

Физиология растений, 2021, T. 68, № 3, стр. 315-325
Сезонные изменения содержания хлорофиллов и каротиноидов в хвое сосны обыкновенной (Pinus sylvestris L.) в зоне влияния теплового поля газового факела
С. А. Шавнин a, *, И. А. Юсупов a, Н. В. Марина b, А. А. Монтиле a, **, Д. Ю. Голиков a
a Федеральное государственное бюджетное учреждение науки
“Ботанический сад Уральского отделения Российской академии наук”
Екатеринбург, Россия
b Федеральное государственное бюджетное образовательное учреждение высшего образования
“Уральский государственный лесотехнический университет”
Екатеринбург, Россия
* E-mail: sash@botgard.uran.ru
** E-mail: org17@mail.ru
Поступила в редакцию 25.06.2020
После доработки 02.07.2020
Принята к публикации 02.07.2020
Аннотация
Цель работы – установление закономерностей и проведение сравнительного анализа сезонных изменений содержания хлорофиллов и каротиноидов в хвое молодых деревьев сосны обыкновенной (Pinus sylvestris L.), произрастающих на разном удалении от факела сжигания нефтяного газа, в зоне создаваемого им теплового поля на территории Ханты-Мансийского автономного округа (Югры), в течение периода “осень–зима–весна”. В ходе исследований проведена верификация двух гипотез: (1) тепловое поле факела сжигания нефтяного газа, имитирующее действие потепления климата на растения, влияет на состояние пигментного комплекса фотосинтетического аппарата (ФСА) хвои сосны обыкновенной; (2) трансформация пигментного комплекса ФСА хвои в течение периода “осень–зима–весна” чувствительна к долговременному повышению температуры среды на 1–2°С. В сезонных динамиках отдельных показателей ФСА в течение зимы наблюдаются максимумы и минимумы, объясняемые регуляцией содержания хлорофиллов и каротиноидов при изменении температуры воздуха. Сезонные изменения этих показателей у деревьев, произрастающих в условиях с отличающимся температурным режимом, не совпадают. Их закономерности свидетельствуют о последовательном изменении глубины зимней трансформации структурно-функционального состояния ФСА хвои на разном удалении от факела. Активность ФСА хвои в течение всего изучаемого периода была выше вблизи факела и уменьшалась по мере удаления от него. В осенне-зимний период, когда для хвои характерно уменьшение содержания хлорофилла, это снижение наименьшее вблизи факела (секция I), где температура выше фоновой на 1–2°С, среднее – на среднем расстоянии от факела (секция III) и наибольшее – на максимальном удалении от факела (секция VII). При этом соотношение содержания хлорофиллов a и b в хвое деревьев на секции I стабильно ниже, чем на секции VII. Это указывает на то, что в хлоропластах хвои деревьев, расположенных в секции I, количество светособирающих комплексов больше количества фотосистем. Величина отношения общего содержания хлорофиллов a и b к содержанию каротиноидов в хвое деревьев вблизи факела выше, чем на секциях III и VII, а также имеет в сезонной динамике два достоверных максимума, что объясняется повышенной сохранностью пула зеленых пигментов. Отличия сезонных изменений содержания хлорофиллов и каротиноидов в хвое и корреляционных связей данных показателей с температурой воздуха указывают на различие механизмов их регуляции. Количество хлорофиллов определяется балансом процессов распада и синтеза, результат которого зависит от влияния факторов среды на скорость синтеза хлорофиллов и реализацию защитных функций каротиноидами. Последние дополнительно синтезируются в фотосинтезирующих клетках в осенне-зимний период, обеспечивая увеличение степени защищенности хлорофиллов от фотодеструкции с осени до конца весны.
ВВЕДЕНИЕ
Сезонная динамика структурно-функциональной организации пигментного аппарата (ПА) хлоропластов листьев является проявлением адаптации фотосинтетического аппарата (ФСА) к действию изменяющихся условий среды. Актуальность ее изучения определяется недостаточной исследованностью механизмов сезонной трансформации ФСА, в том числе на уровне первичных фотосинтетических реакций преобразования энергии. Изучение этих процессов у вечнозеленых растений, произрастающих в климатических зонах с холодной зимой, позволило установить ряд основных закономерностей. В годичном цикле выделяют два основных этапа: уменьшение содержания хлорофиллов a и b (Хл a и Хл b) в осенне-зимний период и восстановление их пулов в начале активной вегетации весной и летом [1–4]. На Среднем Урале максимальное содержание хлорофиллов в двухлетней хвое молодых деревьев сосны обыкновенной (Pinus sylvestris L.) приходится на летний период, снижается в сентябре–феврале, продолжает уменьшаться в марте–апреле и восстанавливается в мае [1]. Следует отметить, что в северных широтах в структуре ПА наблюдается преобладание количества хлорофиллов в ССК по сравнению с фотосистемами [4, 5], свидетельствующее о высокой светопоглощающей способности хвои, позволяющей компенсировать низкий уровень накопления зеленых пигментов в этих условиях.
Содержание каротиноидов (Кар) в хвое отдельных видов ели, пихты и сосны обыкновенной возрастает зимой и уменьшается весной [2, 4–8]. При этом с наступлением зимы у большинства видов хвойных наблюдается уменьшение отношения суммарного содержания хлорофиллов к Кар, которое объясняется защитной функцией каротиноидов. Они предохраняют ассимиляционный аппарат от фотодинамического разрушения в условиях низких температур за счет тушения синглетного кислорода, поглощения и рассеивания энергии возбуждения молекул хлорофиллов и стабилизации физического состояния мембран и белков антенных комплексов [9, 10]. Роль каротиноидов, по-видимому, возрастает в условиях снижения интенсивности физиолого-биохимических процессов в клетках хвои при низких температурах. У хвойных видов растений основным механизмом фотозащиты в зимне-весенний период является зеаксантин-зависимая тепловая диссипация энергии возбуждения в реакционных центрах ФС II [2, 11].
Качественные и количественные изменения ПА хлоропластов хвои при переходе в зимнее состояние сопровождаются рядом структурных перестроек в клетках фотосинтетических тканей. В них происходят последовательный распад гранальной структуры и уменьшение числа тилакоидов в хлоропластах, уменьшение объема и дробление вакуолей, перераспределение пластид внутри клетки и трансформация мембранных структур [12–14]. В клетках мезофилла хвои ели сибирской (Picea obovata Ledeb.) и ели колючей (Picea pungens Engelm.) исчезают зерна крахмала, а также изменяются форма и локализация хлоропластов [15]. Зимние условия приводят к ингибированию фотохимической активности ФС II, подавлению нециклического электронного транспорта в хлоропластах [16, 17], частичному разрушению хлорофилл-белковых ССК [18] и изменению липидного состава мембран хлоропластов [19, 20]. На фоне этих процессов в хвое сосны обыкновенной наблюдаются последовательные изменения температурной зависимости выхода флуоресценции Хл a при полностью открытых реакционных центрах ФС II [1, 21], величины миллисекундной компоненты флуоресценции Хл a ФС II и параметров ее индукционных кривых [22]. В условиях Среднего Урала снижение фотохимической активности ФСА продолжается на протяжении всего зимнего периода, а ее минимальные значения наблюдаются в феврале.
Температурный режим относится к числу основных факторов среды, оказывающих влияние на состояние ПА [23–25], а также на скорость депонирования углерода и продуктивность лесных экосистем бореальной зоны. Глобальные изменения климата в ряде регионов сопровождаются повышением температуры среды на 1–2°С за несколько десятков лет (https://unstats.un.org/sdgs/report/ 2019/The-Sustainable-Development-Goals-Report-2019.pdf). Скорость приращения среднегодовой температуры воздуха на территории Российской Федерации с 1976 по 2018 гг. составила 0.47°С за 10 лет [26]. При этом изучение влияния климатических изменений на реальные природные объекты связано с преодолением значительных технических и методических трудностей. Одним из перспективных методических подходов к изучению влияния потепления климата на растения является исследование состояния деревьев, произрастающих в максимально приближенных к естественным условиях среды, модифицированных тепловым полем длительно функционирующего факела сжигания нефтяного газа [27]. Проведение наблюдений в зоне теплового воздействия факела позволяет исследовать комплекс адаптационных реакций, происходящих в ФСА листьев растений при повышении температуры среды.
При планировании наших исследований было выдвинуто две рабочие гипотезы: (1) тепловое поле факела сжигания нефтяного газа, имитирующее действие потепления климата на растения, влияет на состояние пигментного комплекса ФСА хвои сосны обыкновенной (одного из основных лесообразующих видов северо-таежной подзоны таежной зоны); (2) трансформация состояния пигментного комплекса ФСА хвои в течение периода “осень–зима–весна” чувствительна к долговременному повышению температуры среды на 1–2°С. Подтверждением гипотезы 1 будет являться несовпадение сезонных динамик показателей состояния пигментного комплекса ФСА хвои деревьев сосны обыкновенной, произрастающих в условиях с отличающимся температурным режимом (на разном удалении от факела). При этом на наибольшем расстоянии от факела температура среды соответствует фоновой. Гипотеза 2 будет подтверждена, если закономерности изучаемых сезонных динамик будут свидетельствовать о последовательном изменении глубины зимней трансформации структурно-функционального состояния ФСА хвои на разном удалении от факела. В соответствии с вышеприведенными гипотезами, цель работы – установить закономерности и провести сравнительный анализ сезонных изменений содержания хлорофиллов и каротиноидов в хвое деревьев сосны обыкновенной, произрастающих на разном удалении от факела сжигания нефтяного газа в зоне создаваемого им теплового поля, в течение периода “осень–зима–весна”.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили на территории Покачевского участкового лесничества Мегионского лесничества Ханты-Мансийского автономного округа – Югры. Климат Ханты-Мансийского региона континентальный. За последние 10 лет в районе исследований среднегодовая температура воздуха составляла –3.6°C. Самым холодным месяцем является январь (средняя температура –22°C), а самым теплым – июль (средняя температура +16.9°C). При этом в течение зимнего периода наблюдаются значительные изменения усредненных по декадам среднесуточных температур (рис. 1). Безморозный период составляет 98 дней, а вегетационный – 131 день. Среднегодовое количество осадков – 492 мм с максимумом в июле–сентябре (239 мм). Продолжительность сохранения снежного покрова составляет 201 день, а его максимальная высота (77–85 см) наблюдается в третьей декаде марта.
Рис. 1.
Средние за декаду температуры воздуха в период проведения исследований по данным метеостанции г. Ханты-Мансийска (Погодные сервисы: архив погоды, http://pogoda-service.ru).

Объектом исследований являлись молодняки сосны обыкновенной (Pinus sylvestris L.), в которых вблизи газового факела четвертой дожимной насосной станции Покачевского месторождения нефти по стандартной методике была заложена постоянная пробная площадь. Древостой естественного возобновления произрастает на участке вырубленного спелого сосняка. На пробной площади выделено семь секций (I–VII) шириной 10 м и длиной 60 м каждая, которые располагаются перпендикулярно радиусу к факелу. Ближние границы секций I, III и VII находятся на расстоянии 70 м, 90 м и 130 м от факела соответственно. Тип леса – сосняк лишайниковый, древостой одноярусный, чистый 10С, возраст 12–15 лет, III–IV классов бонитета, густота – 2900–6400 шт/га, средние диаметры и высоты отличаются на разных секциях (2.5–5.6 см и 1.3–2.3 м соответственно) [27].
Температура среды на секции VII соответствует фоновой. Разность температур воздуха в тепловом поле факела между секциями I и VII на высоте 1.3 м от уровня земли составляет до 1.5°C, а почвы – до 6°C [28]. Приведенные значения характерны для общей конфигурации теплового поля. В связи с мозаичностью микрорельефа и неоднородностью растительности в пределах секции, температура на разных участках не одинакова и изменяется в течение суток и всего вегетационного периода. Тенденция к увеличению температуры с приближением к факелу достоверна, поэтому влияние теплового поля изучалось с помощью закладки секций, расположенных на различном расстоянии от него.
В исследованиях ПА использовали хвою предыдущего года формирования. В период с 15.08.2004 г. по 30.05.2005 г. образцы хвои отбирали 1–2 раза в месяц на высоте 1.5 м с боковых ветвей обращенной к факелу средней части кроны пяти модельных (средних по диаметру и высоте) деревьев на секциях I, III и VII. Образцы хранили в течение 24–26 ч в термостате при температуре, соответствующей температуре воздуха в момент сбора. С целью уменьшения неизбежного, ввиду большого объема собираемого материала, влияния дефолиации на состояние остающейся хвои, образцы отбирали равномерно из разных частей побегов. Для анализов использовали метод средней пробы. Каждое измерение проводили в трех биологических повторностях. Пигменты экстрагировали ацетоном (80%) из гомогената хвои при температуре 4–6°C. Содержание пигментов определяли с помощью измерения оптической плотности фильтрованных экстрактов на спектрофотометре СФ-46 (“ЛОМО”, Россия) при длинах волн 470, 646.8 и 663.2 нм [29], а их количество оценивали пересчетом на массу абсолютно сухой хвои. Для оценки соотношения количеств ССК и ФС I и ФС II хлоропластов использовали отношение Хл a/Хл b, а при изучении особенностей строения ПА – отношение суммарного содержания хлорофиллов (Хл (a + b)) к Кар (Хл (a + b)/Кар).
Статистический анализ полученных данных проводили с помощью стандартных пакетов прикладных программ Statistica 8.0 (StatSoft Inc., 2007) и Excel (Microsoft, 2007). Использовали корреляционный и регрессионный анализы. На графиках динамик измеренных и расчетных показателей приведены средние величины и стандартные ошибки. При корреляционном анализе сезонных динамик изменения параметров пигментного комплекса с изменением температуры приземного воздуха использовали значения средних декадных температур, а при отборе проб в начале декады – данные о температуре за предшествующую декаду.
РЕЗУЛЬТАТЫ
Анализ сезонных изменений Хл a в хвое (рис. 2) позволяет выделить три основных этапа. На первом (середина августа – середина сентября) величина показателя максимальна, затем в конце сентября она резко уменьшается и наступает второй этап, который продолжается до первой декады мая. В этот период значение Хл a минимально, относительно стабильно и совпадает на секциях III и VII, а на секции I достоверно возрастает и проявляет два максимума – в ноябре и феврале, которые по величине не достигают летних значений. В конце мая наступает следующий (третий) этап, на котором Хл а в хвое возрастает до значений августа на секции I, а на секциях III и VII наблюдается тенденция к его восстановлению до летних величин. Относительное сходство сезонных динамик Хл a у деревьев на разных секциях, несмотря на отличия абсолютных величин, подтверждается высокими значениями их парных корреляций (r = = 0.74–0.81, P < 0.05). При этом сезонные изменения Хл a на всех секциях коррелируют с динамикой температур воздуха (r = 0.58–0.67, P < 0.05).

Сезонные изменения Хл b в хвое (рис. 3) в целом аналогичны наблюдаемым динамикам Хл a по срокам наступления этапов и основным закономерностям. Величина этого показателя в течение всего периода исследований, как правило, выше на секции I, минимальна на секции VII и имеет промежуточные (или совпадающие с секцией VII) значения на секции III. Кроме того, по сравнению с Хл a, восстановление уровня Хл b на секции I на втором этапе более полное и в периоды зимних максимумов его значения совпадают с показаниями августа. Промежуточное положение сезонных изменений показателя на секции III подтверждается результатами корреляционного анализа, согласно которым динамики на секциях I и III достоверно не коррелируют, но существуют связи между секциями I и VII (r = 0.59, P < 0.05), а также III и VII (r = 0.67, P < 0.05). Следует отметить, что величины корреляционной связи Хл b в хвое с температурой воздуха на секциях I и III одинаковы (r = 0.5, r = 0.45, P < 0.05) и несколько выше на секции VII (r = 0.69, P < 0.05).

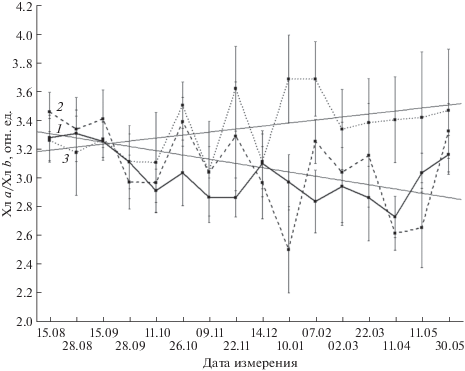
Сезонные изменения величины отношения Хл a/Хл b (рис. 4) также рассматриваются с выделением трех этапов трансформации ФСА хвои, при этом их общие закономерности на разных секциях не совпадают. Наиболее плавно (с наименьшими амплитудами варьирования) величины Хл a/Хл b изменяются на секции I, на которой первый этап (август–середина сентября) характеризуется периодом стабильности, сменяемым понижением (до середины октября), после чего наступает второй этап, когда уровень Хл a/Хл b в хвое устойчиво низкий. В мае происходит его восстановление до летних значений (третий этап). На секции VII значения показателя в течение всего периода исследований менее устойчивы при общем слабом положительном линейном тренде. Например, в период с 14 декабря по 10 января наблюдается резкое увеличение Хл a/Хл b в хвое (на 20% относительно величины в декабре). На секции III варьирование Хл a/Хл b выше, а линейный тренд его значений отрицателен. В течение зимы значения Хл a/Хл b, как правило, являются промежуточными между соответствующими значениями для секций I и VII, а на третьем этапе в мае наблюдается их восстановление до уровня августа. Следует отметить, что на относительно слабое сходство сезонных динамик Хл a/Хл b в хвое деревьев на разных секциях указывает отсутствие достоверных парных корреляционных связей между ними. При этом на ближней к факелу секции существует достаточно высокая связь (r = 0.65, P < 0.05) значений Хл a/Хл b с температурой воздуха, которая значительно слабее на секциях III и VII (r = 0.43 и 0.3 соответственно, P < 0.05).
Рис. 4.
Сезонная динамика отношения Хл a/Хл b в хвое Pinus sylvestris L. на секциях I (1), III (2) и VII (3).
Величина Кар в хвое возрастает в сентябре (рис. 5), затем этот показатель на секциях I и III стабилизируется до начала февраля и сохраняется до середины мая, после чего снижается в конце мая до наблюдавшегося в августе уровня.
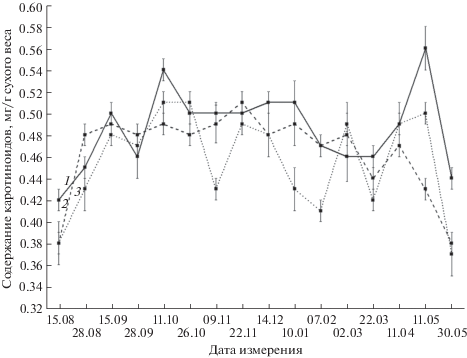
На секции VII Кар в хвое варьирует в течение всей зимы, а в мае также наблюдается аналогичное другим секциям снижение этого показателя. В течение всего периода исследований величина Кар в хвое на ближней к факелу секции I, как правило, выше, чем на фоновой секции VII. При этом сезонная динамика показателя на секции VII коррелирует с секциями I и III (r = 0.71 и 0.64 соответственно; P < 0.05), однако парной кореляции между секциями I и III не наблюдается. Следует отметить также, что величина Кар в хвое не коррелирует с температурой воздуха ни на одной из секций.
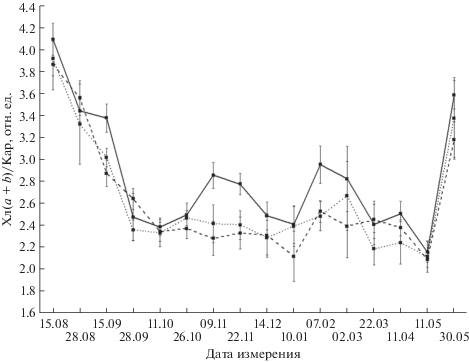
В сезонных динамиках отношения Хл (a + b)/Кар в хвое (рис. 6) явно выделяются три характерных, как и для Хл a и Хл b, этапа (снижение–относительная стабилизация–возрастание). При этом величина данного показателя на всех секциях значительно уменьшается в течение августа–сентября, после чего сохраняется на относительно стабильном низком уровне до первой декады мая, а в конце мая проявляет тенденцию к восстановлению до значений августа. Следует отметить, что на ближней к факелу секции I наблюдались периоды достоверного возрастания величины Хл (a + b)/Кар в хвое в ноябре и феврале, обусловленные относительным увеличением Хл b на фоне стабильных значений размеров пула каротиноидов. На секциях III и VII Хл (a + b)/Кар в течение зимы варьируют незначительно. Минимальные значения этого показателя достигаются на всех секциях в начале мая. Особо следует отметить высокие значения линейных парных корреляций сезонных динамик этой характеристики ФСА на всех трех секциях I, III и VII (r = 0.91–0.96, P < 0.05), а также их достоверную связь с температурой воздуха (r = 0.6, 0.69 и 0.66 соответственно; P < 0.05).
ОБСУЖДЕНИЕ
Анализ сезонных изменений Хл a и Хл b в хвое сосны в изучавшийся период показал, что их общая направленность одинакова на всех секциях (уровень значений максимален в последний месяц лета и минимален зимой) и соответствует известной [1]. Частичное восстановление величин Хл a и Хл b происходит в конце мая и наиболее выражено вблизи факела на секции I. При этом наиболее высокое содержание хлорофиллов соответствует активному состоянию ФСА хвои на завершающем этапе вегетации и в начале сезона роста следующего года. Осенью происходит закономерное снижение содержания хлорофиллов в хвое, связанное с наступлением низких температур, переходом к зимнему состоянию и торможением биохимических процессов. Выход из зимнего покоя начинается весной после локального минимума в конце марта.
Совокупность известных сведений о последовательных сезонных изменениях ПА хвои, структурных перестройках ФСА на уровне клеток и хлоропластов, а также о трансформации флуоресцентных характеристик Хл a в осенне-зимний период указывает на возможность существования в хвое вечнозеленых растений при низких температурах активных адаптационных процессов, включая биосинтез отдельных пигментов. Следует отметить, что уровень значений зимних температур в районе исследований не постоянен (в отдельные месяцы средняя температура изменяется на 10–15°C и более), что является, по-видимому, фактором, способным влиять на общее состояние ФСА. На это указывают факты наличия корреляционных связей сезонных динамик большинства показателей состояния ПА хвои с температурой среды, а также существование локальных по времени увеличений и уменьшений значений этих показателей, в том числе в середине зимы. Содержание зеленых пигментов в хвое сосны вблизи факела (секция I) в среднем выше в течение всего изучаемого периода и с удалением от него уменьшается. Данная закономерность является одним из основных подтверждений первой гипотезы. Кроме того, на секции I зимой наблюдается статистически значимое частичное восстановление пулов Хл a и Хл b в хвое. Это свидетельствует о повышенной активности ФСА хвои на секции I по сравнению с другими, и, соответственно, о существенном влиянии факела на состояние ПА. Наибольшие величины Хл b в хвое на секции I, наименьшие на секции VII и промежуточные на секции III в зимний период свидетельствуют о том, что, во-первых, влияние факела распространяется на расстояние не менее 100 м, и, во-вторых, что это влияние на секции III меньше, чем на секции I. Наличие нескольких подъемов и спадов на кривых сезонных динамик содержания Хл a и Хл b в хвое на секции I, а также достоверных, но менее выраженных изменений этих показателей на секциях III и VII зимой, указывает на продолжение в этот период процессов сезонной трансформации состояния ПА хлоропластов.
На удаленных от факела секциях средний уровень величин отношения Хл a/Хл b в хвое выше, чем на ближней к нему секции. Сезонные динамики этого показателя указывают на уменьшение в хлоропластах относительного количества ССК по сравнению с количеством ФС I и ФС II зимой. Данный эффект наиболее выражен на фоновой секции и наименее – вблизи факела. На секции I Хл a/Хл b в хвое зимой устойчиво низкое, в том числе за счет частичного восстановления пула Хл b, обеспечивающего более высокий уровень количества ССК. Таким образом, на фоне общей зимней депрессии ПА хвои, степень его деградации максимальная за пределами зоны влияния теплового поля факела на секции VII, наименьшая – на секции I, а на секции III – промежуточная. Причем на всех секциях соотношение количеств ССК и фотосистем регулируется преимущественно за счет изменения пула Хл b. Отличие величин парных корреляций сезонных динамик хлорофиллов, наблюдаемых на отдельных секциях, подтверждает несовпадение в изучаемый период параметров адаптационных процессов в ФСА хвои на разном удалении от факела.
Увеличение Кар в хвое зимой соответствует известным данным, полученным в ряде исследований состояния ПА вечнозеленых растений [30], и объясняется участием каротиноидов в защите молекул хлорофиллов от фотодеструкции в этот период. Их роль особенно возрастает в условиях снижения интенсивности физиолого-биохимических процессов в клетках хвои при низких температурах. Следует отметить, что корреляционный анализ сезонных динамик содержания хлорофиллов и Кар в хвое не выявил достоверной связи между ними на всех секциях. Кроме того, в отличие от Хл a и Хл b, не установлено корреляций Кар с температурой среды. Эти факты указывают на относительную независимость регуляции величин пулов данных групп пигментов ФСА.
Сравнительно устойчивый повышенный уровень Кар в хвое в период сентябрь–апрель наблюдается только на ближних к факелу секциях, а на секции VII в сезонной динамике существуют отдельные подъемы и спады. Эта особенность объясняется, по-видимому, отличающимся от наблюдаемого на секциях I и III характером проявления процесса регуляции размеров каротиноидного комплекса на фоновом участке, где влияние теплового поля факела отсутствует.
Уменьшение в осенне-зимний период Хл (a + + b)/Кар в хвое наблюдалось также другими исследователями [11]. Согласно полученным данным начало уменьшения этого параметра соответствует снижению среднесуточной температуры в октябре до значений ниже 5°C. Общее сходство сезонных динамик этого показателя на разных секциях, подтверждаемое высокой степенью их корреляции между собой, свидетельствует о важности связанных с соотношением Хл (a + b)/Кар адаптационных механизмов для перехода ПА хлоропластов к зимнему покою. В этом состоянии величина Хл (a + b)/Кар составляет 2.4–2.8 и обеспечивает, по-видимому, необходимую сохранность ФСА хвои на уровне первичных звеньев трансформации световой энергии. Наличие локальных максимумов в ноябре и феврале на секции I дополнительно подтверждает высказанное выше утверждение о большей активности ФСА хвои зимой вблизи факела. Эти относительные максимумы возникают в результате увеличения Хл a и Хл b (на фоне относительно стабильного Кар), указывающего на смещение в эти месяцы баланса процессов синтеза и распада хлорофиллов в сторону синтеза.
Результаты анализа особенностей сезонных динамик содержания отдельных пигментов и их соотношений указывают на непрерывность адаптационных реакций ФСА хвои в течение всего осенне-зимнего периода и их зависимость от удаления от факела, что доказывает справедливость второй гипотезы. Кроме того, они позволяют выделить ряд основных закономерностей трансформации ПА хлоропластов хвои сосны в течение изучавшегося периода, а также высказать ряд предположений о природе процессов и механизмов, объясняющих эти закономерности. Сезонное понижение температуры среды приводит к замедлению, но не к полному прекращению физиолого-биохимических процессов, которое характерно для состояния ФСА зимой. При этом в результате фотоокисления хлорофиллов, снижается их содержание до определенного уровня, который поддерживается за счет защитных функций комплекса каротиноидов, а также биосинтеза отдельных пигментов. Величина соотношения хлорофиллов ССК и фотосистем хлоропластов варьирует при фоновых условиях среды вокруг среднего значения, в том числе в условиях низких температур, так как отражает работу систем контроля содержания зеленых пигментов. Количество каротиноидов в ССК и ФС возрастает зимой, что обеспечивает сохранение ПА на уровне, соответствующем минимально активному состоянию ФСА хвои. Регуляция Кар происходит относительно независимо от содержания хлорофиллов, однако она сопряжена, по-видимому, с поддержанием в определенном диапазоне значений величины отношения Хл (a + b)/Кар. Влияние газового факела, в первую очередь связанное с повышением температуры воздуха, обеспечивает более высокую активность ПА хлоропластов за счет реализации возможностей адаптационного потенциала ФСА хвои, включая регуляцию размеров пулов отдельных пигментов.
Анализ полученных результатов позволяет заключить следующее. (1) Сезонная трансформация ПА хлоропластов хвои сосны обыкновенной, наблюдаемая в период “конец лета–осень–зима–весна” связана с уменьшением температуры среды ниже физиологического оптимума активной вегетации и представляет собой совокупность последовательно протекающих во времени процессов адаптации ФСА хвои к изменению условий среды. Многократность наблюдений в изучаемый период позволяет дополнить известные сведения об особенностях изменений состояния ПА. Также установлено наличие локальных по времени максимумов и минимумов в сезонной динамике его отдельных показателей в течение зимы, связанное, по-видимому, с регуляцией содержания хлорофиллов и каротиноидов при изменении температуры среды.
(2) Тепловое поле факела влияет на протекание адаптационных процессов, определяющих закономерности сезонных динамик показателей состояния ПА хвои деревьев сосны обыкновенной на разном удалении от его источника. Его действие приводит к изменениям сезонных динамик всех изученных показателей состояния ФСА хвои. Активность ФСА хвои в течение всего изучаемого периода выше вблизи факела и уменьшается по мере удаления от него. В осенне-зимний период вблизи факела (секция I), где температура выше фоновой на 1–2°С, снижение содержания хлорофиллов меньше, чем на максимальном удалении от него (секция VII). На среднем расстоянии от факела (секция III) эти эффекты выражены слабее. При этом отношение Хл a/Хл b на секции I устойчиво ниже, чем на VII, что указывает на увеличение соотношения количеств ССК и фотосистем в хлоропластах. Величина Хл (a + b)/Кар на секции I выше, чем на секциях III и VII и имеет два достоверных максимума, что связано, по-видимому, с повышенной сохранностью пула зеленых пигментов за счет защиты хлорофиллов от фотодеструкции вблизи факела. Полученные в ходе исследований результаты подтверждают правильность обеих выдвинутых рабочих гипотез.
(3) Отличие сезонных изменений содержания хлорофиллов и Кар в хвое, а также наличие корреляций сезонных динамик величин Хл a и Хл b с температурой воздуха независимо от расстояния до факела и отсутствие такой связи у пула Кар, указывают на различие механизмов их регуляции. При этом, по-видимому, количество хлорофиллов определяется балансом процессов распада и синтеза, результат которого зависит от влияния факторов среды, включая температуру, реализации защитных функций каротиноидами, скорости синтеза хлорофиллов и активности ФСА хвои в целом. Данный баланс определяет соотношение количества хлорофиллов фотосистем и ССК в хлоропластах, которое в фоновых условиях среды варьирует относительно среднего уровня в течение года. При этом в осенне-зимний период в фотосинтезирующих клетках дополнительно синтезируются каротиноиды, что увеличивает степень защищенности хлорофиллов от фотодеструкции. Регуляция Кар, по-видимому, осуществляется в соответствии с величиной Хл (a + b)/Кар, которая достигает минимума и поддерживается на низком уровне в период с осени до конца весны.
Авторы выражают благодарность ведущему инженеру Уральского государственного лесотехнического университета Г. Н. Новоселовой за неоценимую помощь в проведении лабораторных исследований.
Работа выполнена при финансовой поддержке Комплексной программы Уральского отделения Российской академии наук на 2018–2020 гг. (грант № 18-4-4-10) и в рамках госзадания Федерального государственного бюджетного учреждения науки “Ботанического сада Уральского отделения Российской академии наук”.
Авторы заявляют об отсутствии конфликта интересов. Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов исследований.
Список литературы
Кривошеева А.А., Шавнин С.А., Калинин В.А., Венедиктов П.С. Влияние промышленных загрязнений на сезонные изменения содержания хлорофилла в хвое сосны обыкновенной // Физиология растений. 1991. Т. 38. С. 163.
Ensminger I., Sveshnikov D., Campbell D.A., Funk C., Jansson S., Lloyd J., Shibistova O., Quist G. Intermittent low temperatures constrain spring recovery of photosynthesis in boreal Scots pine forests // Global Change Biology. 2004. V. 10. P. 1. https://doi.org/10.1111/j.1365-2486.2004.00781.x
Яцко Я.Н., Дымова О.В., Головко Т.К. Пигментный комплекс зимне- и вечнозеленых растений в подзоне средней тайги европейского Северо-Востока // Ботан-. журн. 2009. Т. 94. С. 1812.
Тужилкина В.В. Фотосинтетические пигменты хвои ели сибирской в среднетаежных лесах европейского северо-востока России // Сибирский лесной журнал. 2017. № 1. С. 65. https://doi.org/10.15372/SJFS20170107
Yatsko Y.N., Dymova O.V., Golovko T.K. Violaxanthin cycle pigment de-epoxidation and thermal dissipation of light energy in three boreal species of evergreen conifer plants // Russ. J. Plant Physiol. 2011. V. 58. P. 169. https://doi.org/10.1134/S1021443711010249
Ottander C., Campbell D., Öquist G. Seasonal Changes in Photosystem II Organization and Pigment Composition in Pinus sylvestris // Planta. 1995. V. 197. P. 176. https://doi.org/10.1007/BF00239954
Wang K.-Y., Kellomäki S., Zha T. Modifications in photosynthetic pigments and chlorophyll fluorescence in 20-year-old pine trees after a four-year exposure to carbon dioxide and temperature elevation // Photosynthetica. 2003. V. 41. P. 167. https://doi.org/10.1023/B:PHOT.0000011948.00870.db
Тужилкина В.В. Пигментный комплекс хвои сосны в лесах Европейского Северо-востока // Лесоведение. 2012. № 4. С. 16.
Kalituho L., Rech J., Jahns P. The roles of specific xanthophylls in light utilization // Planta. 2007. V. 225. P. 423. https://doi.org/10.1007/s00425-006-0356-3
Öguist G., Huner N.P.A. Photosynthesis of overwintering evergreen plants // Annu. Rev. Plant Biol. 2003. V. 54. P. 329. https://doi.org/10.1146/annurev.arplant.54.072402.115741
Sofronova V.E., Dymova O.V., Golovko T.K., Chepalov V.A., Petrov K.A. Adaptive changes in pigment complex of Pinus sylvestris needles upon cold acclimation // Russ. J. Plant Physiol. 2016. V. 63. P. 433. https://doi.org/10.1134/S1021443716040142
Ходасевич Э.В., Арнаутова А.И., Мышковец С.Н. Ультраструктурная организация хлоропластов в связи с обратимой деградацией фонда пигментов у хвойных. // Физиология растений. 1978. Т. 25. С. 810.
Martin B., Öquist G. Seasonal and experimentally induced changes in the ultrastructure of chloroplasts of Pinus sylvestris // Physiol. Plantarum. 1979. V. 46. P. 42. https://doi.org/10.1111/j.1399-3054.1979.tb03183.x
Маслова Т.Г., Мамушина Н.С., Зубкова Е.К., Буболо Л.С., Тютерева Е.В. Структурно-функциональная характеристика фотосинтетического аппарата листа зимнезеленых растений в ходе круглогодичной вегетации // Ботан. журнал. 2015. Т. 100. С. 1142. https://doi.org/10.1134/S0006813615110022
Овсянников А.Ю., Семкина Л.А. Сезонные изменения активности фотосистемы II и локализации хлоропластов в клетках хвои растений рода Picea (Pinacea) // Ботан. журн. 2014. Т. 99. С. 971.
Öquist G. Martin B. Inhibition of photosynthetic electron transport and formation of inactive chlorophyll in winter stressed Pinus sylvestris // Physiol. Plantarum. 1980. V. 48. P. 33. https://doi.org/10.1111/j.1399-3054.1980.tb03215.x
Öquist G.; Ogren E. Effects of winter stress on photosynthetic electron transport and energy distribution between the two photosystems of pine as assayed by chlorophyll fluorescence kinetics // Photosynthetic Research. 1985. V. 7. P. 19. https://doi.org/10.1007/BF00032919
Öquist G., Martensson O., Martin B., Malmberg G. Seasonal effects on chlorophyll-protein complexes isolated from Pinus sylvestris // Physiol. Plantarum. 1978. V. 44. P. 187. https://doi.org/10.1111/j.1399-3054.1978.tb08616.x
Новицкая Ю.Е. Сезонная и возрастная динамика основных фракций липидов в хвое сосны обыкновенной // Физиолого-биохимические исследования сосны обыкновенной на Севере. Петрозаводск. 1987. С. 39.
Öquist G. Seasonally induced changes in acid lipids and fatty acids of chloroplast thylakoids of Pinus sylvestris // Plant Physiol. 1982. V. 69. P. 869. https://doi.org/10.1104/pp.69.4.869
Гаевский Н.А., Сорокина Г.А., Гольд В.М., Миролюбская И.В. Сезонные изменения фотосинтетического аппарата древесных и кустарниковых растений // Физиология растений. 1991. Т. 38. С. 685.
Шавнин С.А., Фомин А.С. Сезонные изменения флуоресценции хлорофилла хвои сосны обыкновенной // Физиология растений. 1993. Т. 40. С. 209.
Leverenz J.W. The effects of illumination sequence, CO2 concentration, temperature and acclimation on the convexity of the photosynthetic light response curve // Physiol. Plant. 1988. V. 74. P. 332. https://doi.org/10.1111/j.1399-3054.1988.tb00639.x
Huner N.P.A., Elfman B., Krol M., McIntosh A. Growth and development at cold-hardening temperatures. Chloroplast ultrastructure, pigment content, and composition // Can. J. Bot. 1984. V. 62. P. 53. https://doi.org/10.1139/b84-009
Strand M., Öquist G. Inhibition of photosynthesis by freezing temperatures and high light levels in cold-acclimated seedlings of Scots pine (Pinus Sylvestris L.). Effects on the light-limited and light-saturated rates of CO2 assimilation // Physiol. Plant. 1985. V. 64. P. 425. https://doi.org/10.1111/j.1399-3054.1985.tb08517.x
Бардин М.Ю., Ранькова Э.Я., Платова Т.В., Самохина О.Ф., Егоров В.И., Николаева А.М., Громов С.А., Алексеев Г.В., Радионов В.Ф., Александров Е.И., Дементьева Т.В., Коршунова Н.Н., Лавров А.С., Стерин А.М., Хохлова А.В. и др. Доклад об особенностях климата на территории Российской Федерации за 2018 год; http://www.meteorf.ru/upload/pdf_download/o-klimate-rf-2018.pdf
Shavnin S.A., Yusupov I.A., Montile A.A., Golikov D.Yu., Montile A.I. Effect of increased ambient temperature on the growth rate of young pine forests in the impact zone of a petroleum gas flare // Russian Journal of Ecology. 2009. V. 40. P. 1. https://doi.org/10.1134/S1067413609010019
Шавнин С.А., Юсупов И.А., Артемьева Е.П., Голиков Д.Ю. Влияние повышения температуры среды на формирование наземной растительности вблизи газового факела // Известия высших учебных заведений. Лесной журнал. 2006. № 1. С. 21.
Lichtenthaler H.K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes // Methods in enzymology / Ed. Colowick S.P., Kaplan N.O. San Diego: Academic Press, 1987. V. 148. P. 350. https://doi.org/10.1016/0076-6879(87)48036-1
Sudachkova N.E., Milyutina I.L., Romanova L.I. Adaptive responses of scots pine to the impact of adverse abiotic factors on the rhizosphere // Russian Journal of Ecology. 2009. V. 40. P. 387. https://doi.org/10.1134/S1067413609060022
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений