hort. 2004. V. 691. P. 133. https://doi.org/10.17660/ActaHortic.2005.691.14
Lopez R.G., Runkle E.S. Growing your crops above their base temperature // Greenhouse Grower. 2014. V. 6. P. 53.
Los D.A., Murata N. Membrane fluidity and its roles in the perception of environmental signals // Biochem. Biophys. Acta. 2004. V. 1666. P. 142.
Ruelland E., Vaultier M.N., Zachowski A., Hurry V. Cold signalling and cold acclimation in plants // Adv. Bot. Res. 2009. V. 49. P. 35.
Марковская Е.Ф., Шибаева Т.Г. Низкотемпературные сенсоры у растений: гипотезы и предположения // Известия РАН. Серия биологическая. 2017. № 2. С. 120. https://doi.org/10.7868/S000233291702014X
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений