

Физиология растений, 2022, T. 69, № 6, стр. 589-596
Светорегуляция изоферментов цикла трикарбоновых кислот в растениях
Д. Н. Федорин a, А. Т. Епринцев a, *, А. У. Игамбердиев b
a Федеральное государственное бюджетное образовательное учреждение высшего образования
Воронежский государственный университет
Воронеж, Россия
b Мемориальный университет Ньюфаундленда
Ньюфаундленд, Канада
* E-mail: bc366@bio.vsu.ru
Поступила в редакцию 04.02.2022
После доработки 05.04.2022
Принята к публикации 25.04.2022
- EDN: TBWPDF
- DOI: 10.31857/S0015330322060045
Аннотация
Светорегуляция цикла трикарбоновых кислот (ЦТК) в зеленых листьях растений осуществляется активными формами фитохрома, криптохрома, а также другими механизмами, которые проявляют ингибирующее действие по отношению к активности ряда митохондриальных изоферментов. Трансдукция фитохромного сигнала в ядро клетки осуществляется путем перераспределения свободных катионов кальция между компартментами клетки, что приводит к активации кальмодулинов. Взаимодействуя с Са2+-СаМ-зависимой киназой, кальмодулины обеспечивают контроль за функционированием транскрипционного фактора PIF3, регулирующего экспрессию генов изоферментов 2-оксоглутаратдегидрогеназы, сукцинатдегидрогеназы, аконитатгидратазы, малатдегидрогеназы, фумаратгидратазы и цитратсинтазы в зеленых листьях растений, таким образом контролируя скорость функционирования ЦТК. Высокий метильный статус промоторов генов, кодирующих эти ферменты, приводит к подавлению их транскрипции, что предполагает эпигенетический механизм светорегуляции их экспрессии. Изменение метильного статуса отдельных CG-динуклеотидов в составе CpG-островков промоторов генов обеспечивает регуляцию их функционирования в листьях растений в условиях различного освещения.
ВЛИЯНИЕ СВЕТА НА ДЫХАТЕЛЬНЫЙ МЕТАБОЛИЗМ РАСТЕНИЙ
Влияние света на скорость дыхательного метаболизма растений является установленным фактом, что показано для некоторых ферментативных систем, таких как сукцинатдегидрогеназа (СДГ), комплекс I митохондриальной электрон-транспортной цепи (ЭТЦ), НАД-фосфоглицеральдегид дегидрогеназа [1–4]. Важное значение в контроле окислительного метаболизма клетки играют ферментные системы, в частности, локализованные в митохондриях. Показано, что светозависимость характерна для аконитатгидратазы (АГ), фумаратгидратазы (ФГ), цитратсинтазы (ЦС), малатдегидрогеназы (МДГ) [4–7]. Данные энзимы негативно регулируются светом, что проявляется в снижении их активности более чем в 2 раза по сравнению с условиями темноты. Снижение активности СДГ, МДГ, АГ, ФГ, ЦС на свету вызывает изменение скорости функционирования ЦТК как основного поставщика энергетических эквивалентов в клетки при дыхательном метаболизме. Это находит отражение и в изменении работы ЭТЦ митохондрий, поскольку свет тормозит работу комплекса I [1]. Однако в растительной клетке существуют системы, обеспечивающие несопряженное с запасанием энергии свободное окисление субстратов, такие как альтернативная цианидрезистентная оксидаза и ротенон-резистентные НАД(Ф)·Н-дегидрогеназы, позволяющее митохондриям эффективно регулировать энергетический статус клетки в условиях интенсивного фотосинтеза [8, 9].
В условиях функционирования фотосинтеза цикл Кребса не является основным источником энергии, а обеспечивает поставку углеродных скелетов и выступает в качестве регулятора окислительно-восстановительного гомеостаза клетки. Для ряда энзимов ЦТК характерно наличие генетически детерминированных изоферментов, имеющих внемитохондриальную локализацию в клетке растений. Регуляция их активности носит светозависимый характер [3, 7]. Увеличение каталитической активности цитоплазматических изоферментов ЦС и АГ на свету, вероятно, необходимо для активации работы фотосинтетического метаболизма, что обусловлено необходимостью увеличения скорости работы цикла Кальвина [3] за счет мобилизации пула запасных органических кислот клетки (в первую очередь цитрата, так как малат может мобилизовываться при помощи системы малик-энзимов), обеспечивая субстратами анаболические процессы. Очевидно, на свету ЦТК может функционировать не замкнуто и поставлять углеродные скелеты для биосинтетических процессов [10, 11]. Было высказано предположение, что цитрат является основным экспортируемым продуктом цикла Кребса в условиях фотосинтеза [12].
Активность ферментов ЦТК в условиях света значительно снижается по сравнению с таковыми показателями в темноте. При этом, уменьшение скорости функционирования характерно как для маркерного фермента цикла Кребса (СДГ) [4, 13], так и для митохондриальных изоферментов ЦС, АГ, ФГ [8, 14].
Изменение активности изоферментов СДГ, ЦС и АГ в листьях кукурузы при изменении светового режима растений свидетельствует об их светозависимости. Установлено, что в регуляции митохондриальных и внемитохондриальных изоферментов СДГ, ЦС, ФГ и АГ принимают участие фоторецепторные системы. Одним из способов такой регуляции является контроль уровня транскриптов генов в клетке [15].
ФИТОХРОМНАЯ РЕГУЛЯЦИЯ МИТОХОНДРИАЛЬНЫХ ФЕРМЕНТОВ
Фитохромная система может регулировать митохондриальное дыхание как посредством регуляции активности уже синтезированных ферментов, так и посредством модификации экспрессии их генов [3, 16]. Многие ферменты сопряженного митохондриального дыхания, а также первый комплекс ЭТЦ митохондрий снижают скорость своего функционирования в присутствии активной формы фитохрома в клетке [4, 13, 14], в то время как ферменты альтернативных путей электронного транспорта увеличивают свою активность [8, 9].
В реализации фитохромного сигнала чаще всего задействованы внутриклеточные и внутриядерные переносчики сигналов [17, 18]. Поскольку фитохром А имеет в большей степени цитозольную локализацию [19], а сигнал передается в ядро на молекулу ДНК, то возникает необходимость задействования клеточных интермедиатов в реализации фитохромного сигнала в пределах клетки. Один короткий импульс красного света индуцирует импорт фитохрома А в ядро, который происходит очень быстро (в течение нескольких минут), тогда как перенос фитохрома В в ядро относительно медленный и занимает часы [19]. При этом, в клетке имеется сигнальный механизм передачи фитохромного сигнала, где внутриклеточными посредниками являются Ca2+ и цАМФ [20–22].
Активная форма фитохрома вызывает увеличение содержания свободных катионов кальция в ядрах клеток, что наблюдается при облучении растений красным светом с длиной волны 660 нм. Изменение концентрации свободных ионов кальция в ядрах листьев кукурузы зависит от состояния фитохромной системы и может быть обусловлено его перераспределением между компартментами клетки [23, 24] или высвобождением из кальциевых депо [25].
Результаты исследований с применением специфического ингибитора кальциевых каналов (рутений красный) и комплексона (ЭГТА) свидетельствуют, что изменение содержания свободного кальция в ядрах клеток листьев кукурузы связано с его перераспределением между компартментами клетки, в частности, с переносом ионов Са2+ из цитоплазмы в ядро [26]. Увеличение концентрации кальция в ядрах в ответ на облучение растений КС, приводит к запуску каскадных механизмов фитохром-зависимой регуляции экспрессии генов посредством внутриядерных трансдукторов сигнала.
Установлено, что изменение уровня транскриптов генов СДГ [26–28], ФГ [29] и колебания концентрации катионов кальция в ядрах растений в условиях освещения светом разного спектрального состава находятся в определенной корреляционной зависимости. Вероятно, катионы кальция играют значительную роль в трансдукции фитохромного сигнала, модулируя различные механизмы регуляции экспрессии генетического материала клетки.
Фитохром-зависимое внутриклеточное перераспределение свободных катионов кальция вызывает изменения в функционировании Ca2+-зависимых протеинкиназы и фосфатазы, осуществляющих контроль взаимодействия ДНК и транскрипционных факторов и регулирующих экспрессию генов [26, 30].
Кальмодулины могут обеспечивать реализацию кальциевого и фосфатного путей передачи сигнала, а также участвуют в фосфорилировании [31]. Кроме того, кальмодулины проявляют свое действие через непосредственное воздействие на белковые молекулы [32].
Фитохром-зависимое увеличение концентрации кальция в ядерной фракции приводит к активации кальмодулина 7, проявляющего киназную реакцию и способного модулировать работу транскрипционных факторов. Активная форма фитохрома вызывает увеличение количества мРНК гена белка кальмодулина 7, в частности calm7-4, локализованного в 3 хромосоме. Посредником во внутриядерной передаче фитохромного сигнала является транскрипционный фактор PIF3, инактивация которого за счет фосфорилирования наблюдается в ответ на накопление активной формы фитохрома А в клетке (рис. 1).
Рис. 1.
Гипотетическая схема трансдукции фитохромного сигнала и механизма кальций-зависимой регуляции экспрессии генов при участии кальмодулина и транcкрипционного фактора PIF3, обеспечивающего изменения сродства РНК-полимеразы к промоторам генов, в листьях растений. Фк – фитохром красный, Фдк – фитохром дальний красный, Ф – фосфатная группа, PIF3 – фитохром-индуцируемый транскрипционный фактор.
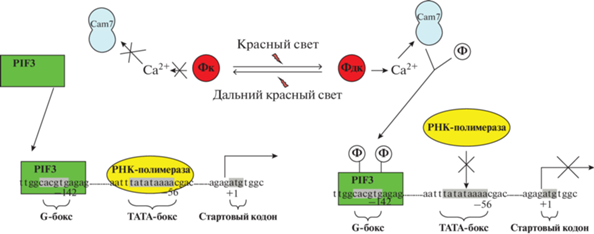
Наличие активной формы фитохрома в клетке способствует уменьшению экспрессии генов ФГ и СДГ в листьях кукурузы при их облучении красным светом (660 нм) [6, 33]. Кроме того, известно, что фитохром А играет доминирующую роль в регуляции деградации PIF1 на свету, в то время как фитохромы В и D также влияют на разрушение PIF1, но проявляют свое действие в условиях более длительного освещения [34].
В отличие от PIF3, транскрипционный фактор PIF1 не взаимодействует непосредственно с фитохромами А и В, однако имеет важное значение в регуляции метаболизма в темноте. Эти данные свидетельствуют о том, что регуляция световых процессов в клетках растений может осуществляться путем удаления негативных регуляторов (например, PIF) путем светозависимого протеолиза [35].
Важное значение в PIF-зависимой регуляции генов играют особенности структуры промоторов генов, в частности, специфических участков связывания (G- и Е-участки) для транскрипционного фактора семейства PIF [36–38]. Наличие данного участка в составе промотора исследуемых генов указывает на возможность регуляции их транскрипции на уровне изменения сродства РНК-полимеразы к промоторам генов (рис. 1) [39].
ФИТОХРОМ-ЗАВИСИМОЕ МЕТИЛИРОВАНИЕ ПРОМОТОРОВ ГЕНОВ МИТОХОНДРИАЛЬНЫХ ФЕРМЕНТОВ
Не все гены имеют в составе нуклеотидной последовательности промоторов CpG-островки, однако, они подвергаются фитохром-зависимой регуляции, следовательно, контроль за их экспрессией может осуществляться иными способами, в том числе эпигенетическим за счет изменения метильного статуса ДНК. Метилирование ДНК – один из факторов, влияющих на экспрессию генов. Изменение метильного статуса цитозина в растительных организмах в большей степени характерно для промоторных областей генов [40]. Ингибирование скорости транскрипции генов является результатом изменения состояния хроматина (увеличения его конденсации при увеличении метильного статуса), что препятствует присоединению транскрипционных факторов к соответствующему гену. При этом, изменение статуса метилирования одного нуклеотида практически не влияет на процесс присоединения транскрипционного фактора к промотору гена [41].
Значительные отличия в регуляции генов наблюдаются при отсутствии в составе промотора CpG-островка. Изменение метильного статуса отдельных цитозинов промотора данных генов оказывает значительное влияние на уровень его экспрессии. Подобный эффект был обнаружен для транскрипционного фактора AP-2 [42], участвующего в регуляции роста и развития растений, созревании плодов, защитной реакции и метаболизме [43].
Применение метода метилспецифичной ПЦР позволило установить роль метильного статуса промоторов генов ЦС и АГ, 2ОГДГ, СДГ, ФГ и МДГ в их регуляции. Как правило, метильный статус меняется в регуляторных областях генов (промоторах) или в транскрибируемых областях. При этом метильный статус отдельных нуклеотидов коррелирует с уровнем транскрипции соответствующих генов [44, 45]. Для генов флавопротеина CДГ [46], а также генов митохондриальных изоферментов АГ [7], ФГ и ЦС [14] показано, что увеличение метильного статуса их промоторов в условиях наличия активной формы фитохрома в клетке соотносится с уменьшением количества транскриптов. Невысокий метильный статус CG-динуклеотидов промоторов генов sdh1-2, aco1, fum1 и csy1 в растениях, формируемый в клетках растений в темноте и при действии ДКС, способствует увеличению содержания мРНК генов исследуемых изоферментов метаболизма ди- и трикарбоновых кислот и их каталитической активности (рис. 2). Противоположное действие вызывает красный свет, что приводит к увеличению статуса метилирования промоторов генов исследуемых изоферментов и угнетению их каталитической активности.
Рис. 2.
Схема внутриклеточной трансдукции фитохромного сигнала в растительной клетке и регуляции экспрессии генов митохондриальных изоферментов ЦТК. Фк – фитохром красный, Фдк – фитохром дальний красный, Ф – фосфатная группа, PIF3 – фитохром-индуцируемый транскрипционный фактор, СаМ – кальмодулин.
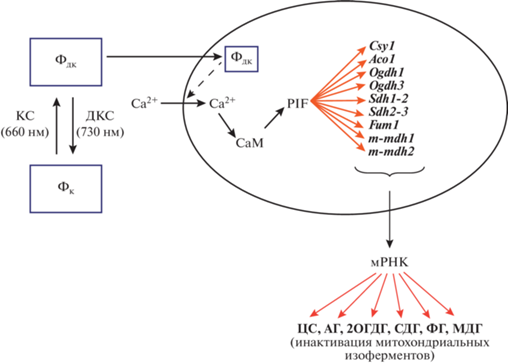
Для генов sdh1-2, aco1, fum1 и csy1 изоферментов СДГ, ЦС, ФГ и АГ показано наличие в составе их промоторов CpG-островков (табл. 1), что является необходимым условием их регуляции посредством изменения метильного статуса ДНК [47–49]. Отсутствие CpG-островков в промоторах генов sdh2-3, ogdh1 и ogdh2 митохондриальных изоферментов кукурузы свидетельствует об ином механизме контроля их экспрессии, не связанного с изменением метильного статуса регуляторной области соответствующих генов [39].
Таблица 1.
Распределение CpG-островков в составе промоторов генов ферментов цикла трикарбоновых кислот и их регуляция за счет изменения метильного статуса
| ген | Наличие CpG-островка | Регуляция за счет метилирования |
|---|---|---|
| Csy1 | 1 островок | Есть |
| Aco1 | 2 островка | Есть |
| Ogdh1 | Нет островка | Нет |
| Ogdh2 | Нет островка | Нет |
| Ogdh3 | 2 островка | Нет |
| Sdh1-2 | 2 островка | Есть |
| Sdh2-3 | Нет островков | Нет |
| Fum1 | 1 островок | Есть |
| m-mdh1 | 2 островка | Нет |
| m-mdh2 | 1 островок | Нет |
Изменение метильного статуса промоторов генов митохондриальных изоферментов играет важную роль в их фитохром-зависимой регуляции. Для генов изоферментов СДГ, АГ, ФГ и ЦС показано, что увеличение метильного статуса их промоторов соотносится с низким содержанием транскриптов (рис. 3). Невысокая степень метилирования CG-динуклеотидов промоторов генов sdh1-2, aco1 и csy1 в растениях, экспонируемых на свету и после облучения красным светом, совпадала с высоким уровнем экспрессии исследуемых генов и, как следствие, высокой каталитической активностью энзимов [7, 14, 45]. Стоит отметить, что регуляция экспрессии характерна для генов изоферментов ЦТК, в промоторе которых присутствует CpG-островок. Обратная ситуация наблюдается в темноте и при облучении растений дальним красным светом и последовательным облучением красным и дальним красным светом. В данных вариантах опыта наблюдается увеличение величины метильного статуса CG-динуклеотидов промоторов генов исследуемых ферментов, что приводит к снижению скорости их функционирования.
Рис. 3.
Гипотетическая схема эпигенетической регуляции экспрессии генов изоферментов ЦТК фитохромной системой, осуществляемая посредством кальций-зависимой регуляции активности ДНК-метилтрансферазы. Фк – фитохром красный, Фдк – фитохром дальний красный, Ф – фосфатная группа, СаМ – кальмодулин, SAM – S-аденозилметионин, SAH – S-аденозилгомоцистеин.

ЗАКЛЮЧЕНИЕ
Регуляция функционирования цикла трикарбоновых кислот в листьях растений в условиях светового режима обусловлена наличием в клетке митохондриальных и цитоплазматических изоферментов. Светозависимость активности 2-ОГДГ, АГ, ФГ, СДГ, ЦС и МДГ обусловлена длиной волны света, воздействующего на растения, и имеет дифференциальный характер по отношению к митохондриальным и цитоплазматическим изоферментам. Активная форма фитохрома А осуществляет основную, а фитохром В вспомогательную роль в регуляции функционирования изоферментов цикла Кребса, приводя к уменьшению уровня транскриптов митохондриальных форм ферментов и их активности и увеличению данных показателей для цитоплазматических изозимов. Активация фитохрома способствует увеличению содержания свободных катионов кальция в ядре посредством мембранных кальциевых каналов, обеспечивая, таким образом, трансдукцию сигнала в ядро клетки [50]. Кальциевая сигнализация опосредуется кальмодулинами, которые принимают непосредственное участие в световой регуляции клеточного метаболизма [51], обеспечивая реализацию фоторецепторного сигнала на уровне функционирования ферментов ЦТК. Факторы семейства PIF являются посредниками во внутриядерной передаче фитохромного сигнала, где особая роль отводится транскрипционному фактору PIF3, что соотносится с увеличением уровня экспрессии его гена на свету [26].
Важное значение в регуляции экспрессии генов отводится изменению метильного статуса ДНК за счет функционирования ДНК-метилтрансфераз [52, 53]. Активация кальмодулинов фитохромом А может способствовать увеличению активности ДНК-метилтрансфераз при их фосфорилировании, что напрямую отражается на величине метильного статуса ДНК. Для генов СДГ, ЦС, ФГ и АГ, содержащих в промоторах CpG-островки, установлена четкая зависимость между уровнем транскриптов и величиной статуса метилирования отдельных CG-динуклеотидов. Высокий метильный уровень промоторов генов митохондриальных форм исследуемых ферментов приводит к снижению содержания их транскриптов в растениях на свету и при облучении красным светом. При этом, для промоторов генов цитоплазматических изоферментов АГ и ЦС показана обратная зависимость.
Работа выполнена при поддержке Российского фонда фундаментальных исследований, проект № 20-04-00296.
Авторы заявляют об отсутствии конфликта интересов. Настоящая работа не содержит каких-либо исследований с участием людей и животных в качестве объектов исследования.
Список литературы
Escobar M.A., Franklin K.A., Svensson A.S., Salter M.G., Whitelam G.C., Rasmusson A.G. Light Regulation of the Arabidopsis Respiratory Chain. Multiple Discrete Photoreceptor Responses Contribute to Induction of Type II NAD(P)H Dehydrogenase Genes // Plant Physiol. 2004. V. 136. P. 2710. https://doi.org/10.1104/pp.104.046698
Lyubimov V.Yu., Kreslavskii V.D. Phytochrome B-dependent Regulation of Reductive Phase of Photosynthetic Carbon Assimilation // Rus. J. Plant Physiol. 2017. V. 64. P. 776. https://doi.org/10.1134/S1021443717040082
Igamberdiev A.U., Eprintsev A.T., Fedorin D.N., Popov V.N. Phytochrome-mediated regulation of plant respiration and photorespiration // Plant Cell Environ. 2014. V. 37. P. 290. https://doi.org/10.1111/pce.12155
Lyubimov V.Yu., Kreslavski V.D., Shmarev A.N. Photoregulation of the Cytoplasmic PGA Dehydrogenase Complex in Wheat Leaves // Rus. J. Plant Physiol. 2020. V. 67. P. 797. https://doi.org/10.1134/S102144372005009X
Popov V.N., Fedorin D.N., Eprintsev A.T. Light Regulation of Succinate Dehydrogenase Expression in Arabidopsis thaliana Leaves // Rus. J. Plant Physiol. 2007. V. 54. P. 360. https://doi.org/10.1134/S1021443707030107
Eprintsev A.T., Fedorin D.N., Sazonova O.V. Phytochrome-Dependent Regulation of Fumarate Hydratase Activity in Maize Green Leaves // Rus. J. Plant Physiol. 2015. V. 62. P. 441. https://doi.org/10.1134/S102144371504007X
Eprintsev A.T., Fedorin D.N., Cherkasskikh M.V., Igamberdiev A.U. Regulation of expression of the mitochondrial and cytosolic forms of aconitase in maize leaves via phytochrome // Plant Physiol. Biochem. 2020. V. 146. P. 157. https://doi.org/10.1016/j.plaphy.2019.11.018
Popov V.N. Possible role of free oxidation processes in the regulation of reactive oxygen species production in plant mitochondria // Biochem. Soc. Trans. 2003. V. 31. P. 1316. https://doi.org/10.1042/bst0311316
Мальцева Е.В., Шацких А.С., Епринцев А.Т., Попов В.Н. Экспрессионная регуляция путей несопряженного и разобщенного дыхания в митохондриях томата // Вестник ВГУ. Серия: химия. биология. Фармация. 2012. № 2. С. 165.
Igamberdiev A.U., Gardestrom P. Regulation of NAD- and NADP-dependent isocitrate dehydrogenases by reduction levels of pyridine nucleotides in mitochondria and cytosol of pea leaves // Biochim. Biophys. Acta, Bioenerg. 2003. V. 1606. P. 117. https://doi.org/10.1016/s0005-2728(03)00106-3
Fernie A.R., Carrari F., Sweetlove L.J. Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport // Curr. Opin. Plant Biol. 2004. V. 7. P. 254. https://doi.org/10.1016/j.pbi.2004.03.007
Hanning I., Heldt H.W. On the function of mitochondrial metabolism during photosynthesis in spinach leaves (Spinacia oleracea L.). Partitioning between respiration and export of redox equivalents and precursors for nitrate assimilation products // Plant Physiology. 1993. V. 103. P. 1147. https://doi.org/10.1104/pp.103.4.1147
Епринцев А.Т., Федорин Д.Н., Анохина Г.Б., Седых А.В. Молекулярно-биохимические аспекты световой регуляции 2-оксоглутаратдегидрогеназы в растениях // Физиология растений. 2020. Т. 67. С. 206. https://doi.org/10.31857/S0015330320010054
Eprintsev A.T., Fedorin D.N., Dobychina M.A., Igamberdiev A.U. Regulation of expression of the mitochondrial and peroxisomal forms of citrate synthase in maize during germination and in response to light // Plant Sci. 2018. V. 272. P. 157. https://doi.org/10.1016/j.plantsci.2018.04.017
Quail P.H. Phytochome-interacting factors. In Light and Plant Development // Oxford, UK: Blackwell Publishing Ltd, 2007. P. 81. https://doi.org/10.1104/pp.17.01384
Kreslavski V.D., Carpentier R., Klimov V.V., Allakhverdiev S.I. Transduction mechanisms of photoreceptor signals in plant cells // J. Photochem. Photobiol., C. 2009. V. 10. P. 63. https://doi.org/10.1016/j.jphotochemrev.2009.04.001
Klose C., Viczian A., Kircher S., Schafer E., Nagy F. Molecular mechanisms for mediating light-dependent nucleo/cytoplasmic partitioning of phytochrome photoreceptors // New Phytol. 2015. V. 206. P. 965. https://doi.org/10.1111/nph.13207
Oh S., Montgomery B.L. Phytochrome-dependent coordinate control of distinct aspects of nuclear and plastid gene expression during anterograde signaling and photomorphogenesis // Front Plant Sci. 2014. V. 30: e171. https://doi.org/10.3389/fpls.2014.00171
Kircher S., Gil P., Kozma-Bognar L., Fejes E., Speth V., Husselstein-Muller T., Bauer D., Adam E., Schafer E., Nagy F. Nucleocytoplasmic partitioning of the plant photoreceptors phytochrome A, B, C, D, and E is regulated differentially by light and exhibits a diurnal rhythm // Plant Cell. 2002. V. 14. P. 1541. https://doi.org/10.1105/tpc.001156
Bowler C., Neuhaus G., Yamagata H., Chua N.H. Cyclic GMP and calcium mediate phytochrome phototransduction // Cell. 1994. V. 77. P. 73. https://doi.org/10.1016/0092-8674(94)90236-4
Neuhaus G., Bowler C., Hiratsuka K., Yamagata H., Chua N.H. Phytochrome-regulated repression of gene expression requires calcium and cGMP // EMBO J. 1997. V. 16. P. 2552. https://doi.org/10.1093/emboj/16.10.2554
Li J., Li G., Wang H., Denga X.W. Phytochrome signaling mechanisms // Arabidopsis Book. 2011. V. 9: e0148. https://doi.org/10.1199/tab.0148
Echevarria W., Leite M.F., Guerra M.T., Zipfel W.R., Nathanson M.H. Regulation of calcium signals in the nucleus by a nucleoplasmic reticulum // Nat. Cell Biol. 2003. V. 5. P. 440. https://doi.org/10.1038/ncb980
Leite M.F., Thrower E.C., Echevarria W., Koulen P., Hirata K., Bennett A.M., Ehrlich B.E., Nathanson M.H. Nuclear and cytosolic calcium are regulated independently // Proc. Natl. Acad. Sci. USA. 2003. V. 100. P. 2975. https://doi.org/10.1073/pnas.0536590100
Xiong T.C., Jauneau A., Ranjeva R., Mazars C. Isolated plant nuclei as mechanical and thermal sensors involved in calcium signaling // Plant J. 2004. V. 40. P. 12. https://doi.org/10.1111/j.1365-313X.2004.02184.x
Eprintsev A.T., Fedorin D.N., Igamberdiev A.U. Ca2+ is involved in phytochrome A-dependent regulation of the succinate dehydrogenase gene sdh1-2 in Arabidopsis // J. Plant Physiol. 2013. V. 170. P. 1349. https://doi.org/10.1016/j.jplph.2013.04.006
Eprintsev A.T., Selivanova N.V., Fedorin D.N., Bashkin S.S., Selezneva E.A., Dadakina I.V., Machmud Ali S. The Role of Calcium Cations in the Mechanism of Phytochrome-Dependent Regulation of the sdh1-2 Gene Expression and Succinate Dehydrogenase Activity in Maize Leaves // Biochemistry (Moscow), Supplement Series A: Membrane and Cell Biolog. 2012. V. 29. P. 310. https://doi.org/10.1134/S1990747812030051
Eprintsev A.T., Fedorin D.N., Karabutova L.A., Pokusina T.A. Light Regulation of Succinate Dehydrogenase Subunit B Gene SDH2-3 Expression in Maize Leaves // Rus. J. Plant Physiol. 2016. V. 63. P. 505. https://doi.org/10.1134/S102144371604004X
Eprintsev A.T., Fedorin D.N., Sazonova O.V., Igamberdiev A.U. Light inhibition of fumarase in Arabidopsis leaves is phytochrome A-dependent and mediated by calcium // Plant Physiology and Biochemistry. 2016. V. 102. P. 161. https://doi.org/10.1016/j.plaphy.2016.02.028
Galon Y., Finkler A., Fromm H. Calcium-regulated transcription in plants // Molecular Plant. 2010. V. 3. P. 653. https://doi.org/10.1093/mp/ssq019
Enslen H., Tokumitsu H., Soderling T.R. Phosphorylation of CREB by CaM-kinase IV activated by CaM-kinase IV kinase // Biochem. Biophys. Res. Commun. 1995. V. 207. P. 1038. https://doi.org/10.1006/bbrc.1995.1289
Crivici A., Ikura M. Molecular and structural basis of target recognition by calmodulin // Annu. Rev. Biophys. Biomol. Struct. 1995. V. 24. P. 85. https://doi.org/10.1146/annurev.bb.24.060195.000505
Popov V.N., Eprintsev A.T., Fedorin D.N., Igamberdiev A.U. Succinate dehydrogenase in Arabidopsis thaliana is regulated by light via phytochrome A // FEBS Letters. 2010. V. 584. P. 199. https://doi.org/10.1016/j.febslet.2009.11.057
Castillon A., Shen H., Huq E. Phytochrome interacting factors: central players in phytochrome-mediated light signaling networks // Trends Plant Sci. 2007. V. 12. P. 514. https://doi.org/10.1016/j.tplants.2007.10.001
Shen H., Zhu L., Castillon A., Majee M., Downie B., Huq E. Light-induced phosphorylation and degradation of the negative regulator PHYTOCHROME-INTERACTING FACTOR1 from Arabidopsis depend upon its direct physical interactions with photoactivated phytochromes // Plant Cell. 2008. V. 20. P. 1586. https://doi.org/10.1105/tpc.108.060020
Ezer D., Shepherd S.J.K., Brestovitsky A., Dickinson P., Cortijo S., Charoensawan V., Box M.S., Biswas S., Jaeger K.E., Wigge P.A. The G-box transcriptional regulatory code in Arabidopsis // Plant Physiol. 2017. V. 175. P. 628. https://doi.org/10.1104/pp.17.01086
Oh E., Kang H., Yamaguchi S., Park J., Lee D., Kamiya Y., Choi G. Genome-wide analysis of genes targeted by PHYTOCHROME INTERACTING FACTOR3-LIKE5 during seed germination in Arabidopsis // Plant Cell. 2009. V. 21. P. 403. https://doi.org/10.1105/tpc.108.064691
Martínez-García J.F., Huq E., Quail P.H. Direct targeting of light signals to a promoter element-bound transcription factor // Science. 2000. V. 288. P. 859. https://doi.org/10.1126/science.288.5467.859
Shin J., Park E., Choi G. PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis // Plant J. 2007. V. 49. P. 981. https://doi.org/10.1111/j.1365-313X.2006.03021.x
Antequera F., Bird A. Number of CpG islands and genes in human and mouse // Proc. Natl. Acad. Sci. USA. 1993. V. 90. P. 11995. https://doi.org/10.1073/pnas.90.24.11995
Kass S.U., Landsberger N., Wolffe A.P. DNA methylation directs a time-dependent repression of transcription initiation // Curr. Biol. 1997. V. 7. P. 157. https://doi.org/10.1016/s0960-9822(97)70086-1
Comb M., Goodman H.M. CpG methylation inhibits proenkephalin gene expression and binding of the transcription factor AP-2 // Nucleic Acids Res. 1990. V. 18. P. 3975. https://doi.org/10.1093/nar/18.13.3975
Gu C., Guo Z.-H., Hao P.-P., Wang G.-M., Jin Z.-M., Zhang S.-L. Multiple regulatory roles of AP2/ERF transcription factor in angiosperm // Bot. Stud. 2017. V. 58: 6. https://doi.org/10.1186/s40529-016-0159-1
Xu J., Zhou S., Gong X., Song Y., van Nocker S., Ma F., Guan Q. Single-base methylome analysis reveals dynamic epigenomic differences associated with water deficit in apple // Plant Biotechnol. J. 2018. V. 16. P. 672. https://doi.org/10.1111/pbi.12820
Liang L., Chang Y., Lu J., Wu X., Liu Q., Zhang W., Su X., Zhang B. Global methylomic and transcriptomic analyses reveal the broad participation of DNA methylation in daily gene expression regulation of Populus trichocarpa // Front. Plant Sci. 2019. V. 10. P. 243. https://doi.org/10.3389/fpls.2019.00243
Eprintsev A.T., Fedorin D.N., Karabutova L.A., Zenishcheva M.A., Igamberdiev A.U. Expression of the genes encoding A and B subunits of succinate dehydrogenase in maize is regulated via methylation of their promoters // Plant Biotech. Kingston, Canada: Queens University, 2016. P. 58.
Bewick A.J., Schmitz R.J. Gene body DNA methylation in plants // Curr. Opin. Plant Biol. 2017. V. 36. P. 103. https://doi.org/10.1016/j.pbi.2016.12.007
Teixeira F.K., Colot V. Gene body DNA methylation in plants: a means to an end or an end to a means? // EMB-O J. 2009. V. 28. P. 997. https://doi.org/10.1038/emboj.2009.87
Zhang H., Lang Z., Zhu J.K. Dynamics and function of DNA methylation in plants // Nat. Rev. Mol. Cell Biol. 2018. V. 19. P. 489. https://doi.org/10.1038/s41580-018-0016-z
Lau O.S., Deng X.W. Plant hormone signaling lightens up: integrators of light and hormones // Curr. Opin. Plant Biol. 2010. V. 13. P. 571. https://doi.org/10.1016/j.pbi.2010.07.001
Enslen H., Tokumitsu H., Soderling T.R. Phosphorylation of CREB by CaM-kinase IV activated by CaM-kinase IV kinase // Biochem. Biophys. Res. Commun. 1995. V. 207. P. 1038. https://doi.org/10.1006/bbrc.1995.1289
Zhang H., Lang Z., Zhu J.K. Dynamics and function of DNA methylation in plants // Nat. Rev. Mol. Cell Biol. 2018. V. 19. P. 489. https://doi.org/10.1038/s41580-018-0016-z
Finnegan E.J., Kovac K.A. DNA methyltransfereses // Plant Mol. Biol. 2000. V. 43. P. 189. https://doi.org/10.1023/A1006427226972
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений