

Генетика, 2021, T. 57, № 12, стр. 1450-1457
Полногеномное исследование связи метилирования CpG-сайтов с агрессивным поведением
С. А. Боринская 1, *, А. В. Рубанович 1, **, А. К. Ларин 2, А. В. Казанцева 3, Ю. Д. Давыдова 3, Э. В. Генерозов 2, Э. К. Хуснутдинова 3, Н. К. Янковский 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
2 Федеральный научно-клинический центр физико-химической медицины
Федерального медико-биологического агентства
119435 Москва, Россия
3 Институт биохимии и генетики Уфимского научного центра Российской академии наук
450054 Уфа, Россия
* E-mail: borinskaya@vigg.ru
** E-mail: rubanovich@vigg.ru
Поступила в редакцию 02.08.2021
После доработки 09.08.2021
Принята к публикации 10.08.2021
Аннотация
Профиль метилирования ДНК может отражать как генетические особенности, так и внешние воздействия, которые влияют на ряд признаков человека, включая характеристики поведения, в том числе антисоциальное поведение. Для исследования связи метилирования CpG-сайтов с агрессивным поведением был определен уровень метилирования 800К-сайтов в ДНК, выделенной из периферической крови у 122 мужчин, совершивших убийства, и у 163 мужчин контрольной группы. Для 25 топовых по эффекту CpG-сайтов (p < 1E-22) ассоциация с агрессивным поведением воспроизводились в тестовой выборке и оставалась значимой на уровне эпигенома после поправки на сопутствующие факторы (возраст, курение, национальность). Среди факторов, ассоциированных с генами, с которыми связаны выявленные CpG-сайты, представлены: неблагоприятные условия в детстве, курение, злоупотребление алкоголем и астма, что соответствует ранее опубликованным данным по факторам, связанным с физической агрессией. Полученные результаты вносят вклад в понимание биологических основ агрессии и выявление биомаркеров агрессивного поведения.
Агрессивное поведение представляет одну из важных социальных проблем. По данным Вcемирной оргaнизaции здрaвоохрaнения, убийства и умышленное причинение тяжкого вреда здоровью входит в ведущие причины cмертноcти во вcем мире для людей в возрасте 15–44 лет [1]. В России наиболее высокий уровень убийств наблюдался в 1990-е–2000-е годы, достигая в отдельные периоды 28–30 на 100 000 населения, но к 2018–2020 гг. снизился до 5.4–5.8 на 100 000 человек [2, 3].
Исследования показывают, что агрессивное поведение, в том числе экстремально агрессивное, ведущее к совершению серийных убийств, имеет наследственную составляющую [4–9], при этом влияние наследственности, оцененное в близнецовых исследованиях и исследованиях приемных детей, составляет около 50% [6, 10].
Средовые воздействия, такие как ряд заболеваний или стресс, или курение матери во время беременности, неблагоприятные условия воспитания в детстве и другие могут приводить к повышению риска формирования асоциального, в том числе агрессивного поведения. Особенно значимы воздействия, перенесенные в пренатальный и постнатальный периоды, которые, как показано для человека и модельных животных, могут вызывать долговременные эпигенетические модификации активности генов, связанных с регуляцией поведения, в том числе за счет изменения уровня метилирования специфичных CpG-сайтов [11–17]. Показано, для некоторых признаков предикция на основе уровня метилирования ассоциированных с признаком эпигенетических маркеров более эффективна, чем на основе полигенного показателя риска [18, 19]. Изменения уровня метилирования CpG-сайтов, связанные с агрессивным поведением, показаны как в исследованиях отдельных генов, так и в полногеномом анализе ассоциаций эпигенетических маркеров (EWAS, epigenome-wide association studies) (см. обзор и ссылки в [17]). Уровень агрессии в этих исследованиях определяется по преступным деяниям, либо опросникам, оценивающим проявления агрессивного поведения. Однако биомаркеры агрессивного поведения, в том числе экстремальной физической агрессии, проявляющейся как умышленное убийство, которые воспроизводились бы от исследования к исследованию, до настоящего времени не выявлены [17, 20], поэтому поиск их представляется актуальным.
В данной работе мы провели полногеномное исследование ассоциации уровня метилирования CpG-сайтов в ДНК периферической крови с агрессивным поведением у мужчин, совершивших убийство, в сравнении смужчинами контрольной популяционной выборки.
МАТЕРИАЛЫ И МЕТОДЫ
В исследовании были использованы образцы венозной крови лиц, совершивших убийства и госпитализированных для психиатрического обследования в 2016–2018 гг. Все образцы крови были собраны у неродственных индивидов под контролем биоэтического комитета ИБГ УФИЦ РАН с соблюдением процедуры “информированного согласия” и сбором данных о месте рождения, национальной принадлежности, курении, потреблении алкоголя и социоэкономических характеристиках. В группу лиц с агрессивным поведением (совершивших убийства) вошло 134 мужчины в возрасте 40.9 ± 13.4 лет – русских, татар, башкир, удмуртов и других национальностей. Контрольная группа представлена образцами: ДНК из ранее собранных популяционных коллекций лаборатории анализа генома ИОГен РАН и коллекций биологических материалов человека ИБГ УНЦ РАН и включала 163 мужчин в возрасте 46.1 ± 9.4 – лет русских, татар, башкир, удмуртов и др. ДНК выделена из лимфоцитов периферической крови методом фенольно-хлороформной экстракции.
Анализ паттернов метилирования проводился согласно методике производителя с помощью чипа InfiniumMethylationEPICBeadChipKit (Illumina, США) (EPIC 850K), который обеспечивает определение уровня метилирования 863904 CpG-сайтов, покрывающих весь геном [21, 22]. Вслед за экспериментальным определением уровня метилирования проводили нормализацию данных с контролем качества определения метилирования по каждому сайту, контролем качества образца (исключаются образцы с большим количеством сайтов с некачественно определенным уровнем метилирования), нормализацией значений для коррекции технических ошибок, нормализация значений для коррекции групповых эффектов (чип/чип корреляция) согласно процедурам, описанным в [22]. Из общего количества сайтов, для которых были получены оценки уровней метилирования (beta) с использованием описанного выше метода нормализации данных, после контроля качества метилирования по сайтам и образцам осталось 823 843 CpG-сайтов, а четыре образца не прошли контроль качества.
Исследуемые образцы были разделены на две группы – обучающая выборка (52 мужчин с агрессивным поведением и 126 в контрольной группе) и подтверждающая выборка (82 и 37 образцов соответственно). Обучающая и подтверждающая выборки были независимыми и анализировались в различные периоды исследования. Значимость различий уровней метилирования CpG-сайтов у убийц по сравнению с контролем оценивалась по непараметрическому тесту Манна–Уитни. Анализ возможных эффектов сопутствующих факторов (возраст, курение, национальность) проводился по методу ANOVA, реализованному в программе SPSS v. 12.
Фенотипы, сопряженные с изменениями паттернов метилирования генов, к которым относятся выявленные CpG-сайты, приведены согласно открытой базе данных Атлас EWAS [23], который с одержит описания 288 974 CpG в 35 594 генах.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
При анализе метилома с помощью чипа EPIC 850 K у мужчин с агрессивным поведением в сравнении с мужчинами контрольной выборки в обучающей выборке было обнаружено 19 876 CpG-сайтов, уровень метилирования которых отличался со статистической значимостью выше порогового согласно поправке на множественное тестирование (поправка Бонферрони (0.05/823 843 = 6.1Е–9)). Из них 25 топовых по эффекту CpG-сайтов (p < 1E–22) уверенно воспроизводились в тестовой выборке (табл. 1, рис. 1). Влияние сопутствующих факторов (возраст, курение, национальность) оказалось незначимым (табл. 2).
Таблица 1.
Топовые по эффекту CpG-сайты, уровень метилирования которых ассоциирован с агрессивным поведением
| Название CpG-сайта | Ген | Локализация | Фенотипы | Значимость различий | |
|---|---|---|---|---|---|
| обучающая выборка | тестовая выборка | ||||
| cg20450977 | MTRNR2L8 | Тело гена, вне островка | Неблагоприятные условия в детстве, ИМТ, бесплодие | 1.9E-44 | 1.0E-47 |
| cg24159721 | RP11-206L10.17 | То же | Fibroblasts | 8.3E-44 | 5.5E-48 |
| cg27559706 | DHFR | Промотер, шельф | Курение, синдром Дауна | 1.5E-33 | 1.5E-36 |
| cg15890734 | DHFR | Тело гена, шельф | То же | 4.0E-32 | 2.9E-46 |
| cg11628021 | – | – | – | 4.9E-32 | 1.5E-03 |
| cg05001044 | RP11-206L10.17 | Тело гена, вне островка | Возраст, болезнь Крейтцфельдта–Якоба, синдром Дауна | 6.7E-32 | 7.7E-34 |
| cg11894324 | RP5-857K21.5 | Тело гена, шельф | – | 2.3E-30 | 2.2E-39 |
| cg16581032 | – | – | – | 2.2E-29 | 6.9E-48 |
| cg17759706 | ANKRD11 | Тело гена, шельф | Неблагоприятные условия в детстве, возраст, алкогольная зависимость, курение, астма, ИМТ | 4.8E-28 | 1.6E-05 |
| cg16340030 | STX3 | Тело гена, вне островка | Астма, синдром Дауна | 7.3E-28 | 1.3E-10 |
| cg16267121 | MTRNR2L3 | Промотер, шельф | Астма, колоректальный рак | 1.5E-27 | 2.7E-46 |
| cg23106865 | MTRNR2L6 | Промотер, вне островка | То же | 1.7E-27 | 5.8E-08 |
| cg22400896 | DENND5A | Тело гена, вне островка | Возраст, курение, лейкемия | 3.2E-27 | 1.2E-05 |
| cg11330075 | hsa-mir-1977, RP5-857K21.4 | Тело гена, шельф | – | 8.8E-27 | 1.2E-45 |
| cg05812299 | MTRNR2L5 | Промотор, шельф | Астма, колоректальный рак | 2.9E-25 | 4.7E-23 |
| cg25191519 | SPG7 | Тело гена, шельф | Неблагоприятные условия в детстве, возраст, алкогольная зависимость, курение, астма, ИМТ, синдром Дауна | 3.3E-25 | 1.3E-05 |
| cg27573888 | C1orf38 | Тело гена, вне островка | – | 1.6E-23 | 2.0E-05 |
| cg27237039 | SNRNP48 | Тело гена, шельф | Возраст, синдром Дауна | 3.2E-23 | 2.6E-03 |
| cg03228760 | SS18L1 | То же | Возраст, курение, алкогольная зависимость | 4.1E-23 | 1.1E-09 |
| cg14215390 | PHF20 | Промотер, берег | Возраст, курение | 5.1E-23 | 1.9E-03 |
| cg12376350 | hsa-mir-1977, RP5-857K21.4 | Тело гена, шельф | – | 1.1E-22 | 4.9E-38 |
| cg22914762 | MTRNR2L13 | Промотор, вне островка | Болезнь Бехчета, рецептор фактора некроза опухоли | 1.1E-22 | 4.9E-15 |
| cg23127585 | – | Шельф | Синдром Шегрена | 3.6E-22 | 6.1E-06 |
| cg26634885 | PLCD3 | Промотор, островок | Шизофрения, возраст, курение, астма, синдром Дауна | 5.7E-21 | 7.4E-07 |
| cg03838322 | ZNF326 | Тело гена, шельф | Высокое потребление насыщенных жирных кислот, инсулиновая резистентность | 5.8E-21 | 1.1E-04 |
Рис. 1.
Примеры гипер- и гипометилирования CpG-сайтов у убийц по сравнению с контролем. А – агрессивное поведение, К – популяционный контроль, индекс “1” маркирует тестовую выборку, оси абсцисс – исследованные выборки.
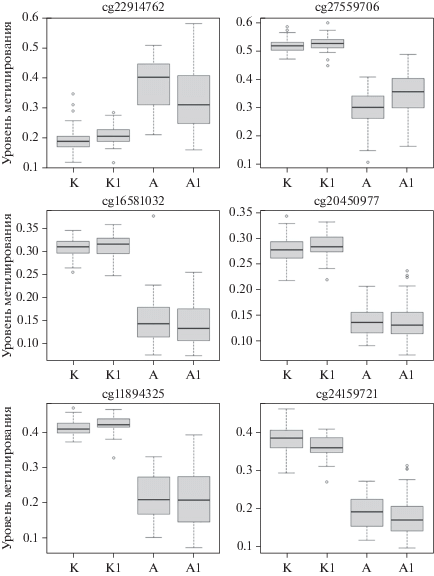
Таблица 2.
Значимость влияния ковариат (возраст, курение, национальность) на различия уровней метилирования между группами с агрессивным поведением и контролем
| Значимость действия фактора (р-значения) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Факторы | статус | возраст | курение | национальность | ||||
| Сравнения: | А-К | А1-К1 | А-К | А1-К1 | А-К | А1-К1 | А-К | А1-К1 |
| cg20450977 | 3.4E-56* | 7.8E-41* | 0.901 | 0.432 | 0.079 | 0.743 | 0.478 | 0.113 |
| cg24159721 | 9.5E-54* | 3.3E-39* | 0.576 | 0.056 | 0.007* | 0.563 | 0.859 | 0.069 |
| cg27559706 | 6.9E-64* | 1.7E-23* | 0.006* | 0.410 | 0.185 | 0.016* | 0.627 | 0.425 |
| cg15890734 | 3.5E-54* | 9.9E-36* | 0.386 | 0.141 | 0.964 | 0.514 | 0.129 | 0.123 |
| cg11628021 | 4.3E-27* | 4.0E-03* | 0.358 | 0.562 | 0.464 | 0.748 | 0.694 | 0.273 |
| cg05001044 | 3.0E-62* | 6.1E-22* | 0.294 | 0.240 | 0.765 | 0.959 | 0.938 | 0.770 |
| cg11894324 | 2.5E-58* | 1.9E-26* | 0.088 | 0.212 | 0.239 | 0.606 | 0.312 | 0.684 |
| cg16581032 | 2.0E-51* | 5.8E-36* | 0.428 | 0.277 | 0.299 | 0.459 | 0.637 | 0.383 |
| cg17759706 | 1.8E-18* | 6.9E-06* | 0.418 | 0.779 | 0.492 | 0.683 | 0.709 | 0.632 |
| cg16340030 | 2.5E-12* | 8.5E-06* | 0.306 | 0.423 | 0.292 | 0.263 | 0.612 | 0.789 |
| cg16267121 | 3.2E-45* | 4.6E-33* | 0.035 | 0.235 | 0.220 | 0.644 | 0.167 | 0.346 |
| cg23106865 | 1.0E-18* | 1.1E-04* | 0.767 | 0.959 | 0.273 | 0.778 | 0.210 | 0.029 |
| cg22400896 | 2.9E-12* | 5.5E-05* | 0.736 | 0.120 | 0.116 | 0.156 | 0.446 | 0.294 |
| cg11330075 | 1.4E-46* | 3.8E-32* | 0.736 | 0.254 | 0.571 | 0.747 | 0.779 | 0.519 |
| cg05812299 | 4.9E-38* | 1.4E-15* | 0.487 | 0.050 | 0.435 | 1.0E-04* | 0.196 | 0.008* |
| cg25191519 | 4.2E-17* | 1.5E-04* | 0.548 | 0.859 | 0.392 | 0.100 | 0.501 | 0.073 |
| cg27573888 | 2.5E-15* | 3.2E-05* | 0.517 | 0.307 | 0.546 | 0.075 | 0.229 | 0.708 |
| cg27237039 | 3.2E-11* | 2.0E-03* | 0.501 | 0.145 | 0.828 | 0.405 | 0.687 | 0.129 |
| cg03228760 | 2.3E-18* | 3.4E-07* | 0.050 | 0.935 | 0.964 | 0.907 | 0.168 | 0.990 |
| cg14215390 | 2.2E-14* | 9.0E-03* | 0.590 | 0.218 | 0.379 | 0.114 | 0.574 | 0.550 |
| cg12376350 | 1.5E-27* | 5.3E-27* | 0.760 | 0.191 | 0.026* | 0.328 | 0.275 | 0.068 |
| cg22914762 | 8.5E-33* | 7.4E-10* | 0.719 | 0.084 | 0.769 | 0.001* | 0.054 | 0.002* |
| cg23127585 | 2.2E-11* | 1.3E-04* | 0.322 | 0.425 | 0.229 | 0.751 | 0.636 | 0.316 |
| cg26634885 | 3.5E-09* | 7.5E-06* | 0.226 | 0.146 | 0.251 | 0.047* | 0.737 | 0.169 |
| cg03838322 | 3.9E-14* | 2.0E-03* | 0.005 | 0.999 | 0.345 | 0.905 | 0.560 | 0.943 |
Согласно описанию CpG-сайтов чипа EPIC 850K Infinium Methylation EPIC v1.0 B5 Manifest File (https://support.illumina.com/downloads/infinium-methylationepic-v1-0-product-files.html) 22 из выявленных нами сайтов расположены в теле, шельфе или в промотрном участке 20 структурных генов и генов микроРНК, тогда как для трех сайтов связь с генами неизвестна. Согласно информации, представленной в атласе EWAS [23], метилирование других CpG-сайтов в генах, с которыми связаны выявленные нами CpG-сайтов, ассоциировано с курением и возрастом (по восьми случаям среди наших 25 сайтов), с неблагоприятными условиями в детстве и злоупотреблением алкоголя (по трем генам), с астмой (сайты в семи генах), столько же с синдромом Дауна и один сайт в гене, ассоциированном с шизофренией. Для того, чтобы оценить обогащенность выявленной группы генов генами с какими-либо определенными функциями, мы сравнили частоту встречаемости ассоциированных с генами фенотипов в выявленной нами группе и в целом в атласе EWAS (табл. 3). В выявленной группе генов в два раза чаще, чем в среднем по атласу EWAS представлены гены, ассоциированные с неблагоприятными условиями в детстве, курением, потреблением алкоголя и астмой. Изменение уровня метилирования аналогичных групп генов (ассоциированных с неблагоприятными условия в детстве, курением и потреблением алкоголя и иммунными функциями) выявлено в опубликованных исследованиях связи метилирования с агрессивным поведением [17, 24]. В настоящем исследовании указанное превышение частоты было статистически значимым лишь для астмы (p = 0.019); для неблагоприятных условий в детстве, курения и потребления алкоголя превышения частот оказались на уровне тенденции (0.05 < p < 0.07). Для остальных выявленных фенотипов частота встречаемости близка к средней частоте встречаемости в атласе EWAS.
