

Физиология растений, 2023, T. 70, № 1, стр. 14-26
Фотосинтез: теория и эксперимент
В. И. Звалинский *
Тихоокеанский океанологический институт им. В. И. Ильичева Дальневосточного отделения
Российской академии наук
Владивосток, Россия
* E-mail: biomar@mail.ru
Поступила в редакцию 01.06.2022
После доработки 01.06.2022
Принята к публикации 02.06.2022
- EDN: AMHBSL
- DOI: 10.31857/S0015330322600395
Аннотация
Разработанный нами ранее подход для описания цепей сопряженных ферментативных реакций процесса фотосинтеза впервые позволил предложить строгую математическую модель непрямоугольной гиперболы, которая описывает зависимость скорости фотосинтеза от интенсивности света (I), концентрации CO2 и взаимодействие этих двух факторов. Фундаментальными параметрами этой модели являются световая (IK) и углекислотная (CK) субстратные константы, а также кривизна гиперболы (γ), параметры сопряжения (rIP и rCP) и максимальная скорость фотосинтеза (Pm). Предложены специальные спрямляющие координаты, в которых все семейство гипербол имеет вид прямой линии. Главные параметры могут быть найдены только при построении экспериментальных данных в спрямляющих координатах. Предлагаемая модель непрямоугольной гиперболы применима как для описания процесса фотосинтеза у морских водорослей и высших растений, так и для поглощения витамина B12 водорослями, и зависимости скорости выедания пищи зоопланктоном и рыбой.
ВВЕДЕНИЕ
Исследования световой кривой фотосинтеза (P–I зависимость) начались более ста лет назад. Первое количественное описание P–I зависимости было сделано в 1905 г. Blackman [1]. Его модель основана на “законе минимума” Либиха: прямолинейно восходящей части, заканчивающейся резким переломом, за которым следует горизонтальная часть (I – интенсивность света; Pm – максимальный фотосинтез; α – тангенс угла наклона):
(1a)
${\begin{gathered} P = \alpha I(0 < I < {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} \alpha }} \right. \kern-0em} \alpha }); \\ P = {{P}^{{\text{m}}}}(I > {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} \alpha }} \right. \kern-0em} \alpha }). \\ \end{gathered} }$Однако реальные кривые имеют более плавный переход от линейного участка к насыщению, по сравнению с ломаной линией Blackman. В 1935 г. Baly [2] использовал кинетику изолированного фермента в виде прямоугольной гиперболы [3], заменив концентрацию фермента на интенсивность света:
(1б)
${P = {{P}^{{\text{m}}}}\frac{I}{{{{I}_{{{1 \mathord{\left/ {\vphantom {1 2}} \right. \kern-0em} 2}}}} + I}}},$где P и Pm – текущий и максимальный фотосинтез, I и I1/2 – интенсивность света и константа полунасыщения.
Эта модель также неудовлетворительно описывала реальную P–I зависимость. В отличие от модели Blackman, модель Baly была “слишком плавной”, по сравнению с экспериментальными кривыми. Стало очевидным, что реальные световые кривые фотосинтеза находятся между ломаной Blackman и прямоугольной гиперболой Baly. Подробное исследование световой и углекислотной кривых фотосинтеза было проведено Рабиновичем [4]. Автор заключил, что зависимости P–I–CO2 должны быть представлены непрямоугольной гиперболой или кривыми более высокого порядка. Однако приемлемой математической формулировки этих зависимостей тогда предложено не было.
Позднее было предложено большое количество математических выражений для описания световой кривой фотосинтеза [5–8]. Все вновь предложенные функции располагались между ломаной Blackman и прямоугольной гиперболой Baly. Все модели, кроме моделей [1, 2], были эмпирическими. Проведенный анализ показывает, что в настоящее время для описания первичного продуцирования используются практически только эмпирические модели или модель [2].
Для характеристики световой (${P{\kern 1pt} - {\kern 1pt} I}$) или углекислотной (${P{\kern 1pt} - {\kern 1pt} {\text{C}}{{{\text{O}}}_{{\text{2}}}}}$) кривой фотосинтеза наиболее часто используются два параметра – тангенс угла наклона начального участка (α) и максимальный уровень фотосинтеза (Pm) [6, 9]. Угол α – эмпирический параметр, который не следует из теории кривой фотосинтеза. Иногда используется константа полунасыщения Михаэлиса-Ментен (I1/2) при P = 1/2Pm [3, 4]. Однако на сегодняшний день не определены параметры однозначно характеризующие кривые фотосинтеза.
Целью данной работы является проведение теоретического и экспериментального анализа модели непрямоугольной гиперболы процесса фотосинтеза в зависимости от двух факторов: интенсивности света и концентрации углекислоты, обращая особое внимание на взаимодействие этих двух факторов.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами для экспериментальных измерений служили оптически тонкие (D ≤ 0.2 единицы) образцы морских макроводорослей, а также суспензия одноклеточной водоросли Porphyridium cruentum и симбиотической микроводоросли Zooxanthellae. Скорость фотосинтеза измерялась по скорости выделения кислорода амперометрическим методом [10]. Измерения на Zooxanthellae проводились во время морских экспедиций на научно-исследовательском судне “Академик Александр Несмеянов”. Измерения на морских водорослях проводились на морской экспериментальной станции Института биологии моря Дальневосточного отделения РАН, расположенной на острове Попова в заливе Петра Великого. Данные по углекислотной зависимости процесса фотосинтеза были взяты из монографии Е. Рабиновича [4].
Математические расчеты и построения проводились с помощью программы Grapher.
РЕЗУЛЬТАТЫ
Основы подхода
Ранее нами был разработан подход для описания цепей сопряженных циклических реакций [8, 9, 11]. Была рассмотрена упрощенная модель процесса фотосинтеза в виде цепи от 3 до 10 сопряженных циклических реакций, которая позволяет получить математическое описание зависимости скорости процесса от интенсивности света, концентрации и сопротивления диффузии CO2. Предложены специальные спрямляющие координаты, в которых все семейство гипербол имеет вид прямой линии. С помощью этих координат установлено, что световая и углекислотная зависимости скорости фотосинтеза морских водорослей в первом приближении описываются уравнением непрямоугольной гиперболы.
Общая модель фотосинтеза
Общая модель фотосинтеза была взята из работы [12]. Упрощенная схема этого процесса может быть представлена в виде последовательной цепи сопряженных реакций. Поскольку последовательная цепь сопряженных реакций описывается цепной дробью [11], то схема на рис. 1 имеет решение в виде цепной дроби (поскольку цепная дробь имеет ограничение по высоте, дробь разделена на три слагаемых (а, б, в)):
(2)
${{\text{а}})\,\,1 - \frac{{{{r}_{{67}}}V}}{{1 - \frac{{{{r}_{{56}}}V}}{{1 - \frac{{{{r}_{{45}}}V}}{{1 - \frac{{{{r}_{{34}}}V}}{{1 - \frac{{{{r}_{{23}}}V}}{{1 - \frac{V}{{\beta I}} - \frac{{{{r}_{{12}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}} \right. \kern-0em} {{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}}}}}}}}}}}}\,\,\,\,{\text{б)}}\,\,1 - \frac{{{{r}_{{1213}}}V}}{{1 - \frac{{{{r}_{{1112}}}V}}{{1 - \frac{{{{r}_{{1011}}}V}}{{1 - \frac{{{{r}_{{910}}}V}}{{1 - \frac{V}{{\alpha I}} - \frac{{{{r}_{{89}}}V}}{{1 - \frac{{{{r}_{{78}}}V}}{{{{z}_{{67}}}}}}}}}}}}}}}}$Рис. 1.
Схема процесса фотосинтеза как полисубстратной цепи сопряженных циклических реакций. Представление взаимодействия компонентов через относительные сопротивления (rij): H2O/O2 – цикл разложения воды; ${\text{CO}}_{2}^{{\text{e}}}$ и ${\text{CO}}_{2}^{{\text{i}}}$ – концентрация углекислоты вне и внутри клетки; Z-Z+2, Q-Q+, … NADH-NADH+ … ATP-ADP – компоненты системы; PSI, PSII – фотосистемы I и II; αI и βI – доли света, поглощаемыми фотосистемами I и II; 13 – доставка энергии [I] в цикл Кальвина, 14 – цикл Кальвина; 1, 2 … 6, 7 … 16, 17 – порядковые номера взаимодействующих компонентов.
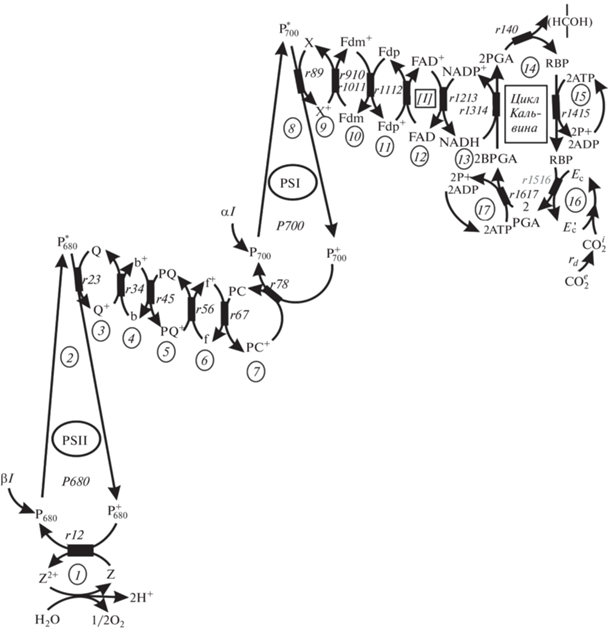
где V – относительная скорость процесса; rij – значения относительных сопротивлений, равные ${{{r}_{{ij}}} = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {({{k}_{{ij}}}[E_{i}^{0}][E_{j}^{0}])}}} \right. \kern-0em} {({{k}_{{ij}}}[E_{i}^{0}][E_{j}^{0}])}}}$, где kij – константа взаимодействия, $E_{i}^{0}$, $E_{j}^{0}$ – полная концентрация взаимодействующих компонентов, Pm – скорость цепи; αI, βI – относительные интенсивности света первой и второй систем; I – относительная интенсивность света, доставленная в цикл Кальвина; H2O – концентрация воды; Ce – относительная концентрация углерода вне клетки; rd – сопротивление диффузии.
Два первых слагаемых, (а) и (б), представляют решение для электрон-транспортной цепи (ЭТЦ), тогда как (в) – решение для цикла Кальвина. В цикле Кальвина (рис. 1) сопротивление r140 и компоненты “15” и “17” можно заменить эквивалентными сопротивлениями γ1 и γ2, соответственно, и их сумма (γ = γ1 + γ2 + r140) представлена в окончательном решении (г). Суммарная световая энергия, доставленная в цикл Кальвина, на рис. 1 представлена компонентом “13”, а в решении (г) − символом [I].
Цикл Кальвина
На рис. 2а отдельно представлен цикл Кальвина, что дает возможность подробно рассмотреть взаимодействие интенсивности света и концентрации углекислоты (CO2). CO2 включается в цикл восстановления (P) с помощью фермента EC. Сложный цикл восстановления углерода (P) представлен в виде одного “гиперцикла ключевого” фермента (компонента) EP. Поглощенные кванты света (I) включаются в гиперцикл через ферментативную циклическую систему компонента (EI), являясь источником энергии для функционирования цикла восстановления углерода (P). Каждое взаимодействие (реакция) сопровождается изменением состояния соответствующего фермента (рис. 2а).
Рис. 2.
Цикл Кальвина, описывающий взаимодействие интенсивности света и концентрации углекислоты: а – модель с абсолютными скоростями процесса – P “гиперцикл” восстановления углерода; б – та же модель с относительной скоростью (VP), относительной интенсивности света (I) и относительной концентрацией (С); в – модель зависимости от интенсивности света; г – модель зависимости от концентрации углекислоты. EP, EP’, EP” – различные состояния “ключевого” фермента (компонента); kIP, kCP – реакции второго порядка взаимодействия ферментативной циклической системы “I” (компонент EI) и циклической системы “C” (компонент EC); kP – реакция выделения продукта; [I] – интенсивность света; [Ce] и [Ci] – внешняя и внутренняя концентрации СО2; EI, EI’, EC, EC’ – различные состояния компонентов EI и EC; kI и kC – константы скорости включения субстратов I и C; kD – коэффициент диффузии СО2; rIP, rCP, rP, rd – относительные сопротивления.
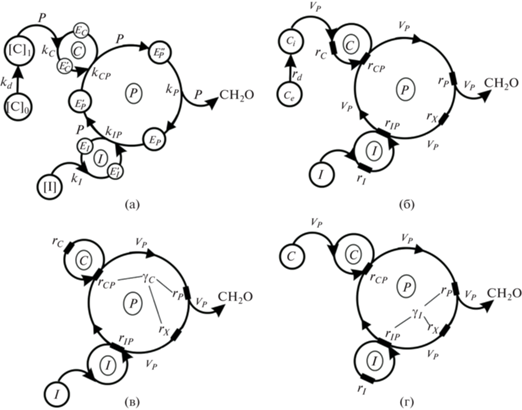
В стационарном состоянии, когда скорости взаимодействий (P) во всех звеньях равны между собой, схема представляется системой уравнений (рис. 3):
(3)
$\begin{gathered} 1.\,\,P = {{k}_{{\text{I}}}}[I][{{E}_{{\text{I}}}}]; \hfill \\ 2.\,\,P = {{k}_{{{\text{IP}}}}}[E_{{\text{I}}}^{'}][E_{{\text{P}}}^{'}]; \hfill \\ 3.\,\,P = {{k}_{{\text{d}}}}([{{C}_{{\text{e}}}}] - [{{C}_{i}}]); \hfill \\ 4.\,\,P = {{k}_{{\text{C}}}}[{{C}_{i}}][{{E}_{{\text{C}}}}]; \hfill \\ 5.\,\,P = {{k}_{{{\text{CP}}}}}[E_{{\text{C}}}^{'}][{{E}_{{\text{P}}}}]; \hfill \\ 6.\,\,P = {{k}_{{\text{P}}}}[E_{{\text{P}}}^{{''}}], \hfill \\ \end{gathered} $Рис. 3.
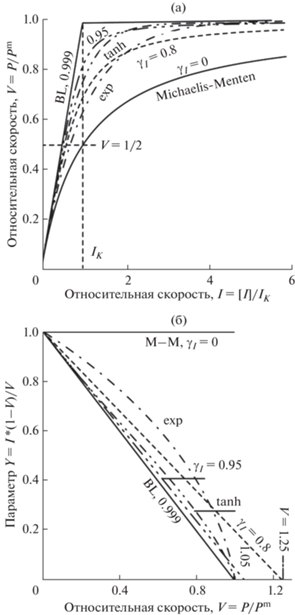
Световые (I – P) кривые фотосинтеза, полученные из уравнений (11), (13) и (14) в обычных (а) и спрямляющих (б) координатах. Значения параметра непрямоугольности гиперболы γI = 0.0 (M-M, прямоугольная гипербола Baly); 0.5, 0.9 (цифры на кривых) и 0.9999 (BL – ломаная Blackman). Для сравнения приведены функции экспоненты (exp) и гиперболического тангенса (tanh).
где P – скорость процесса; [I] – интенсивность света, доставленного в цикл Кальвина; [Ce] и [Ci] – внешняя и внутренняя (внутри клетки) концентрации углекислоты; [Ei] – концентрации различных форм ферментов (компонентов); kij – константы скорости процессов; kd – коэффициент диффузии.
Система (3) в своем составе имеет 10 переменных, большую часть из которых невозможно определить или измерить. По этой причине систему (3) в таком виде решить невозможно. Необходимо исключить не измеряемые величины и объединить в измеряемые. Если в стационарном состоянии суммарная концентрация всех форм каждого компонента $[E_{i}^{0}]$ остается неизменной, а сумма относительных концентраций равна “1” – ${{{[{{E}_{i}}]} \mathord{\left/ {\vphantom {{[{{E}_{i}}]} {[E_{i}^{0}]}}} \right. \kern-0em} {[E_{i}^{0}]}} + {{[E_{i}^{'}]} \mathord{\left/ {\vphantom {{[E_{i}^{'}]} {[E_{i}^{0}]}}} \right. \kern-0em} {[E_{i}^{0}]}} = {{e}_{i}} + e_{i}^{'} = 1}$, тогда:
(4)
${\begin{gathered} [E_{{\text{I}}}^{0}] = [{{E}_{{\text{I}}}}] + [E_{{\text{I}}}^{'}], \hfill \\ [E_{{\text{C}}}^{0}] = [{{E}_{{\text{C}}}}] + [E_{{\text{C}}}^{'}], \hfill \\ [E_{{\text{P}}}^{0}] = [{{E}_{{\text{P}}}}] + [E_{{\text{P}}}^{'}] + [E_{{\text{P}}}^{{''}}] \hfill \\ \end{gathered} }\,\,\,\,{\text{или}}\,\,\,\,{\begin{gathered} 1 = {{e}_{{\text{I}}}} + e_{{\text{I}}}^{'}; \hfill \\ 1 = {{e}_{{\text{C}}}} + e_{{\text{C}}}^{'}; \hfill \\ 1 = {{e}_{{\text{P}}}} + e_{{\text{P}}}^{'} + e_{{\text{P}}}^{{ }}. \hfill \\ \end{gathered} }$Из первого и второго уравнений системы (3) исключается величина [Ci] и с учетом уравнений (4) (${{{e}_{{\text{C}}}} = {{[{{E}_{{\text{C}}}}]} \mathord{\left/ {\vphantom {{[{{E}_{{\text{C}}}}]} {[E_{{\text{C}}}^{0}]}}} \right. \kern-0em} {[E_{{\text{C}}}^{0}]}}}$) получим:
(5)
$\begin{gathered} P = \left( {\left\{ {{{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}][{{C}_{e}}]} \right\} - \frac{{{{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}]}}{{{{k}_{{\text{d}}}}}}P} \right){{e}_{{\text{C}}}} \\ = \left( {\left\{ {{{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}][{{C}_{e}}]} \right\} - {{r}_{{\text{d}}}}P} \right){{e}_{{\text{C}}}}. \\ \end{gathered} $Как видно, полный поток CO2 при фотосинтезе состоит из внешнего потока углерода ${{{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}][{{C}_{{\text{e}}}}]}$ за вычетом поправки на диффузию rdP.
Анализ показывает, что система (3) содержит две задачи: 1 – задача о максимальной величине фотосинтеза (Pm); 2 – о воздействии внутренних и внешних факторов на параметры системы (по нашему мнению, именно эта проблема ранее не позволила решить задачу о световой кривой). Величина Pm – параметр, который характеризует систему в целом и служит своеобразным эталоном, с которым можно сравнивать другие параметры системы. Величину максимальной скорости (Pm) невозможно однозначно выразить через внутренние параметры, ее в каждом конкретном случае можно только измерить. Поэтому в дальнейшем величину Pm необходимо исключить из рассмотрения в системе (3).
Поделив обе части уравнений системы (3) на величину Pm и обозначив V = P/Pm с учетом уравнений (4) и (5), система (3) примет вид (рис. 2б):
(6)
${\begin{gathered} 1.\,\,V = \left\{ {(k{}_{{\text{I}}}[E_{{\text{I}}}^{0}]){{P}^{{\text{m}}}}} \right\}[I]{{e}_{{\text{I}}}}; \hfill \\ 2.\,\,V = \left\{ {{{({{k}_{{{\text{IP}}}}}[E_{{\text{I}}}^{0}][E_{{\text{P}}}^{0}])} \mathord{\left/ {\vphantom {{({{k}_{{{\text{IP}}}}}[E_{{\text{I}}}^{0}][E_{{\text{P}}}^{0}])} {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}} \right\}(1 - {{e}_{{\text{I}}}})e_{{\text{P}}}^{'}; \hfill \\ 3.\,\,V = \left( {\left\{ {{{({{k}_{{\text{C}}}}[E_{{\text{С}}}^{0}])} \mathord{\left/ {\vphantom {{({{k}_{{\text{C}}}}[E_{{\text{С}}}^{0}])} {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}} \right\}[{{C}_{{\text{e}}}}] - {{r}_{{\text{d}}}}V} \right){{e}_{{\text{C}}}}; \hfill \\ 4.\,\,V = \left\{ {{{\left( {{{k}_{{{\text{CP}}}}}[E_{{\text{C}}}^{0}][E_{{\text{P}}}^{0}]} \right)} \mathord{\left/ {\vphantom {{\left( {{{k}_{{{\text{CP}}}}}[E_{{\text{C}}}^{0}][E_{{\text{P}}}^{0}]} \right)} {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}} \right\}(1 - {{e}_{{\text{C}}}}){{e}_{{\text{P}}}}; \hfill \\ 5.\,\,V = \left\{ {{{\left( {k{}_{{\text{P}}}[E_{{\text{P}}}^{0}]} \right)} \mathord{\left/ {\vphantom {{\left( {k{}_{{\text{P}}}[E_{{\text{P}}}^{0}]} \right)} {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}} \right\}e_{{\text{P}}}^{{''}}. \hfill \\ \end{gathered} }$Именно эта операция – переход от абсолютной скорости (Pm) к относительной (V) позволил перейти от абсолютных не измеряемых величин к относительным измеряемым величинам: в системе уравнений (6) потоки можно сопоставить с главным потоком Pm.
В уравнениях 1 и 3 системы (6) имеются два однотипных параметра: ${{{I}_{{\text{K}}}} = \left\{ {{{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {(k{}_{{\text{I}}}[E_{{\text{I}}}^{0}])}}} \right. \kern-0em} {(k{}_{{\text{I}}}[E_{{\text{I}}}^{0}])}}} \right\}}$ и ${{{С}_{{\text{K}}}} = \left\{ {{{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {({{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}])}}} \right. \kern-0em} {({{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}])}}} \right\}}$. Эти параметры имеют размерность концентрации и характеризуют максимально возможное усвоение световой энергии и, соответственно ей, максимально возможную скорость утилизации углекислоты. В уравнениях 3 и 5 системы (6) имеется два однотипных безразмерных параметра: ${{{r}_{{{\text{IP}}}}} = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {({{k}_{{{\text{IP}}}}}[E_{{\text{I}}}^{0}][E_{{\text{P}}}^{0}])}}} \right. \kern-0em} {({{k}_{{{\text{IP}}}}}[E_{{\text{I}}}^{0}][E_{{\text{P}}}^{0}])}}}$ и rCP = ${ = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {({{k}_{{{\text{CP}}}}}[E_{{\text{C}}}^{0}][E_{{\text{P}}}^{0}])}}} \right. \kern-0em} {({{k}_{{{\text{CP}}}}}[E_{{\text{C}}}^{0}][E_{{\text{P}}}^{0}])}}}$. Параметры rIP и rСP – относительные сопротивления (по отношению к V) реакций усвоение света и утилизации углекислоты. Они характеризуют пропускную способность реакций циклов сопряжения светового (EI) и углекислотного (EC) ферментов и цикла “ключевого” фермента EP. Выражение ${{{r}_{{\text{P}}}} = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {(k{}_{{\text{P}}}[E_{{\text{P}}}^{0}])}}} \right. \kern-0em} {(k{}_{{\text{P}}}[E_{{\text{P}}}^{0}])}}}$ определяет пропускную способность реакций выделения продукта.
Благодаря введению безразмерных кинетических параметров I = [I]/Ik и Ce = [Ce]/Ck и относительных сопротивлений звеньев rIP, rCP, rP оказалось возможным получить достаточно простые математические выражения для описания световой и углекислотной кривых скорости первичного продуцирования. Используя уравнения (4) и (5), систему (6) можно представить в следующем виде:
(7)
${\begin{gathered} V = I{{e}_{{\text{I}}}} \hfill \\ V = ({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V){{e}_{{\text{C}}}} \hfill \\ V = (1 - {{e}_{{\text{I}}}}){{e_{{\text{P}}}^{'}} \mathord{\left/ {\vphantom {{e_{{\text{P}}}^{'}} {{{r}_{{{\text{IP}}}}}}}} \right. \kern-0em} {{{r}_{{{\text{IP}}}}}}} \hfill \\ V = (1 - {{e}_{{\text{C}}}}){{{{e}_{{\text{P}}}}} \mathord{\left/ {\vphantom {{{{e}_{{\text{P}}}}} {{{r}_{{{\text{CP}}}}}}}} \right. \kern-0em} {{{r}_{{{\text{CP}}}}}}} \hfill \\ V = {{e_{{\text{P}}}^{{''}}} \mathord{\left/ {\vphantom {{e_{{\text{P}}}^{{''}}} {{{r}_{{\text{P}}}}}}} \right. \kern-0em} {{{r}_{{\text{P}}}}}} \hfill \\ \end{gathered} }\,\,\,\,{\text{или}}\,\,\,\,\begin{gathered} \begin{gathered} {{e}_{{\text{I}}}} = {V \mathord{\left/ {\vphantom {V I}} \right. \kern-0em} I}; \hfill \\ {{e}_{{\text{C}}}} = {V \mathord{\left/ {\vphantom {V {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}} \right. \kern-0em} {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}; \hfill \\ e_{{\text{P}}}^{'} = {{{{r}_{{{\text{IP}}}}}V} \mathord{\left/ {\vphantom {{{{r}_{{{\text{IP}}}}}V} {(1 - {{e}_{{\text{I}}}})}}} \right. \kern-0em} {(1 - {{e}_{{\text{I}}}})}} = {{{{r}_{{{\text{IP}}}}}V} \mathord{\left/ {\vphantom {{{{r}_{{{\text{IP}}}}}V} {(1 - {V \mathord{\left/ {\vphantom {V I}} \right. \kern-0em} I})}}} \right. \kern-0em} {(1 - {V \mathord{\left/ {\vphantom {V I}} \right. \kern-0em} I})}}; \hfill \\ {{e}_{{\text{P}}}} = {{{{r}_{{{\text{CP}}}}}V} \mathord{\left/ {\vphantom {{{{r}_{{{\text{CP}}}}}V} {(1 - {{e}_{{\text{C}}}})}}} \right. \kern-0em} {(1 - {{e}_{{\text{C}}}})}} = {{{{r}_{{{\text{CP}}}}}V} \mathord{\left/ {\vphantom {{{{r}_{{{\text{CP}}}}}V} {(1 - {V \mathord{\left/ {\vphantom {V {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}} \right. \kern-0em} {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}})}}} \right. \kern-0em} {(1 - {V \mathord{\left/ {\vphantom {V {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}} \right. \kern-0em} {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}})}}; \hfill \\ \end{gathered} \hfill \\ e_{{\text{P}}}^{{''}} = {{r}_{{\text{P}}}}V. \hfill \\ \end{gathered} $Суммируя три последних уравнения системы (7) в правой части и учитывая, что ${{e}_{{\text{P}}}} + e_{{\text{P}}}^{'} + e_{{\text{P}}}^{{''}} = 1$, получаем выражение:
(8)
${\frac{{{{r}_{{{\text{IP}}}}}V}}{{{\text{1}} - {V \mathord{\left/ {\vphantom {V I}} \right. \kern-0em} I}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}} \right. \kern-0em} {({{C}_{{\text{e}}}} - {{r}_{{\text{d}}}}V)}}}} + ({{r}_{{\text{P}}}} + {{r}_{X}})V = 1}.$Как указывалось выше, помимо реакций выделения продукта (rP), цикл P включает реакции цикла Кальвина (рис. 1 циклы (15) и (17); рис. 2: γ = rP + rx). Тогда уравнение (8) примет вид:
(9)
${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \frac{V}{{{{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}}}}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[{{C}_{{\text{e}}}}]} \mathord{\left/ {\vphantom {{[{{C}_{{\text{e}}}}]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}} - {{r}_{{\text{d}}}}V}}}} = 1 - \gamma V}.$Соотношение (9) является наиболее общим фундаментальным выражением для модели фотосинтеза, которое описывает совместное влияние интенсивности света и концентрации углекислоты с учетом ее диффузии на скорость процесса (рис. 2б). Уравнение (9) – соотношение шестой степени по отношению к скорости V. Поскольку в стационарном состоянии сопротивление диффузии невозможно отделить от других составляющих включения СО2, для простоты будем рассматривать (9) без учета диффузии (rd= 0):
(10)
${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \frac{V}{{{{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}}}}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}}}}}} = 1 - \gamma V}.$Из (9) и (10) ясно, что член ${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {}}} \right. \kern-0em} {}}({{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}})}}}$ отображает утилизацию энергии света, тогда как член ${\frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}})}}} \right. \kern-0em} {({{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}})}}}}}$ – усвоение углекислоты, а их взаимодействие выражается через общий параметр γ (рис. 2б). Следует отметить, что в уравнении (10) члены, отображающие утилизацию энергии и усвоение углекислоты симметричны по отношению к общему параметру γ. Соотношение (10) – уравнение пятой степени по отношению к скорости V и оно не может быть разрешено относительно “V”, а может быть представлено в параметрических координатах (11а) – X = [I]; (11б) – X = [C]:
(11a)
${\begin{gathered} {{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}} = \frac{V}{{1 - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \gamma V - \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}}}}}}}}}}; \hfill \\ Y = V, \hfill \\ \end{gathered} }$(11б)
${\begin{gathered} {{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}} = \frac{V}{{1 - \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \gamma V - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \frac{V}{{{{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}}}}}}}}}}; \hfill \\ Y = V. \hfill \\ \end{gathered} }$Как указывалось выше, параметры IK и CK служат показателями максимальной продуктивности данного растения: параметр IK отображает максимально возможную доставленную энергию в цикл Кальвина в единицу времени и одновременно с ней параметр CK – максимально возможное количество фиксированной углекислоты за тот же промежуток времени. Величины IK и CK, как и их отношение IK/СK, являются константами для каждого растения. В дальнейшем IK будем обозначать “световую субстратную константу” рассматриваемой системы, а CK – “углекислотную субстратную константу”. Интенсивность света и концентрацию углекислоты удобно выражать в единицах их субстратных констант: I = = [I]/IK и Ce = [Ce]/CK. Результаты расчетов даются в долях от IK или CK.
Однофакторные уравнения от [I] и [С]
Уравнения (11а) и (11б) можно “разложить” на два однофакторных уравнения для световой (12а) и углекислотной (12б) зависимости (рис. 2в и 2г). Соотношения (12а) и (12б) – уравнения третьего порядка относительно V. Их удобно разрешить по отношению к субстратам [I] или [C] и построить в параметрических координатах; световая кривая будет иметь вид (12а), а углекислотная – (12б):
(12)
${\text{а}})\,\,~{\begin{gathered} X = [I] = {{I}_{{\text{K}}}}\frac{V}{{1 - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {{\gamma }_{{\text{I}}}}V}}}} \hfill \\ Y = V \hfill \\ \end{gathered} };\,\,\,\,{\text{б}})\,\,{\begin{gathered} X = [С] = {{C}_{{\text{K}}}}\frac{V}{{1 - \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - {{\gamma }_{{\text{C}}}}V}}}}; \hfill \\ Y = V \hfill \\ \end{gathered} }$Здесь учтено, что ${{{\gamma }_{{\text{I}}}} = {{r}_{{\text{P}}}} + {{r}_{X}} + {{r}_{{{\text{CP}}}}} = \gamma + {{r}_{{{\text{CP}}}}}}$ (рис. 2в) и ${{{\gamma }_{{\text{C}}}} = \gamma + {{r}_{{{\text{IP}}}}}}$ (рис. 2г).
Однако, чтобы написать математическое выражение или построить график для конкретного образца, необходимо найти все параметры уравнения (12а) (γI, rIP, IK) и (12б) (γC, rCP, CK). Ниже описаны подходы, на основании которых можно найти эти параметры. Уравнения (9)–(12) удалось получить благодаря разделению задачи на две – о максимальной скорости (Pm), и о воздействии внутренних и внешних факторов на эту скорость. Величина (Pm) входит во все параметры модели: относительной скорости ${V = {P \mathord{\left/ {\vphantom {P {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}}$, субстратных констант ${{{С}_{{\text{K}}}} = \left\{ {{{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {({{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}])}}} \right. \kern-0em} {({{k}_{{\text{C}}}}[E_{{\text{C}}}^{0}])}}} \right\}}$, ${{{I}_{{\text{K}}}} = \left\{ {{{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {(k{}_{I}[E_{{\text{I}}}^{0}])}}} \right. \kern-0em} {(k{}_{I}[E_{{\text{I}}}^{0}])}}} \right\}}$, сопротивления rIP,rCP. Именно поэтому невозможно выразить Pm через внутренние параметры системы.
Уравнения второго порядка
Уравнения (12) можно упростить до кривых второго порядка (имея в виду, что ${{{\gamma }_{{\text{I}}}} + {{r}_{{{\text{IP}}}}} = 1}$ или ${{{\gamma }_{{\text{C}}}} + {{r}_{{{\text{CP}}}}} = 1}$). Так, для световой кривой (12а) получаем:
Таким образом, световая и углекислотная кривые будут выглядеть как (13а) и (13б) (в правых уравнениях I = [I]/IK и C = [C]/CK):
(13a)
$\begin{gathered} {[I] = {{I}_{{\text{K}}}}\frac{V}{{1 - V}}(1 - {{\gamma }_{{\text{I}}}}V)}\,\,\,\,{\text{или}} \\ {V = \frac{{1 - I}}{{2{{\gamma }_{{\text{I}}}}}}\left\{ {1 - \sqrt {1 - \frac{{4{{\gamma }_{{\text{I}}}}I}}{{{{{(1 - I)}}^{2}}}}} } \right\},} \\ \end{gathered} $(13б)
$\begin{gathered} {[C] = {{C}_{{\text{K}}}}\frac{V}{{1 - V}}(1 - {{\gamma }_{{\text{C}}}}V)}\,\,\,\,{\text{или}} \\ {V = \frac{{1 - I}}{{2{{\gamma }_{{\text{C}}}}}}\left\{ {1 - \sqrt {1 - \frac{{4{{\gamma }_{{\text{C}}}}C}}{{{{{(1 - C)}}^{2}}}}} } \right\}.} \\ \end{gathered} $Уравнения (13) незначительно отличаются от (12) – полного описания световых и углекислотных кривых. В силу своей простоты, их целесообразно использовать при моделировании растительных экосистем, которые на порядок точнее описывают процессы, по сравнению с прямоугольной гиперболой Михаэлиса–Ментен (уравнение Baly [2]).
Спрямляющие координаты для семейства гипербол
Уже в работе [4] отмечалось, что световые и углекислотные кривые являются непрямоугольными гиперболами или кривыми более высокого порядка. Мы не нашли работ, где с должной определенностью проверялось бы соответствие этих кривых гиперболам.
Нами в 1986 г. были предложены спрямляющие координаты для семейства гипербол, в которых можно однозначно отличить гиперболу от других похожих кривых [11]. Если соотношения (13а) и (13б) переписать в виде:
(14a)
${X = V;\,\,\,\,Y = [I]\frac{{1 - V}}{V} = {{I}_{{\text{K}}}}(1 - {{\gamma }_{{\text{I}}}}V)},$то при построении (14) в координатах ${Y = \left( {[I]\frac{{1 - V}}{V}} \right)}$ от X = V → Сmax (или ${Y = \left( {[С]\frac{{1 - V}}{V}} \right)}$ от X = V → Imax) эти соотношения будут иметь вид прямой с углом наклона – γI (или – γС, последнее на рисунке не представлено) (рис. 3). Так, при γI = 0 уравнение (14а) превращается в уравнение прямоугольной гиперболы [2, 3]: ${I = \frac{V}{{{\text{1}} - V}}}$ или ${V = \frac{I}{{{\text{1}} + I}}}$. При увеличении параметра γI → 1 (γI = = 0.9999) оно переходит в ломаную Blackman [1]. Таким образом, уравнение (14а) описывает кривые самой различной кривизны: от прямоугольной гиперболы Baly до ломаной Blackman (рис. 3). В обычных координатах все кривые имеют один и тот же начальный угол наклона и максимальный уровень насыщения, тогда как представленные ${V - I}$ кривые имеют одну и ту же интенсивность света насыщения, равную субстратной константе – [I] = IK (рис. 3).
Важно отметить, что световая константа (IK) и параметр кривизны (γI) являются фундаментальными параметрами для модели непрямоугольной гиперболы световой кривой. В соответствии с этой моделью параметр γI может принимать любую величину от 0 до 1 и описать световую кривую любой кривизны. На рис. 3 приведены функции $V = \exp (I)$ и $V = \tanh (I)$, которые, по словам авторов, наилучшим образом описывают экспериментальные световые кривые [6]. Однако эти кривые не следуют из теории и не спрямляются в предложенных координатах (рис. 3б, кривые “exp” и “tanh”). Соотношения (14а) и (14б) являются моделями “полностью” непрямоугольных гипербол.
Соотношения (13а) для световой кривой могут быть представлены (преобразованы) в канонической форме: где X = I и ${Y = V;\,\,\gamma \equiv {{\gamma }_{{\text{I}}}} \equiv {{\gamma }_{{\text{C}}}};}$ a и b – полуоси:
Это доказывает, что соотношения (13) и (14) являются непрямоугольными гиперболами.
Экспериментальные световые и углекислотные кривые и их параметры
Как следует из моделей (13) и (14), для того чтобы определить параметры экспериментальной кривой недостаточно построить кривую только в “обычных” координатах ${P \to I}$ (или ${P \to C}$). В таких координатах невозможно найти главные параметры модели: субстратные константы IK (CK) и кривизну непрямоугольной гиперболы γI (γC). Для того чтобы определить эти параметры, необходимо полученные экспериментальные точки скорости фотосинтеза построить в спрямляющих координатах: данные измерений пересчитать в относительную скорость ${X = V = {P \mathord{\left/ {\vphantom {P {{{P}^{{\text{m}}}}}}} \right. \kern-0em} {{{P}^{{\text{m}}}}}}}$ и параметр ${Y = {{[I](1 - V)} \mathord{\left/ {\vphantom {{[I](1 - V)} V}} \right. \kern-0em} V}}$ или ${Y = [С]{{(1 - V)} \mathord{\left/ {\vphantom {{(1 - V)} V}} \right. \kern-0em} V}}$, и полученные результаты построить в координатах Y от X (рис. 4б, г).
Рис. 4.
Экспериментальные световые и углекислотные кривые процесса фотосинтеза: а, б – световые кривые микроводоросли Porphyridium cruentum; в, г – световые кривые макроводоросли Porphyra sp.; д, е – углекислотная кривая водного растения Cabomba caroliniana, построенных в обычных (а, в, д) и спрямляющих (б, г, е) координатах; д, е – данные [10].
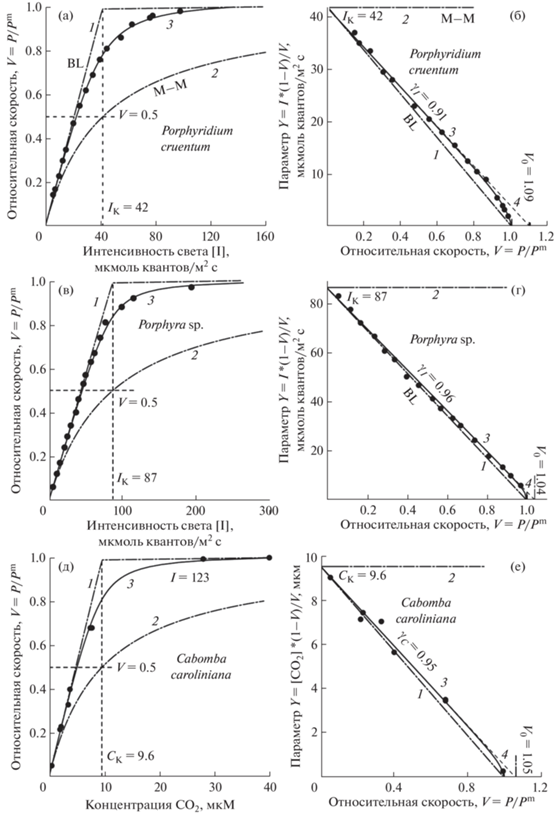
После этого через все экспериментальные точки в спрямляющих координатах в интервале ${X = 0{\kern 1pt} - {\kern 1pt} 0.9}$ проводится прямая (гипербола) ${Y = {{Y}_{0}}(AX + B)}$ до пересечения с осью ${X = V}$, где ${{{Y}_{0}} = {{I}_{{\text{K}}}}}$ – световой субстратный параметр (рис. 4б, г), а Y0 = CK – углекислотный субстратный параметр (рис. 4е; прямая “4”). Прямая Y0 → X0 соответствует непрямоугольной гиперболы с тангенсом угла наклона ${{{X}_{0}} = {{V}_{0}} = {{ - B} \mathord{\left/ {\vphantom {{ - B} A}} \right. \kern-0em} A} = {1 \mathord{\left/ {\vphantom {1 {{{\gamma }_{{\text{I}}}}}}} \right. \kern-0em} {{{\gamma }_{{\text{I}}}}}}}$ или 1/γC, отсюда ${{{r}_{{{\text{IP}}}}} = 1 - {{\gamma }_{{\text{I}}}}}$ или ${{{r}_{{{\text{СP}}}}} = 1 - {{\gamma }_{{\text{С}}}}}$ (рис. 4б, г, е). Таким образом, определяется “кривизна” гиперболы (γI) и сопротивление сопряжения светового (rIP) IK или углекислотного (γC и rCP) CO2 цикла. Для ориентира проведены линии, соответствующие ломаной Blackman [1], ${Y = {{Y}_{0}}(1 - X)}$, и прямоугольной гиперболы Baly ${Y = {{Y}_{0}}}$ [2] (рис. 4б, г, е; прямые 1 и 2), служащие предельными границами изменения кривых.
Однако в спрямляющих координатах реальные световые и углекислотные кривые в интервале ${V \approx {\text{0}}{\text{.9}}{\kern 1pt} - {\kern 1pt} {\text{1}}{\text{.0}}}$ отклоняются от непрямоугольной гиперболы (от прямой 4, рис. 4б, 4г, 4е), достигая V = 1.0 при Y = 0. В спрямляющих координатах полные уравнения (11а, 11б) будут иметь вид:
(15)
$\begin{gathered} ~{a)\,\,\begin{gathered} Х = V \to {{С}_{{\max }}}; \hfill \\ Y = [I]\frac{{1 - V}}{V} = {{I}_{{\text{K}}}}\frac{{1 - V}}{{1 - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {{\gamma }_{{\text{I}}}}V}}}} \hfill \\ \end{gathered} }, \\ {\text{б}})\,\,{\begin{gathered} X = V \to {{I}_{{\max }}}; \hfill \\ Y = [С]\frac{{1 - V}}{V} = {{C}_{{\text{K}}}}\frac{{1 - V}}{{1 - \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - {{\gamma }_{{\text{C}}}}V}}}} \hfill \\ \end{gathered} }. \\ \end{gathered} $В уравнениях (15а) и (15б) координата V отображает Cmax и Imax, соответственно. Подставляя в (15) найденные данные rIP, γI (или rCP, γC), получаем уравнение кривой в спрямляющих координатах (рис. 4б, г, е; кривые 3). Члены ${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {{\gamma }_{{\text{I}}}}V}}}$ на световых и ${\frac{{{{r}_{{{\text{CP}}}}}V}}{{{\text{1}} - {{\gamma }_{{\text{C}}}}V}}}$ – на углекислотных кривых описывают отклонение этих кривых от непрямоугольной гиперболы (рис. 4, кривые 3 и 4).
Из рис. 4 видно, что в спрямляющих координатах гипербола (кривая 3) и реальная (кривая 4) отличаются весьма незначительно, а в “стандартных” координатах отличие будет еще меньше (на рисунке не показано). Это означает, что квадратное уравнение (13) является хорошим приближением для точного уравнения третьей степени (12).
Только после этого по соотношениям (12) строятся кривые в “стандартных” координатах – ${X = [I],\,\,Y = V}$ (рис. 4а, 4в, 4д, кривые 3). Без анализа графиков в спрямляющих координатах кривых фотосинтеза невозможно определить фотосинтетические параметры. Таким образом, устанавливается полное аналитическое уравнение конкретной экспериментальной кривой, фундаментальными параметрами которой являются: субстратные параметры – IK (СK), кривизна гиперболы – γI (γС), сопротивления сопряжения – rIP (rCP) (рис. 4б, в, е), при этом максимальная скорость фотосинтеза (Pm) измеряется отдельно.
Из измерений одной кривой можно определить только одно независимое сопротивление: γI – из световой и γС – из углекислотной кривой, а также одно зависимое – ${{{r}_{{{\text{IP}}}}} = 1 - {{\gamma }_{{\text{I}}}}}$ и ${{{r}_{{{\text{СP}}}}} = {\text{1}} - {{\gamma }_{{\text{C}}}}}$. Мы не нашли одновременных экспериментальных данных измерений световых и углекислотных кривых фотосинтеза, полученных на одном и том же образце. Но так как величины rIP и rCP на разных образцах имеют близкие значения (рис. 4б, г, е), то можно допустить, что для одного образца они имеют также близкие значения: rIP ≈ rCP.
Приведенные данные (рис. 4) показывают, что сопротивления сопряжения субстратных (светового и углекислотного) циклов более чем в 20 раз меньше, чем суммарное сопротивление цикла переработки P: rIP ≈ rCP ≈ 0.02–0.05 и γ ≈ 0.92–0.95.
Взаимодействие интенсивности света и концентрации углекислоты
Как указано выше, чтобы установить совместное влияние интенсивности света и концентрации углекислоты на скорость фотосинтеза, необходимо измерить световые и углекислотные на одном и том же образце и построив их в спрямляющих координатах, найти IK, СK, rIP,rCP, и γ. Подставляя эти данные в соотношение (10) или (11) получаем искомое уравнение.
На рис. 5 представлены световые кривые морской макроводоросли Ulva fenestrata и симбиотической водоросли Zooxanthellae (рис. 5а и 5в соответственно), измеренные при двух концентрациях СО2 – при равновесной с воздухом (${\text{CO}}_{2}^{{{\text{atm}}}}$) и при продувании воды воздухом, содержащим +1.0% − насыщение СО2 (${\text{CO}}_{2}^{{{\text{max}}}}$). При насыщении СО2 уровень фотосинтеза почти в 3 раза выше, чем при равновесной ${\text{CO}}_{2}^{{{\text{atm}}}}$ (для U. fenestrata – Vmax/Vatm = 2.7, а для Zooxanthellae – Vmax/Vatm = = 2.63) (рис. 5а, в).
Рис. 5.
Экспериментальные световые кривые процесса фотосинтеза Ulva fenestrata (а, б) и Zooxanthellae (в, г) в морской воде с равновесным содержанием CO2 с атмосферой и насыщенной воздухом с CO2: 1 – 1% CO2, 2 – CO2 атмосферы, 3 – ломаная Blackman (объяснение с тексте).
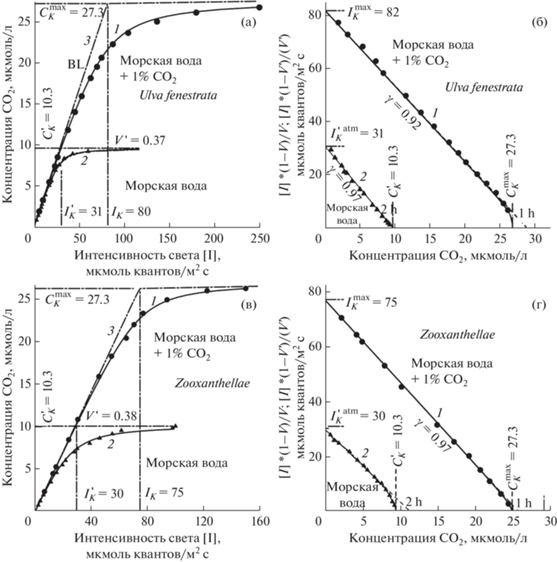
Световые кривые при насыщении ${\text{CO}}_{2}^{{{\text{max}}}}$, как при равновесии с воздухом (${\text{CO}}_{2}^{{{\text{atm}}}}$), в спрямляющих координатах также укладываются на прямую непрямоугольной гиперболы (рис. 5б, г, кривые 1h, 2h).
В спрямляющих координатах при насыщении ${\text{CO}}_{2}^{{{\text{max}}}}$ имеет более высокую $I_{{\text{K}}}^{{\max }}$, чем при ${\text{CO}}_{2}^{{{\text{atm}}}}$, при этом различие было пропорционально изменению уровня фотосинтеза: для U. fenestrata – ${{I_{{\text{K}}}^{{\max }}} \mathord{\left/ {\vphantom {{I_{{\text{K}}}^{{\max }}} {I_{{\text{K}}}^{{\max }}}}} \right. \kern-0em} {I_{{\text{K}}}^{{\max }}}}$ = 83/31 ≈ 2.68 и Vmax/Vatm = 2.7 для Zooxanthellae ${{I_{{\text{K}}}^{{\max }}} \mathord{\left/ {\vphantom {{I_{{\text{K}}}^{{\max }}} {I_{{\text{K}}}^{{\max }}}}} \right. \kern-0em} {I_{{\text{K}}}^{{\max }}}}$ = 75/29 ≈ 2.6 и Vmax/Vatm ≈ 2.61 (рис. 5б, г; кривые 1, 2).
Концентрация углекислоты в атмосфере (${\text{CO}}_{2}^{{{\text{atm}}}}$) ≈ ≈ 340 ррm или ≈13.9 мкмоль/л [13], а равновесная СО2 в воде (${\text{CO}}_{2}^{{{\text{atm}}}}$) ≈10.3 мкмоль/л [10]. Изменение уровня фотосинтеза происходит пропорционально концентрации СО2 (рис. 5б, г), т. е. ${{{С_{{\text{K}}}^{{\max }}} \mathord{\left/ {\vphantom {{С_{{\text{K}}}^{{\max }}} {С_{{\text{K}}}^{{{\text{atm}}}}}}} \right. \kern-0em} {С_{{\text{K}}}^{{{\text{atm}}}}}} \approx }$ ${ \approx \,\,{{{{V}^{{{\text{max}}}}}} \mathord{\left/ {\vphantom {{{{V}^{{{\text{max}}}}}} {{{V}^{{{\text{atm}}}}}}}} \right. \kern-0em} {{{V}^{{{\text{atm}}}}}}} \approx {\text{2}}{\text{.7}}}$, т.е. ${С_{K}^{{\max }} \approx {\text{27}}{\text{.5}}}$ мкмоль/л. Таким образом, как для U. fenestrata, так и для Zooxanthellae справедливо соотношение:
(16a)
${{{I_{{\text{K}}}^{{\max }}} \mathord{\left/ {\vphantom {{I_{{\text{K}}}^{{\max }}} {I_{{\text{K}}}^{{{\text{atm}}}}}}} \right. \kern-0em} {I_{{\text{K}}}^{{{\text{atm}}}}}} \approx {{С_{{\text{K}}}^{{\max }}} \mathord{\left/ {\vphantom {{С_{{\text{K}}}^{{\max }}} {С_{{\text{K}}}^{{{\text{atm}}}}}}} \right. \kern-0em} {С_{{\text{K}}}^{{{\text{atm}}}}}} \approx {{{{V}^{{{\text{max}}}}}} \mathord{\left/ {\vphantom {{{{V}^{{{\text{max}}}}}} {{{V}^{{{\text{atm}}}}}}}} \right. \kern-0em} {{{V}^{{{\text{atm}}}}}}}.}$Этого следовало ожидать, поскольку из общего уравнения (10) следует:
(16б)
${\frac{{{{C}_{{\text{K}}}}}}{{{{I}_{{\text{K}}}}}} = \frac{{[C]}}{{[I]}}}\,\,\,\,{\text{и}}\,\,\,\,{\frac{{[I]}}{{{{I}_{{\text{K}}}}}} = \frac{{[C]}}{{{{C}_{{\text{K}}}}}}}.$Учитывая изложенное выше, можно заключить, что полное представление о взаимодействии интенсивности света и углекислоты возможно только при совместном исследовании световых и углекислотных кривых фотосинтеза (рис. 5а, г). При равновесии с СО2atm измеряется световая кривая “обычных” и в спрямляющих координатах определяются параметры $I_{{\text{K}}}^{{{\text{atm}}}}$, $\gamma _{{\text{I}}}^{{{\text{atm}}}}$, $r_{{{\text{IP}}}}^{{{\text{atm}}}}$ (рис. 5б). Затем устанавливается интенсивность света [I] = $I_{{\text{K}}}^{{{\text{atm}}}}$, измеряется углекислотная кривая и также в спрямляющих координатах определяются параметры $C_{{\text{K}}}^{{{\text{atm}}}}$, $\gamma _{{\text{C}}}^{{{\text{atm}}}}$, $r_{{{\text{CP}}}}^{{{\text{atm}}}}$, (на рисунке не показано). На основе этих величин определяется полное представление о ${I \to P \to {\text{C}}{{{\text{O}}}_{2}}}$кривой образца.
Этот подход позволяет совершенно по-другому определять квантовый выход фотосинтеза (ϕm) (17а) и удельную молярную активность карбоксилирующего фермента (KC) (17б) [4, 12]:
(17)
${\text{а}})\,\,{{{\phi }_{{\text{m}}}} = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {a{{I}_{{\text{K}}}}}}} \right. \kern-0em} {a{{I}_{{\text{K}}}}}}};\,\,\,\,{\text{б}}){{{K}_{{\text{C}}}} = {{{{P}^{{\text{m}}}}} \mathord{\left/ {\vphantom {{{{P}^{{\text{m}}}}} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}},}$где a – поглощение света.
В этом случае полученные величины параметров ϕm и KC определяются на основе всего массива экспериментальных данных, и они будут точнее тех, которые определяются по начальному углу наклона световых или углекислотных кривых.
Таким образом, можно осуществить комплекс измерений при любой другой концентрации CO2, например, при насыщении ${\text{CO}}_{2}^{{{\text{max}}}}$ или концентрации CO2 < ${\text{CO}}_{2}^{{{\text{atm}}}}$. Как показано на рис. 5, при разных концентрациях CO2 параметры световых (γI, rIP) и углекислотных (γC, rCP) кривых даже для одного образца различаются. Такие исследования, на наш взгляд, могут дать ценную информацию о работе цикла восстановления углерода. Уравнения, которые помогут исследовать цикл Кальвина, приведены в таблице 1.
Таблица 1.
Список уравнений процесса фотосинтеза для различных случаев
| № п/п | Уравнения | Для каких случаев |
|---|---|---|
| 1. | ${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \frac{V}{{{{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}}}}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[{{C}_{{\text{e}}}}]} \mathord{\left/ {\vphantom {{[{{C}_{{\text{e}}}}]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}} - {{r}_{{\text{d}}}}V}}}} = 1 - \gamma V.}$ | 1. Фундаментальное выражение для модели процесса фотосинтеза с учетом диффузии (рис. 2б). 2. Без учета диффузии (рис. 2в, г). [I], IK – интенсивность света и его константа; [C], CK – концентрация CO2 и его константа |
| 2. | ${\frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \frac{V}{{{{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}}}}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}}}}}} = 1 - \gamma V}$ | |
| 3. | ${\begin{gathered} {{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}} = \frac{V}{{1 - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - \gamma V - \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - \frac{V}{{{{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}}}}}}}}}}; \hfill \\ Y = V \hfill \\ \end{gathered} }$ | То же самое, что и п. 2, в параметрических координатах |
| 4. | ${\begin{gathered} X = [I] = {{I}_{{\text{K}}}}\frac{V}{{1 - \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {{\gamma }_{{\text{I}}}}V}}}}; \hfill \\ Y = V \to {{C}_{{\max }}} \hfill \\ \end{gathered} }$ | Световая кривая в параметрических координатах (ось Y заменена на концентрацию CO2). См. рис. 5б, г |
| 5. | ${X = V \to {{С}_{{\max }}};}$ ${Y = [I]\frac{{1 - V}}{V} = {{I}_{{\text{K}}}}(1 - {{\gamma }_{{\text{I}}}}V)}$ |
Световая кривая в спрямляющих координатах (ось X заменена на концентрацию CO2). См. рис. 5б, г |
| 6. | ${[I] = {{I}_{{\text{K}}}}\frac{V}{{1 - V}}(1 - {{\gamma }_{{\text{I}}}}V)}$ или ${V = \frac{{{\text{1}} - I}}{{{\text{2}} \times {{\gamma }_{I}}}} \times \{ 1 - \sqrt {1 - \frac{{4 \times {{\gamma }_{I}} \times I}}{{{{{({\text{1}} - I)}}^{{\text{2}}}}}}} \} }$ |
Световая кривая: приближение уравнения второго порядка. – световая кривая относительно [I]. – cветовая кривая относительно V |
ОБСУЖДЕНИЕ
Разработанный ранее подход для описания цепей сопряженных ферментативных реакций [8, 9, 11] позволил разработать модель непрямоугольной гиперболы для процесса фотосинтеза. Математическая модель представляет цепную дробь, которая описывает зависимости скорости фотосинтеза от интенсивности света (I), концентрации CO2 и взаимодействие этих двух факторов. Предложены специальные спрямляющие координаты, в которых все семейство гипербол имеет вид прямой линии. Спрямляющие координаты позволили количественно определить световую (IK) или углекислотную (CK) константы и параметр непрямоугольности гиперболы γI или γC. Рассмотрение взаимодействия интенсивности света и концентрации СО2 позволило с максимальной точностью смоделировать экспериментальные кривые.
Следует особо отметить, что модель непрямоугольной гиперболы применима не только для описания процесса фотосинтеза морских водорослей (рис. 4 и 5), она имеет более общий характер. Кинетика процесса фотосинтеза высших растений также следует этой модели, с кривизной γ ≈ 0.9 [14]; зависимость скорости роста гетеротрофных организмов от концентрации пищи [15] также описывается этой кинетикой с кривизной γ ≈ 0.94. Зависимость скорости потребления клеток фитопланктона животными Daphnia и Calanus от их концентрации [16], скорости выедания пищи рыбой [17] показало, все эти процессы также протекают по кинетике непрямоугольной гиперболы с кривизной γ ≈ 0.95–0.98.
Фотосинтез растительной клетки – главный источник ее скорости роста. Помимо снабжения светом и углекислотой, растительная клетка должна снабжаться биогенными веществами в соответствии со своими потребностями, которая может быть выражена через их постоянную (константу) (N – соединения азота, P – соединения фосфора):
(19)
$\begin{gathered} \frac{{{{r}_{{{\text{IP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}})}}} \right. \kern-0em} {({{[I]} \mathord{\left/ {\vphantom {{[I]} {{{I}_{{\text{K}}}}}}} \right. \kern-0em} {{{I}_{{\text{K}}}}}})}}}} + \frac{{{{r}_{{{\text{CP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}})}}} \right. \kern-0em} {({{[C]} \mathord{\left/ {\vphantom {{[C]} {{{C}_{{\text{K}}}}}}} \right. \kern-0em} {{{C}_{{\text{K}}}}}})}}}} + \\ + \,\,\frac{{{{r}_{{{\text{NP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{[N]} \mathord{\left/ {\vphantom {{[N]} {{{N}_{{\text{K}}}}}}} \right. \kern-0em} {{{N}_{{\text{K}}}}}})}}} \right. \kern-0em} {({{[N]} \mathord{\left/ {\vphantom {{[N]} {{{N}_{{\text{K}}}}}}} \right. \kern-0em} {{{N}_{{\text{K}}}}}})}}}} + \frac{{{{r}_{{{\text{PP}}}}}V}}{{1 - {V \mathord{\left/ {\vphantom {V {({{[P]} \mathord{\left/ {\vphantom {{[P]} {{{P}_{{\text{K}}}}}}} \right. \kern-0em} {{{P}_{{\text{K}}}}}})}}} \right. \kern-0em} {({{[P]} \mathord{\left/ {\vphantom {{[P]} {{{P}_{{\text{K}}}}}}} \right. \kern-0em} {{{P}_{{\text{K}}}}}})}}}} + ... = 1 - \gamma V, \\ \end{gathered} $где NK и PK – субстратные константы соединения азота и фосфора соответственно и т.д.
Мы уже не раз обращали внимание на то, что как световые, так и углекислотные экспериментальные кривые (рис. 4а, в, д) резко отличаются от часто используемой в моделировании экосистем кривой прямоугольной гиперболы Михаэлиса-Ментен; по форме они скорее ближе к ломаной Блэкмана [18, 19].
Работа выполнена при частичной финансовой поддержке Российского фонда фундаментальных исследований, проекты №№ 15-35-50443 и 18-35-00543; приоритетной комплексной программы Российской академии наук “Дальний Восток”, гранты №№ 18-1-007 и 18-1-010; программы фундаментальных научных исследований государственных академий наук на 2021 – 2023 гг. (темы № 0211-2021-0008 “Исследование основных процессов, определяющих состояние и изменчивость океанологических характеристик окраинных морей Азии и прилегающих районов Тихого и Индийского океанов” и № 0211-2021-0014 “Эколого-биогеохимические процессы в морских экосистемах: роль природных и антропогенных факторов”).
Настоящая статья не содержит каких-либо исследований с участием людей и животных в качестве объектов исследования. Автор заявляет об отсутствии конфликта интересов.
Список литературы
Blackman F.F. Optima and limiting factors // Ann. Bot. 1905. V. 19. P. 281.
Baly E.C.C. The kinetics of photosynthesis // Proc. R. Soc. B. 1935. V. 117. P. 218.
Michaelis L., Menten M.L. Die kinetik der invertinwirkung // Biochem. Z. 1913. V. 49. P. 333.
Рабинович Е. Фотосинтез. Москва: Иностранная литература, 1953. Т. 2. 652 с.
Talling J.F. Photosynthetic characteristics of some freshwater plankton diatoms in relation to underwater radiation // New Phytol. 1957. V. 56. P. 29.
Jassby A.D., Platt T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton // Limnol. Oceanogr. 1976. V. 21. P. 540.
Platt T., Gallegos C., Harrison W.G. Photoingibition of photosynthesis in natural assemblages of marine phytoplankton // J. Mar. Res. 1980. V. 38. P. 687.
Звалинский В.И., Литвин Ф.Ф. Зависимость фотосинтеза от концентрации углекислоты, интенсивности и спектрального состава света // Физиология растений. 1988. Т. 35. С. 444.
Звалинский В.И. Формирование первичной продукции в море // Известия ТИНРО. 2006. Т. 147. С. 276.
Haxo F.T., Blinks L.R. Photosynthetic action spectra of marine algae // J. Gen. Physiol. 1960. V. 33. P. 389.
Звалинский В.И., Литвин Ф.Ф. Стационарная кинетика цепей сопряженных циклических реакций // Биохимия. 1986. Т. 51. С. 1741.
Гудвин Т., Мерсер Э. Введение в биохимию растений. Москва: Мир, 1986. Т. 1. 392 с.
NOAA Greenhouse Gas Marine Boundary Layer: ftp://ftp.cmdl.noaa.gov/ccg/co2/in-situ.
Liu J., Wang X., Rong Z., Gao Y., Zhang G., Wang W., Ge L., Mao Y., Guo Z., Wang Q., Zhao Ch. Modified non-rectangular hyperbola equation with plant height for photosynthetic light-response curves of Potentilla anserina and Elymus nutans at various growth phases in the Heihe River Basin, Northwest China // J. Arid Land. 2019. V. 11. P. 764.
Monod I. La technique de culture continue. Theoretic and application // Ann. Inst. Pasteur. Microbiol. 1950. V. 79. P. 390.
Shiotani T., Uye S. Selective feeding of the calanoid copepod Calanus sinicus on the natural microplankton assemblage, with special reference to microzooplankton // North Pacific Marine Organization (PICES) Ninth Annual Meeting Report (October 20–28, 2000). Hakodate, Hokkaido, Japan. 2000. P. 133.
Ивлев В.С. Экспериментальная экология питания рыб. Киев: Наукова думка. 1977. 272 с.
Kishi M.J., Kashiwai M., Ware D.M., Megrey B.A., Eslinger D.L., Werner F.E., Aita M.N., Azumaya T., Fujii M., Hashimoto Sh., Huang D., Iizumi H., Ishida Y., Kang S., Kantakov G.A. et al. NEMURO – a lower trophic level model for the North Pacific marine ecosystem // Ecol. Model. 2007. V. 202. P. 12.
Vichi M., Pinardi N., Masina S. A generalized model of pelagic biogeochemistry for the global ocean ecosystem. Part I: theory // J. Mar. Syst. 2007. V. 64. P. 89.
Дополнительные материалы отсутствуют.
Инструменты
Физиология растений