

Генетика, 2021, T. 57, № 11, стр. 1261-1275
Особенности геномного распределения регионов высокой гомозиготности у коренного населения Северной Евразии на индивидуальном и популяционном уровнях на основе анализа SNP высокой плотности
Н. А. Колесников 1, *, **, В. Н. Харьков 1, А. А. Зарубин 1, М. О. Раджабов 2, М. И. Воевода 3, М. А. Губина 4, Э. К. Хуснутдинова 5, С. С. Литвинов 5, Н. В. Екомасова 5, О. В. Штыгашева 6, Н. Р. Максимова 7, А. Л. Сухомясова 7, В. А. Степанов 1
1 Научно-исследовательский институт медицинской генетики, Томский национальный исследовательский медицинский центр Российской академии наук
634050 Томск, Россия
2 Институт физики им. Х.И. Амирханова, Дагестанский федеральный исследовательский центр
Российской академии наук
367012 Махачкала, Россия
3 Федеральный исследовательский центр фундаментальной и трансляционной медицины
630060 Новосибирск, Россия
4 Федеральный исследовательский центр Институт цитологии и генетики
Сибирского отделения Российской академии наук
630090 Новосибирск, Россия
5 Институт биохимии и генетики, Уфимский федеральный исследовательский центр
Российской академии наук
450054 Уфа, Россия
6 Хакасский государственный университет им. Н.Ф. Катанова
655000 Абакан, Россия
7 Северо-Восточный федеральный университет им. М.К. Аммосова
677000 Якутск, Россия
* E-mail: nikita.kolesnikov@medgenetics.ru
** E-mail: nik.fleming@mail.ru
Поступила в редакцию 29.01.2021
После доработки 11.05.2021
Принята к публикации 20.05.2021
Аннотация
Анализ геномных данных по 886 889 аутосомных SNP позволил оценить уровень инбридинга на основе исследования регионов высокой гомозиготности (FROH) в выборке из 1836 человек, включающей 76 популяций коренного населения Восточной Европы, Кавказа, Волго-Уральского региона, Сибири и Средней Азии. Данные были получены с помощью биочипов Infinium Multi-Ethnic Global-8 Kit. Среди популяций Дагестана, внутри нахско-дагестанской языковой семьи, можно выделить представителей языковых групп с максимальным уровнем FROH: дидойская (0.0727) и андийская (0.0378) группы. Последний показатель близок к таковому в популяциях, говорящих на чукотско-камчатском и нивхском языках Сибири (0.0360). Для популяций Сибири характерно наибольшее значение суммарной длины коротких и средних регионов высокой гомозиготности на человека, по сравнению с другими исследованными популяциями. Популяции коренных народов Дагестана и Сибири в целом характеризуются большей степенью гомозиготизации генома и большей вариабельностью регионов с повышенной гомозиготностью по сравнению с другими популяциями Кавказа, Средней Азии, Европы и Волго-Уральского региона.
Существенная часть любого индивидуального генома человека представлена регионами высокой гомозиготности (англ. ROH – runs of homozygosity). Общая длина и распределение таких участков связаны с генетико-демографической историей популяции и отражают эффекты основателя, уровень инбридинга, а также могут быть следствием действия сильного направленного отбора [1, 2]. Современные методы анализа данных полных геномов или высокоплотных наборов SNP позволяют детально выявлять структуру и распределение ROH в индивидуальных геномах и популяциях с целью уточнения генетической истории популяций, оценки уровня инбридинга, поиска сигналов естественного отбора [1, 3]. Данные по распределению ROH в популяциях могут служить основой для идентификации участков генома, связанных с заболеваниями, в контексте картирования гомозиготности [4]. Общая длина и структура ROH характеризуются существенной межиндивидуальной и межпопуляционной вариабельностью. Как правило, сумма длин коротких и средних ROH у индивида возрастает от восточно-африканских к европейским и азиатским популяциям в силу накопления рекомбинационных событий. Напротив, общий суммарный размер длинных ROH показывает большие межиндивидуальные вариации, отражая, вероятно, эффекты недавнего инбридинга [5, 6]. Кроме того, показано, что частота распределения ROH в геноме коррелирует со скоростью рекомбинации, а также с сигналами недавнего положительного отбора [7].
Предыдущие исследования показали, что у неродственных людей широко распространены короткие ROH, измеряемые несколькими десятками тысяч пар нуклеотидов (тпн) [8]. Регионы высокой гомозиготности средних размеров – от сотен тысяч до нескольких миллионов пн – являются, как правило, результатом недавнего общего происхождения индивидов внутри популяции. Длинные ROH, измеряющиеся десятками миллионов пн, характерны для популяций с высоким уровнем близкородственных браков [5].
Среди многочисленных групп населения Северной Евразии, имеющих различную генетико-демографическую историю, особый интерес для исследования ROH представляют популяции коренных народов Северного Кавказа и Сибири, характеризующиеся, как правило, небольшой численностью и значительной географической изоляцией от других популяций. В частности, ранее для популяций Дагестана, говорящих на языках нахско-дагестанской семьи, были показаны высокие коэффициенты инбридинга, очень большое количество и большая суммарная длина гомозиготных блоков, по сравнению с окружающими популяционными группами Кавказа, Ближнего Востока, Европы, Центральной и Южной Азии [9]. Для популяций Сибири мы ранее на низкоплотных чипах обнаружили значительно большее число и суммарную длину ROH по сравнению с популяциями Волго-Уральского региона, Кавказа, Европы и Средней Азии [10].
Настоящая работа посвящена оценке уровня инбридинга, структуры и вариабельности ROH в популяциях Северной Евразии на основе анализа SNP высокой плотности.
МАТЕРИАЛЫ И МЕТОДЫ
Данные были получены с помощью биочипов “Infinium Multi-Ethnic Global-8 Kit”, содержащих 1 779 819 SNP, в выборке из 1836 человек. Популяции коренного населения Сибири (N = 477) представлены алтайцами (Б – с. Бешпельтир Чемальского муниципального района, N = 24 и К – с. Кулада Онгудайского района, N = 25), бурятами (А – п. Агинское Агинского района, N = 23 и К – с. Курумкан Курумканского района, N = 28), калмыками (N = 29), кетами (N = 15), коряками (N = 20), нивхами (N = 13), татарами (Т – г. Томск, N = 20), тувинцами (N = 28), удэгейцами (N = 15), хантами (К – с. Казым Белоярского района, N = 30 и Р – д. Русскинская Сургутского района, N = 26), хакасами (Т – сагайцами Таштыпского р-на, N = 29 и Ш – качинцами Ширинского р-на, N = 26), чукчами (N = 25), чулымцами (N = 22), эвенками (З – забайкальские (п. Чара Каларского района, с. Моклан и с. Тупик Тунгиро-Олёкминского района), N = 25 и Я – якутские, N = 28) и якутами (N = 26).
Коренные популяции Дагестана (N = 518) включают аварцев (N = 24), даргинцев (N = 28), лакцев (N = 24), табасаран (N = 21), лезгин (N = 28), агулов (N = 24), рутульцев (N = 22), цахуров (N = = 24), арчинцев (N = 24), андийцев (N = 17), ахвахцев (N = 24), тиндинцев (N = 18), каратинцев (N = = 24), багулалов (N = 23), ботлихцев (N = 16), хваршин (N = 15), цезов (N = 24), бежтинцев (N = 22), чамалинцев (N = 24), гунзибцев (N = 17), гинухцев (N = 19), кумыков (N = 37) и караногайцев (N = 19).
Волго-Уральский географический регион и популяции Европы (N = 419) представлены коми (N = 30), марийцами (N = 30), вепсами (N = 30), удмуртами (удмурты N = 30, Б – п. Балезино, N = = 28 и Ш– с. Шаркан, N = 18),чувашами (N = 26), башкирами (Б – Бурзянский р-н, N = 34, П – пермские, N = 15 и С – Салаватский р-н, N = 15), бесермянами (N = 16), карелами (N = 29), мордвой (Э – эрзя, N = 16, М – мокша, N = 30, Ш – шокша, N = 14), татарами (К– казанские, N = 33) и русскими (N = 33).
Коренные популяции Кавказского региона (N = 294) включают абхазов (N = 30), адыгейцев (N = 10), балкарцев (N = 50), ингушей (N = 30), карачаевцев (N = 22), мегрелов (N = 28), ногайцев (N = 20), осетин (С – северные, N = 30, Ю – южные, N = 17), черкесов (N = 30) и чеченцев (N = 27). Популяции северных киргизов (N = 28), узбеков (N = 22), казахов (М – младший жуз, N = 29, С – старший жуз, N = 26) и дунган (N = 23) относятся к популяциям Средней Азии (N = 128).
При биоинформатической обработке полученных данных из анализа были исключены SNP с более чем 10% отсутствующих генотипов. Были исключены все позиции делеций и инсерций. Данные были предварительно отфильтрованы по минимальной частоте редкого аллеля (MAF, minor allele frequency > 0.01). В итоге после фильтрации исходных данных по 1 779 819 маркерам в финальный массив данных вошло 886 889 аутосомных SNP. Кроме этого, были исключены образцы, имеющие более 5% пропущенных SNP, а также метисы первого, второго и третьего поколений. Для нахождения участков, которые с большей вероятностью представляют аутозиготность, перед поиском ROH в программе PLINK v1.9 [11] мы также удалили SNP с высоким неравновесием по сцеплению (LD) (англ. LD – linkage disequilibrium). Для этого использовали сканирование генома в рамке из 50 SNP со скользящим окном 5 SNP и порогом попарного коэффициента сцепления (r2 порог = 0.8). В результате контроль прошли 585 715 SNP. Для анализа ROH в PLINK указали следующие параметры: –homozyg-snp 50, –homozyg-kb 300. Все остальные параметры использовались по умолчанию. Анализ был выполнен с учетом трех различных наборов ROH с учетом минимальной длины 0.5, 1.5 и 5 млн пн (малые, средние и большие ROH).
Для расчета коэффициента инбридинга FROH у каждого индивидуума размеры аутосомных областей ROH, превышающих 1.5 млн пн, суммировали и затем делили на общую длину аутосом, за исключением центромер (2708.613716 млн пн для сборки генома hg19). Как показано ранее, FROH для 1.5 млн пн лучше коррелирует с оценками инбридинга, полученными из анализа родословных [1].
Для оценки возраста ROH применяли программу GARLIC [2], использующую метод, основанный на вероятностной модели для ROH в классах с “коротким” (A, до 0.5 млн пн), “средним” (B, 0.5–1.5 млн пн) и “длинным” (C, более 1.5 млн пн) размерами. Короткие ROH отражают гомозиготность по древним гаплотипам, средние ROH являются результатом недавних демографических изменений в популяции (например, эффект “бутылочного горлышка”) и длинные ROH отражают гомозиготность возникшую в результате недавних кровнородственных браков. Для анализа ROH в GARLIC указали следующие параметры: ‒build hg19, –error 0.001, –winsize 40, –auto-winsize, –auto-winsize-step 2, –kde-subsample 0, –ld-subsample 0, –resample 40.
Для межпопуляционных сравнений были рассчитаны средние значения количества и совокупной длины ROH на геном для каждой популяции. Статистический анализ проводился в программной среде R. Статистическую значимость межпопуляционных различий оценивали при помощи теста Вилкоксона. Для коррекции уровня значимости по причине множественных сравнений использовалась поправка Бонферрони (уровень значимости = 0.05).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Коэффициент инбридинга FROH
Коэффициенты инбридинга FROH для всех индивидов из 76 популяционных выборок оценивали отдельно и группировали по географическим регионам и языковым группам для трех классов ROH (длина больше трех пороговых значений 0.5, 1.5 и 5 млн пн). Коэффициенты FROH для трех диапазонов длин ROH, среднее суммарное количество и средняя суммарная длина гомозиготных участков в исследованных популяциях представлены в табл. 1. Примечательно, что ROH > 1.5 млн пн не было выявлено лишь у четырех индивидов (кумыка, лезгина, эвенка и узбека).
Таблица 1.
Значения коэффициентов FROH для трех диапазонов длин ROH, среднее суммарное количество (А, В, С) и средняя суммарная длина (а, b, c) для трех классов длин исследуемых популяций
| Популяция | Регион | Группа | PLINK | GARLIC | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FROH | FROH | NROH | SROH | Σ | ||||||||||||
| 0.5 | 1.5 | 5 | 0.5 | 1.5 | 5 | A | B | C | a | b | c | NROH | SROH | |||
| Башкиры (Б) | В | 1 | 0.0438 | 0.0122 | 0.0043 | 0.0388 | 0.0094 | 0.0035 | 544 | 189 | 14 | 95 | 95 | 31 | 746 | 221 |
| Башкиры (П) | В | 1 | 0.0450 | 0.0147 | 0.0065 | 561 | 188 | 17 | 95 | 95 | 45 | 765 | 235 | |||
| Башкиры (С) | В | 1 | 0.0368 | 0.0083 | 0.0034 | 439 | 157 | 10 | 69 | 74 | 24 | 606 | 167 | |||
| Татары (К) | В | 1 | 0.0334 | 0.0061 | 0.0023 | 575 | 192 | 9 | 100 | 97 | 20 | 776 | 216 | |||
| Чуваши | В | 1 | 0.0406 | 0.0103 | 0.0032 | 443 | 136 | 15 | 79 | 70 | 31 | 594 | 180 | |||
| Бесермяне | В | 2 | 0.0498 | 0.0169 | 0.0063 | 0.0509 | 0.0199 | 0.0105 | 678 | 202 | 16 | 118 | 110 | 55 | 896 | 283 |
| Коми | В | 2 | 0.0468 | 0.0164 | 0.0084 | 453 | 111 | 13 | 84 | 63 | 43 | 577 | 191 | |||
| Удмурты | В | 2 | 0.0504 | 0.0158 | 0.0047 | 476 | 143 | 19 | 89 | 81 | 46 | 637 | 216 | |||
| Удмурты (B) | В | 2 | 0.0494 | 0.0155 | 0.0037 | 576 | 197 | 24 | 107 | 107 | 52 | 796 | 266 | |||
| Удмурты (Ш) | В | 2 | 0.0514 | 0.0171 | 0.0039 | 574 | 205 | 28 | 98 | 105 | 57 | 806 | 259 | |||
| Вепсы | В | 3 | 0.0445 | 0.0107 | 0.0027 | 0.0458 | 0.0126 | 0.0031 | 432 | 145 | 18 | 76 | 75 | 38 | 594 | 188 |
| Карелы | В | 3 | 0.0432 | 0.0089 | 0.0024 | 453 | 153 | 16 | 80 | 78 | 31 | 622 | 189 | |||
| Марийцы | В | 4 | 0.0437 | 0.0117 | 0.0040 | 0.0496 | 0.0162 | 0.0044 | 450 | 159 | 23 | 81 | 83 | 51 | 632 | 215 |
| Мордва (М) | В | 4 | 0.0396 | 0.0082 | 0.0026 | 595 | 179 | 10 | 109 | 97 | 24 | 784 | 231 | |||
| Мордва (Ш) | В | 4 | 0.0460 | 0.0141 | 0.0067 | 635 | 189 | 13 | 109 | 98 | 39 | 837 | 246 | |||
| Мордва (Э) | В | 4 | 0.0396 | 0.0083 | 0.0026 | 596 | 204 | 16 | 100 | 99 | 30 | 816 | 229 | |||
| Русские | В | 5 | 0.0377 | 0.0061 | 0.0019 | 0.0377 | 0.0061 | 0.0019 | 607 | 189 | 8 | 111 | 102 | 19 | 804 | 231 |
| Агулы | Д | 6 | 0.0490 | 0.0193 | 0.0119 | 0.0464 | 0.0171 | 0.0104 | 617 | 163 | 11 | 114 | 93 | 46 | 790 | 253 |
| Арчинцы | Д | 6 | 0.0782 | 0.0459 | 0.0315 | 547 | 161 | 25 | 106 | 94 | 123 | 733 | 323 | |||
| Лезгины | Д | 6 | 0.0412 | 0.0124 | 0.0076 | 568 | 156 | 8 | 105 | 88 | 33 | 733 | 226 | |||
| Рутульцы | Д | 6 | 0.0433 | 0.0131 | 0.0072 | 445 | 131 | 10 | 78 | 70 | 34 | 586 | 182 | |||
| Табасараны | Д | 6 | 0.0350 | 0.0066 | 0.0028 | 573 | 196 | 9 | 101 | 99 | 20 | 778 | 220 | |||
| Цахуры | Д | 6 | 0.0499 | 0.0206 | 0.0120 | 463 | 132 | 14 | 81 | 70 | 54 | 609 | 206 | |||
| Андийцы | Д | 7 | 0.0696 | 0.0378 | 0.0240 | 0.0578 | 0.0279 | 0.0185 | 577 | 167 | 17 | 101 | 90 | 76 | 761 | 266 |
| Ахвахцы | Д | 7 | 0.1020 | 0.0678 | 0.0430 | 424 | 114 | 35 | 90 | 79 | 174 | 572 | 343 | |||
| Багулалы | Д | 7 | 0.0721 | 0.0400 | 0.0253 | 621 | 171 | 22 | 119 | 103 | 105 | 814 | 328 | |||
| Ботлихцы | Д | 7 | 0.0512 | 0.0196 | 0.0102 | 673 | 230 | 20 | 114 | 117 | 59 | 924 | 290 | |||
| Каратинцы | Д | 7 | 0.0775 | 0.0460 | 0.0321 | 547 | 164 | 24 | 101 | 93 | 122 | 735 | 316 | |||
| Тиндинцы | Д | 7 | 0.0569 | 0.0253 | 0.0139 | 536 | 164 | 16 | 95 | 91 | 66 | 715 | 253 | |||
| Чамалалы | Д | 7 | 0.0697 | 0.0377 | 0.0246 | 501 | 151 | 23 | 91 | 82 | 102 | 675 | 276 | |||
| Бежтинцы | Д | 8 | 0.1126 | 0.0727 | 0.0460 | 0.1105 | 0.0689 | 0.0417 | 423 | 113 | 28 | 101 | 97 | 158 | 563 | 356 |
| Гинухцы | Д | 8 | 0.1437 | 0.1039 | 0.0733 | 259 | 71 | 27 | 80 | 85 | 226 | 357 | 391 | |||
| Гунзибцы | Д | 8 | 0.1305 | 0.0887 | 0.0549 | 318 | 98 | 33 | 85 | 95 | 204 | 449 | 383 | |||
| Хваршины | Д | 8 | 0.0773 | 0.0448 | 0.0309 | 594 | 183 | 24 | 108 | 102 | 121 | 801 | 331 | |||
| Цезы | Д | 8 | 0.1011 | 0.0571 | 0.0293 | 492 | 141 | 31 | 109 | 108 | 137 | 664 | 354 | |||
| Караногайцы | Д | 9 | 0.0318 | 0.0037 | 0.0010 | 0.0324 | 0.0039 | 0.0011 | 582 | 198 | 13 | 99 | 94 | 20 | 793 | 213 |
| Кумыки | Д | 9 | 0.0312 | 0.0036 | 0.0009 | 548 | 183 | 9 | 100 | 95 | 16 | 739 | 210 | |||
| Даргинцы | Д | 10 | 0.0696 | 0.0369 | 0.0243 | 0.0696 | 0.0369 | 0.0243 | 594 | 143 | 19 | 114 | 87 | 97 | 756 | 297 |
| Лакцы | Д | 11 | 0.0666 | 0.0347 | 0.0230 | 0.0666 | 0.0347 | 0.0230 | 587 | 179 | 20 | 107 | 100 | 93 | 786 | 300 |
| Аварцы | Д | 12 | 0.0761 | 0.0442 | 0.0320 | 0.0761 | 0.0442 | 0.0320 | 589 | 178 | 24 | 109 | 100 | 119 | 791 | 328 |
| Абхазы | К | 13 | 0.0335 | 0.0044 | 0.0006 | 0.0350 | 0.0047 | 0.0004 | 519 | 224 | 22 | 86 | 101 | 28 | 764 | 215 |
| Адыгейцы | К | 13 | 0.0339 | 0.0048 | 0.0004 | 552 | 193 | 14 | 91 | 91 | 21 | 759 | 204 | |||
| Черкесы | К | 13 | 0.0317 | 0.0037 | 0.0010 | 589 | 173 | 7 | 106 | 92 | 13 | 769 | 211 | |||
| Балкарцы | К | 14 | 0.0345 | 0.0069 | 0.0028 | 0.0357 | 0.0071 | 0.0027 | 514 | 151 | 8 | 100 | 85 | 21 | 674 | 205 |
| Карачаевцы | К | 14 | 0.0382 | 0.0104 | 0.0055 | 589 | 172 | 10 | 106 | 92 | 30 | 771 | 228 | |||
| Ногайцы | К | 14 | 0.0295 | 0.0030 | 0.0003 | 528 | 239 | 21 | 83 | 102 | 24 | 788 | 209 | |||
| Ингуши | К | 15 | 0.0396 | 0.0099 | 0.0030 | 0.0392 | 0.0094 | 0.0026 | 521 | 196 | 16 | 91 | 98 | 33 | 733 | 222 |
| Чеченцы | К | 15 | 0.0399 | 0.0105 | 0.0034 | 550 | 177 | 15 | 99 | 92 | 33 | 741 | 223 | |||
| Осетины (С) | К | 16 | 0.0363 | 0.0066 | 0.0017 | 0.0359 | 0.0063 | 0.0015 | 515 | 220 | 20 | 86 | 101 | 31 | 755 | 217 |
| Осетины (Ю) | К | 16 | 0.0367 | 0.0069 | 0.0018 | 570 | 212 | 20 | 92 | 97 | 31 | 801 | 220 | |||
| Мегрелы | К | 17 | 0.0369 | 0.0049 | 0.0006 | 0.0369 | 0.0049 | 0.0006 | 529 | 202 | 16 | 93 | 100 | 23 | 747 | 216 |
| Алтайцы (Б) | С | 18 | 0.0533 | 0.0171 | 0.0089 | 0.0519 | 0.0173 | 0.0086 | 517 | 163 | 14 | 93 | 86 | 46 | 695 | 224 |
| Алтайцы (К) | С | 18 | 0.0552 | 0.0168 | 0.0100 | 632 | 177 | 13 | 121 | 101 | 47 | 821 | 269 | |||
| Татары (Т) | С | 18 | 0.0338 | 0.0071 | 0.0037 | 483 | 127 | 5 | 86 | 68 | 17 | 615 | 171 | |||
| Тувинцы | С | 18 | 0.0591 | 0.0151 | 0.0061 | 731 | 242 | 19 | 139 | 136 | 46 | 992 | 322 | |||
| Хакасы (Т) | С | 18 | 0.0664 | 0.0318 | 0.0183 | 585 | 173 | 23 | 111 | 97 | 87 | 781 | 295 | |||
| Хакасы (Ш) | С | 18 | 0.0534 | 0.0146 | 0.0070 | 652 | 199 | 12 | 122 | 112 | 38 | 862 | 273 | |||
| Буряты (А) | С | 19а | 0.0656 | 0.0181 | 0.0076 | 0.0599 | 0.0112 | 0.0027 | 691 | 317 | 31 | 116 | 152 | 50 | 1040 | 318 |
| Буряты (К) | С | 19а | 0.0615 | 0.0166 | 0.0081 | 733 | 235 | 16 | 135 | 130 | 47 | 983 | 312 | |||
| Калмыки | С | 19а | 0.0465 | 0.0058 | 0.0008 | 688 | 260 | 23 | 121 | 126 | 33 | 970 | 280 | |||
| Эвенки (З) | С | 19б | 0.0632 | 0.0195 | 0.0080 | 608 | 195 | 19 | 111 | 110 | 55 | 822 | 276 | |||
| Эвенки (Я) | С | 19б | 0.0786 | 0.0256 | 0.0105 | 705 | 221 | 23 | 138 | 133 | 69 | 949 | 339 | |||
| Удэгейцы | С | 19б | 0.0774 | 0.0246 | 0.0134 | 764 | 219 | 14 | 152 | 136 | 59 | 997 | 347 | |||
| Якуты | С | 19в | 0.0723 | 0.0233 | 0.0097 | 725 | 240 | 25 | 137 | 137 | 68 | 990 | 343 | |||
| Кеты | С | 20а | 0.0707 | 0.0274 | 0.0127 | 0.0720 | 0.0259 | 0.0121 | 753 | 221 | 21 | 139 | 132 | 70 | 995 | 341 |
| Ханты (К) | С | 20б | 0.0716 | 0.0280 | 0.0119 | 620 | 201 | 24 | 123 | 125 | 75 | 844 | 323 | |||
| Ханты (Р) | С | 20б | 0.0716 | 0.0266 | 0.0102 | 633 | 212 | 24 | 120 | 126 | 70 | 869 | 316 | |||
| Чулымцы | С | 20в | 0.0676 | 0.0292 | 0.0167 | 541 | 165 | 22 | 99 | 92 | 79 | 727 | 270 | |||
| Нивхи | С | 21а | 0.1139 | 0.0398 | 0.0133 | 0.1073 | 0.0363 | 0.0119 | 699 | 239 | 28 | 147 | 161 | 84 | 965 | 393 |
| Коряки | С | 21б | 0.1209 | 0.0422 | 0.0142 | 806 | 301 | 30 | 172 | 210 | 98 | 1136 | 480 | |||
| Чукчи | С | 21б | 0.1136 | 0.0409 | 0.0137 | 703 | 257 | 32 | 149 | 174 | 100 | 992 | 424 | |||
| Казахи (М) | С. А. | 22а | 0.0367 | 0.0055 | 0.0018 | 0.0360 | 0.0040 | 0.0006 | 575 | 198 | 13 | 100 | 98 | 19 | 786 | 217 |
| Казахи (С) | С. А. | 22а | 0.0358 | 0.0039 | 0.0002 | 511 | 232 | 25 | 83 | 101 | 28 | 768 | 212 | |||
| Киргизы | С. А. | 22а | 0.0414 | 0.0067 | 0.0022 | 589 | 194 | 8 | 106 | 103 | 19 | 791 | 227 | |||
| Узбеки | С. А. | 22а | 0.0334 | 0.0073 | 0.0043 | 549 | 170 | 6 | 97 | 88 | 20 | 725 | 205 | |||
| Дунгане | С. А. | 22б | 0.0562 | 0.0116 | 0.0070 | 0.0562 | 0.0116 | 0.0070 | 808 | 218 | 6 | 151 | 126 | 27 | 1032 | 304 |
| В | 0.0438 | 0.0122 | 0.0043 | 535 | 173 | 16 | 94 | 90 | 37 | 723 | 221 | |||||
| Д | 0.0704 | 0.0377 | 0.0239 | 525 | 156 | 20 | 100 | 93 | 96 | 701 | 289 | |||||
| К | 0.0357 | 0.0065 | 0.0018 | 543 | 196 | 15 | 94 | 96 | 26 | 755 | 215 | |||||
| С | 0.0702 | 0.0229 | 0.0099 | 663 | 218 | 21 | 127 | 127 | 62 | 902 | 316 | |||||
| С. А. | 0.0406 | 0.0067 | 0.0028 | 606 | 202 | 12 | 107 | 103 | 23 | 820 | 233 | |||||
Примечание 1. В – Волго-Уральский регион и Европа, К – Кавказ, Д – Дагестан, С – Сибирь, С. А. – Средняя Азия. Примечание 2. 1 – алтайская/тюркская; 2 – пермская подгруппа; 3 – прибалтийско-финская подгруппа; 4 – волжско-финская подгруппа; 5 – восточнославянская группа; 6 – нахско-дагестанская/лезгинская ветвь; 7 – нахско-дагестанская/андийская группа; 8 – нахско-дагестанская/дидойская группа; 9 – алтайская/тюркская; 10 – нахско-дагестанская/даргинская ветвь; 11 – нахско-дагестанская/лакская ветвь; 12 – нахско-дагестанская/аваро-андийская группа; 13 – абхазо-адыгские языки; 14 – алтайская/тюркская; 15 – нахско-дагестанская/нахская ветвь; 16 – индоевропейская/иранская группа; 17 – картвельская семья; 18 – алтайская/тюркская; 19 – а) алтайская/монгольская, б) алтайская/тунгусо-маньчжурские языки, в) алтайская/тюркская; 20 – а) енисейская, б) уральская/обско-угорская, в) алтайская/тюркская; 21 – а) палеоазиатский язык, б) чукотско-камчатские языки; 22 – а) алтайская/тюркская, б) сино-тибетские языки/китайские языки.
Максимальные значения по FROH > 1.5 (0.0376) и FROH > 5 (0.0239) наблюдаются в популяциях Дагестана (табл. 1). Наибольшим коэффициентом геномного инбридинга для длин ROH > 1.5 млн пн обладают популяции гинухцев и гунзибцев (FROH = = 0.1039, FROH = 0.0887). Этот показатель для гинухцев является одним из самых высоких в мире и схож лишь с уровнем инбридинга в изолированной популяции индейцев каритиана из бразильской Амазонии с общей численностью в 320 человек (FROH > 1.5 = 0.1067) [3]. Также по уровню гомозиготизации генома значительно выделяется и ряд других малочисленных народов Дагестана – бежтинцы, ахвахцы и цезы (FROH 0.0689, 0.0678 и 0.0571 соответственно). Эти результаты хорошо соотносятся с предыдущими данными по популяциям Дагестана [9], где для народов нахско-дагестанской языковой семьи были выявлены более высокие коэффициенты инбридинга по сравнению с популяциями из других географических регионов и не нахско-дагестанскими популяциями Дагестана.
Внутри нахско-дагестанской языковой семьи по FROH этносы разделяются на три группы, в полном соответствии с их языковой классификацией. Максимальный средний уровень FROH выявлен для популяций дидойской (цезской) группы (0.0727), проживающих в высокогорных регионах в течение сотен поколений в относительной изоляции и имеющих также значительно более низкий уровень разнообразия по гаплогруппам Y-хромосомы, по сравнению с другими северокавказскими популяциями [12]. Для андийской группы показано гораздо меньшее значение (0.0378), находящееся на одном уровне с выборками чукчей, коряков и нивхов (0.0360). Наименьший коэффициент инбридинга выявлен для лезгинской группы (0.0193), занимающей в основном низменные территории и предгорья южной части Дагестана и отличающейся гораздо большей по сравнению с дидойской и андийской группами численностью представляющих ее этносов. Эти результаты полностью согласуются с данными об очень высокой частоте инбредных браков среди малочисленных и более изолированных популяций высокогорных районов по сравнению с равнинными [13].
Среди коренного населения Сибири максимальные значение FROH > 1.5 обнаружены у дальневосточных коряков, чукчей и нивхов (0.0422, 0.0409, 0.0363), что вполне закономерно в связи с их относительной малочисленностью и территориальной изолированностью. Высокое значение показано и для выборки хакасов-сагайцев из предгорного Таштыпского района (0.0318), вдвое превосходящее хакасов-качинцев равнинного Ширинского района. Минимальное значение характерно для томских татар (0.0071). Для остальных популяций наблюдается разброс FROH > 1.5 от 0.0112 до 0.0292. Несмотря на то что FROH в популяциях Дагестана значительно превышает по средним и длинным ROH (значения FROH > 5, FROH > 1.5), для популяций Сибири по более коротким участкам гомозиготности (FROH > 0.5) наблюдаются схожие значения. Это может свидетельствовать о значительных эффектах инбридинга для этих популяций Сибири в прошлом с постепенным уменьшением доли кровнородственных браков к настоящему времени.
Популяции Волго-Уральского региона занимают промежуточное положение среди других регионов по коэффициенту инбридинга для всех трех групп гомозиготных регионов. Это можно объяснить как его территориальным положением на месте пересечения путей различных миграционных потоков, так и давней традицией заключения браков лишь с родственниками не ближе пятого–шестого колена, что характерно для башкир, казанских татар, чувашей, коми, марийцев и мордвы. Внутри Волго-Уральского региона для популяций с низкой эффективной численностью и проживающих на периферии наблюдаются более высокие значения уровня гомозиготности генома. Максимальное значение обнаружено у бесермян (0.0199), обособленных территориально и имеющих численность около 2 тыс. человек, минимальное значение – у казанских татар и русских (0.0061).
Популяции Кавказа и Средней Азии обладают минимальными значениями коэффициентов инбридинга, оцененных по ROH, – от 0.0030 до 0.0116 для FROH > 1.5. Среди популяций Средней Азии максимальные значения наблюдаются у дунган (0.0116) что, вероятно, объясняется их недавней миграцией из Китая с сохранением их языкового, культурного и демографического единства на фоне практически полного отсутствия межнациональных браков. Ранее было показано, что дунгане Узбекистана генетически намного ближе к популяциям Китая и Северо-Восточной Индии, чем к соседним с ними узбекским популяциям, и не имеют с ними недавнего обмена генами [14].
Оценка ROH с помощью PLINK
В отличие от модельного подхода для поиска ROH, этот анализ не зависит от оценки частоты популяционных аллелей. Общая длина и количество ROH для каждого индивидуального генома колеблется от 41.2 до 687.7 млн пн и от 61 до 394 соответственно. Медианное число ROH на индивидуальный геном в исследованных образцах составило 135.4, граница первого квартиля 121.8, граница третьего квартиля 151.5. Медианная длина ROH 128.4 млн пн, граница первого квартиля 103.7 млн пн, третьего 139.3 млн пн. Среднее значение совокупной длины ROH на индивидуума варьирует от 79.9 млн пн в выборке ногайцев до 289.1 млн пн у гинухцев. Минимальное среднее популяционное значение числа ROH на человека наблюдается у узбеков (103.7), максимальное – среди коряков (312).
Доля ROH различной длины значительно колеблется в зависимости от географического региона и языковой группы. Выборки из Дагестана и Сибири, особенно носители языков нахско-дагестанской группы, чукотско-камчатских и нивхского языков, разительно отличаются от таковых из других географических регионов. Среднее значение общей длины ROH на человека в нашей выборке, говорящей на языках дидойской подгруппы, 305.03 млн пн сравнимо со значением 308.6 млн пн у представителей чукотско-камчатских и нивхского языков. Это полностью согласуется с оценкой геномного инбридинга в этих популяциях.
По доле образцов с одним и более ROH длиной больше 5 млн пн по регионам выделяются три различные группы, которые явно связаны с эндогамией и изоляцией: большая часть дагестанских (78.1%) и сибирских (75.5%) образцов имеет длинные ROH, далее идут кавказский, Волго-Уральский регион и Европа (57.9 и 38% соответственно). Для Среднеазиатского региона наблюдается минимальная доля геномов, имеющих длинные ROH (15.6%). Это полностью соответствует полученным ранее данным о наибольшем генетическом разнообразии и многокомпонентности генофонда среднеазиатских популяций в сравнении с другими этносами Северной Евразии. При этом наблюдаются значительные различия между популяциями внутри каждого региона. Например, у казахов старшего жуза всего 7% образцов имеют ROH больше 5 млн пн. Для двух человек из эвенкийской (З) и казахской (С) популяций не было найдено ROH длиннее 1.5 млн пн.
Оценка ROH с помощью GARLIC
Общая длина и количество ROH, идентифицируемые с помощью пакета GARLIC, для каждого индивида колеблются от 124 до 788.6 млн пн и от 319 до 1237 соответственно. Медианное число ROH на индивидуальный геном в исследованных образцах составило 774.5, граница первого квартиля 754.1, граница третьего квартиля 798.5. Медианная длина ROH – 261.4 млн пн, граница первого квартиля 243.9 млн пн, третьего 282.8 млн пн. Среднее значение совокупной длины ROH на индивидуума варьирует от 167.3 млн пн в выборке башкир (С) до 480.2 млн пн у коряков. Минимальное среднее популяционное значение числа ROH на человека наблюдается у гинухцев (356.8) из-за большей длины ROH за счет сильного инбридинга, максимальное – среди коряков (1136).
Средние размеры ROH классов А и B одинаковы среди популяций одного географического региона, за исключением того, что популяции Дагестана и Сибири имеют большую изменчивость (табл. 2). Максимальные значения наблюдаются у дагестанских популяций (гинухцев, гунзибцев, ахвахцев, цезов).
Таблица 2.
Суммарная статистика для значений границ классов длин (тпн) ROH по регионам
| Статистика | Сибирь | Средняя Азия | Дагестан | Кавказ | Волго-Уральский регион и Европа | Все |
|---|---|---|---|---|---|---|
| Классы A и B | ||||||
| Мин. | 305.54 | 287.59 | 315.23 | 279.41 | 301.07 | 279.41 |
| Макс. | 417.37 | 369.87 | 668.22 | 370.51 | 370.79 | 668.22 |
| Сред. знач. | 366.96 | 333.15 | 390.07 | 321.02 | 338.75 | 358.55 |
| SD | 7.28 | 15.00 | 17.94 | 8.89 | 4.35 | 6.67 |
| Классы B и C | ||||||
| Мин. | 942.65 | 777.099 | 981.114 | 787.342 | 1010.64 | 777.10 |
| Макс. | 1553.83 | 1402.82 | 2598.01 | 1200.04 | 1257.98 | 2598.01 |
| Сред. знач. | 1260.15 | 1115.65 | 1380.02 | 999.93 | 1112.28 | 1214.99 |
| SD | 38.33 | 120.00 | 77.19 | 44.52 | 20.96 | 31.39 |
Наибольшие средние значения класса С также наблюдаются у популяций Дагестана и Сибири, далее у популяций Средней Азии, Волго-Уральского региона и Европы, минимальные средние значения – у кавказских популяций. У народов, имеющих смешанное происхождение, – казахов (М, С), бурятов (А), калмыков, караногайцев и кумыков средние размеры ROH аналогичны кавказским популяциям.
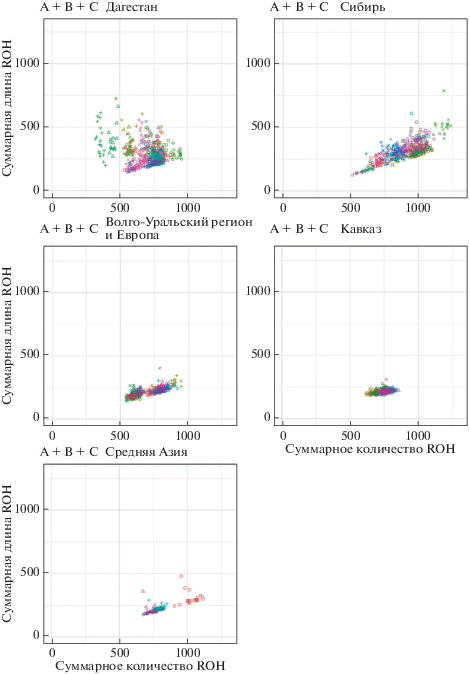
Для сибирских чукчей, коряков, нивхов для всех классов длин наблюдается значительное увеличение суммарной длины и количества ROH на человека по сравнению с другими популяциями, что свидетельствует о продолжительном инбридинге, а также эффекте “бутылочного горлышка” (рис. 1). Для трех этих этносов также наблюдается значительное увеличение суммарной длины среднего класса ROH на человека как по сравнению с остальными популяциями, так и по сравнению с классом коротких ROH внутри популяции, что подтверждает продолжительность инбридинга и небольшую эффективную численность популяций. Далее по коротким и средним длинам ROH выделяются кеты, якуты, эвенки (Я), удэгейцы, тувинцы и буряты (K). Для якутов и тувинцев, несмотря на большой эффективный размер популяции, большое количество и длина коротких и средних ROH вероятно связаны с сильным эффектом основателя, который выявляется на маркерах мтДНК и Y-хромосомы [15, 16]. В популяции эвенков (З) обнаружены образцы с признаками метисации, что может значительно занижать значения по длинам и количеству ROH. При анализе Y хромосомных линий у них наблюдаются европейские гаплогруппы, не характерные для коренных сибирских популяций. При анализе аутосомных блоков, идентичных по происхождению (IBD – англ. identical by descent), у эвенков также наблюдается значительное количество длинных блоков (4 млн пн), общих с европейскими популяциями, что свидетельствует об относительно недавней метисации этой группы людей с пришлыми европейцами. Для бурятов (А) также характерно увеличение суммарной длины среднего класса ROH, но в большей степени не из-за длины фрагментов, а из-за их количества, что свидетельствует об эффекте “бутылочного горлышка” в прошлом (рис. 1). У бурятов (А) и калмыков наблюдается увеличение количества ROH, но без увеличения суммарной длины, наиболее ярко выражено для ROH средней длины, что свидетельствует о том, что они происходили из довольно небольших полуизолированных сообществ, где эндогамия не была редкостью. По средней суммарной длине и количеству для средних и коротких ROH максимальные значения наблюдаются у сибирских популяций (табл. 1).
Рис. 1.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях Сибири.

Для популяций гинухцев, гунзибцев, бежтинцев, ахвахцев и цезов для всех классов ROH наблюдается распределение соотношения суммарной длины и количества, характерное для популяций, подверженных сильному инбридингу (рис. 2). Распределение ROH связано с размером популяции и их эффективной численностью, при этом относительно небольшие популяции Дагестана и Сибири имеют ROH большего количества и размера, чем большие по численности популяции этих регионов. Инбридинг добавляет небольшое количество очень длинных ROH для тех, кто является потомком кровнородственных браков, увеличивая дисперсию суммы ROH, видимую как сдвиг влево на графике длин ROH по сравнению с количеством ROH.
Рис. 2.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях Дагестана.

Кавказский регион характеризуется меньшим разбросом по суммарной длине и количеству регионов высокой гомозиготности на индивида, чем в остальных географических регионах. Для длинных ROH наблюдается тенденция к увеличению количества, но не длины, что вероятно объясняется сильной метисацией населения. Среди кавказских популяций выделяется популяция балкарцев, являющаяся одним из самых высокогорных народов Европы и имеющая более длинные короткие ROH, чем у остальных популяций Кавказа, что может свидетельствовать о сильном инбридинге в прошлом (рис. 3).
Рис. 3.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях Кавказа.

В популяциях, смешанных по происхождению и характеризующихся высоким уровнем генетического разнообразия, объединяются различные гаплотипы, что обычно значительно снижает количество и длину ROH. Это обнаружено у томских и казанских татар, узбеков и башкир. Для Волго-Уральского региона и Европы наблюдается разделение на две группы по количеству ROH, где основной вклад вносят короткие и средние ROH. В первую группу входят марийцы, коми, вепсы, чуваши, карелы, удмурты и башкиры (С), во вторую – бесермяне, мордва (М, Ш, Э), удмурты (Ш, Б), татары (К), башкиры (Б, П) (рис. 4).
Рис. 4.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях Волго-Уральского региона и Европы.

Эффект “бутылочного горлышка” в демографической истории конкретного этноса увеличивают количество ROH, которые обычно все еще относительно короткие, например популяции калмыков и бурятов (А). У некоторых популяций, имеющих как эффект “бутылочного горлышка”, так и распространенность близкородственных браков, наблюдаются большее количество и длина ROH с увеличением дисперсии суммы ROH (коряки, чукчи и нивхи).
Наибольшими отличиями среди популяций Средней Азии характеризуется популяция дунган, в ней наблюдаются высокие значения по количеству и суммарной длине для средних и коротких ROH и минимальные значения для длинных (рис. 5).
Рис. 5.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях Средней Азии.

Доля генома в ROH разной длины и разного количества сильно варьирует по регионам (рис. 6). Наши результаты согласуются с рядом недавних исследований, проведенных с использованием высокоплотных чипов. Уровень гомозиготности по всему геному показывает самый высокий уровень инбридинга в дагестанских и сибирских группах, говорящих на дидойских, чукотко-камчатских и нивхском языках, по сравнению с остальными северо-евразийскими популяционными выборками. Дагестанские и сибирские популяции обладают самыми длинными геномными участками гомозиготности для всех категорий длины ROH и имеют более высокую долю людей с длинными областями аутозиготности.
Рис. 6.
Отношение суммарной длины ROH (млн пн) к суммарному количеству ROH в исследуемых популяциях для каждого географического региона.
Высокий коэффициент инбридинга и наличие длинных аутозиготных сегментов согласуются с предыдущими выводами для коэффициентов инбридинга Дагестана [17–19], рассчитанными на основе данных родословных и высокоплотных чипов, а также с данными по Y-хромосомным маркерам указывающими на эффекты основателя и высокую подразделенность по языковым группам [12]. Аналогично для работ по популяционной генетике сибирских этносов с использованием нейтральных маркерных систем Y-хромосомной, митохондриальной ДНК, показано, что во многих популяциях наблюдается эффект основателя по отдельным гаплогруппам с низким генетическим разнообразием, которое свидетельствует о событиях экспансии численности в относительно недавнее время [15, 20, 21]. Интересно, что полученные нами в настоящей работе оценки существенно не отличаются от тех, которые были получены в предыдущем исследовании нашей группы [10] с использованием гораздо меньшего числа маркеров на меньших выборках сибирских популяций.
Для популяций Сибири характерно большее значение суммарной длины классов А и В на человека, что соответствует данным об увеличении длин классов А и В с удалением популяции от Африки. Эта тенденция похожа на наблюдаемое сокращение разнообразия гаплотипов с увеличением расстояния от Африки. Суммарная длина класса С ROH более изменчива и значительно больше в большинстве популяций Дагестана и Сибири по сравнению с популяциями Кавказа, Средней Азии, Европы и Волго-Уральского региона. Это также свидетельствует о том, что большая доля популяций из Дагестана и Сибири имеют более высокий уровень кровного родства, это аналогично данным, полученным по значениям коэффициента инбридинга (FROH).
Результаты проведенного анализа показывают, что генофонд населения Северной Евразии демонстрирует значительные различия в распределении суммы длин и количества по всем классам ROH как между исследуемыми географическими регионами, этносами, так и внутри локальных популяций. Особенно это характерно для территориально удаленных популяций чукчей и коряков, а также культурно и географически изолированных горных популяций Дагестана. Коренные этносы высокогорных районов Дагестана показывают максимальные значения инбридинга, сохранившегося на высоком уровне до настоящего времени. Большее количество и длина коротких и средних ROH у сибирских популяций указывают на более древний и более продолжительный инбридинг. Получено подтверждение значительной генетической гетерогенности среднеазиатских популяций и относительно низком уровне инбридинга у большинства народов Кавказа и Волго-Уральского региона. Полученные результаты позволяют существенно уточнить имеющиеся данные об особенностях структуры генофонда всех исследованных этносов. Данные о аутозиготных блоках сцепления в конкретных популяциях также могут быть использованы при изучении их компонентного состава предрасположенности к различным наследственным заболеваниям.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 19-34-90101.
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
McQuillan R., Leutenegger A.L., Abdel-Rahman R. et al. Runs of homozygosity in European populations // Am. J. Hum. Genet. 2008. V. 83. № 3. P. 359–372. https://doi.org/10.1016/j.ajhg.2008.08.007
Szpiech Z.A., Blant A., Pemberton T.J. GARLIC: Genomic autozygosity regions likelihood-based inference and classification // Bioinformatics. 2017. V. 33. № 13. P. 2059–2062. https://doi.org/10.1093/bioinformatics/btx102
Lemes R.B., Nunes K., Carnavalli J.E.P. et al. Inbreeding estimates in human populations: Applying new approaches to an admixed Brazilian isolate // PLoS One. 2018. V. 13. № 4. P. e0196360. https://doi.org/10.1371/journal.pone.0196360
Ropers H. New perspectives for the elucidation of genetic disorders // Am. J. Hum. Genet. 2007. V. 81. № 2. P. 199–207. https://doi.org/10.1086/520679
Kirin M., McQuillan R., Franklin C.S. et al. Genomic runs of homozygosity record population history and consanguinity // PLoS One. 2010. V. 5. № 11. e13996. https://doi.org/10.1371/journal.pone.0013996
Pemberton T.J., Absher D., Feldman M.W. et al. Genomic patterns of homozygosity in worldwide human populations // Am. J. Hum. Genet. 2012. V. 91. № 2. P. 275–292. https://doi.org/10.1016/j.ajhg.2012.06.014
Ceballos F.C., Joshi P.K., Clark D.W. et al. Runs of homozygosity: Windows into population history and trait architecture // Nat. Rev. Genet. 2018. V. 19. № 4. P. 220–234. https://doi.org/10.1038/nrg.2017.109
Frazer K.A., Ballinger D.G., Cox D.R. et al. International HapMap Consortium A second generation human haplotype map of over 3.1 million SNPs // Nature. 2007. V. 449(7164). P. 851–861. https://doi.org/10.1038/nature06258
Karafet T.M., Bulayeva K.B., Bulayev O.A. et al. Extensive genome-wide autozygosity in the population isolates of Daghestan // Eur. J. Hum. Genet. 2015. V. 23. № 10.P. 1405–1412. https://doi.org/10.1038/ejhg.2014.299
Колесников Н.А., Харьков В.Н., Зарубин А.А., Степанов В.А. Особенности распределения регионов высокой гомозиготности в геномах представителей коренного населения Северной Евразии // Генетика. 2019. Т. 55. № 10. С. 1231–1236. https://doi.org/10.1134/S0016675819100072
Purcell S., Neale B., Todd-Brown K. et al. PLINK: A tool set for whole-genome association and population-based linkage analyses // Am. J. Hum. Genet. 2007. V. 81. P. 559–575. https://doi.org/10.1086/519795
Глазунова Е.О., Харьков В.Н., Раджабов М.О. и др. Генофонд коренных народов Дагестана цезской группы по маркерам Y-хромосомы // Мед. генетика. 2016. Т. 15. № 4. С. 29–31.https://doi.org/10.1234/XXXX-XXXX-2016-4-29-31
Гаджиев А.Г. Антропология малых популяций Дагестана. Махачкала: Даг. филиал АН СССР, 1971. 368 с.
Zhabagin M., Balanovska E., Sabitov Z. et al. The connection of the genetic, cultural and geographic landscapes of transoxiana // Scientific Reports. 2017. V. 7. № 1. P. 3085. https://doi.org/10.1038/s41598-017-03176
Харьков В.Н., Степанов В.А., Медведева О.Ф. и др. Происхождение якутов: анализ гаплотипов Y-хромосомы // Мол. биология. 2008. Т. 42. № 2. С. 226–237.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Структура генофонда тувинцев по маркерам Y‑хромосомы // Генетика. 2013. Т. 49. № 12. С. 1416–1425. https://doi.org/10.7868/S0016675813120035
Колесников Н.А., Харьков В.Н., Раджабов М.О. и др. Различия по уровню инбридинга в популяциях Дагестана: анализ регионов высокой гомозиготности // Мед. генетика. 2020. Т. 19. № 7(216). С. 21–22.
Bulaeva K.B., Davudov O.M., Pavlova T.A. et al. Genetic subdivision of Dagestan ethnic populations // Genetika. 2003. V. 39. № 1. P. 83–92.
Bulayeva K.B. Overview of genetic-epidemiological studies in ethnically and demographically diverse isolates of Dagestan, Northern Caucasus, Russia // Croat. Med. J. 2006. V. 47. № 4. P. 641–648.
Деренко М.В., Малярчук Б.А., Возняк М. и др. Разнообразие линий Y-хромосомы у коренного населения Южной Сибири // Докл. РАН. 2006. Т. 411. № 2. С. 273–277.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Генофонд бурят: клинальная изменчивость и территориальная подразделенность по маркерам Y-хромосомы // Генетика. 2014. Т. 50. № 2. С. 203–213. https://doi.org/10.7868/S0016675813110088
Дополнительные материалы отсутствуют.