Таблица 3.
Частота встречаемости фенотипов, ассоциированных с генами, с которыми связаны выявленные CpG-сайты
| Фенотип | Данное исследование | Атлас EWAS | p-value | ||
|---|---|---|---|---|---|
| количество ассоцииро-ванных генов | частота | количество ассоцииро-ванных генов | частота | ||
| Неблагоприятные условия в детстве | 3 | 0.188 | 3348 | 0.094 | 0.057 |
| Курение | 8 | 0.500 | 12 603 | 0.354 | 0.072 |
| Потребление алкоголя | 3 | 0.188 | 3273 | 0.092 | 0.053 |
| Возраст | 8 | 0.500 | 14 296 | 0.402 | 0.145 |
| Астма | 7 | 0.438 | 8352 | 0.235 | 0.019 |
| Синдром Дауна | 7 | 0.438 | 12 062 | 0.339 | 0.137 |
| Шизофрения | 1 | 0.063 | 2846 | 0.080 | 0.369 |
В исследованиях эпигенетических маркеров, ассоциированных со средовыми воздействиями, такими как курение [25] или неблагоприятными условиями в детстве [26], наблюдается воспроизводимость большинства связанных с этими признаками эпигенетических маркеров от исследования к исследованию. Однако для агрессивного поведения такие устойчиво воспроизводимые маркеры пока не выявлены [17, 20, 24]. Выявленные в данном исследовании маркеры, ассоциированные с экстремально агрессивным поведением (убийством), необходимо подтвердить в дальнейших исследованиях на независимых выборках. Поиск биомаркеров агрессивного поведения, как указывающих на причинные изменения, лежащих в основе связанных с агрессией нейроэндокринных, иммунных и метаболических нарушений, так и маркеров, отражающих сопряженные влияния окружающей среды, соответствует общей стратегии развития мер предотвращения асоциального поведения, включающей как мониторинг семейных и социоэкономических факторов, так и медико-биологических [20, 27–30].
Работа выполнена в рамках мероприятия 5 “Разработка методики определения статуса генетических локусов и прижизненной модификации участков ДНК, влияющих на психоэмоциональный статус человека” научно-технической программы Союзного государства “Разработка инновационных геногеографических и геномных технологий идентификации личности и индивидуальных особенностей человека на основе изучения генофондов регионов Союзного государства” (“ДНК-идентификация”) при финансовой поддержке Министерства науки и образования РФ (Государственный контракт № 011-17 от 26.09.2017).
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
World Health Organization. Preventing suicide: A global imperative. World Health Organization. 2014. https://apps.who.int/iris/handle/10665/131056
Богданов С.В., Репецкая А.Л. Убийства в России и США: сравнительный анализ криминальной статистики // Криминол. журн. Байкальского гос. ун-та эконом. права. 2009. № 4. С. 13–22.
ЕМИСС [Электронный ресурс]. Режим доступа: https://www.fedstat.ru. (дата обращения: 05.07.2021).
Craig I.W., Halton K.E. Genetics of human aggressive behavior // Hum. Genet. 2009. V. 126. № 1. P. 101–113. https://doi.org/10.1007/s00439-009-0695-9
Pavlov K.A., Chistiakov D.A., Chekhonin V.P. Genetic determinants of aggression and impulsivity in humans // J. Appl. Genet. 2012. V. 53. № 1. P. 61–82. https://doi.org/10.1007/s13353-011-0069-6
Plomin R., DeFries J.C., Knopik V.S., Neiderhiser J. Behavior Genetics (6th ed.). N.Y.: Worth Publ., 2013. 560 p.
Tiihonen J., Rautiainen M., Ollila H.M. et al. Genetic background of extreme violent behavior // Mol. Psychiatry. 2015. V. 20. № 6. P. 786–792. https://doi.org/10.1038/mp.2014.130
Лазебный О.Е., Куликов А.М., Бутовская П.Р. и др. Анализ агрессивного поведения с помощью 250 SNP-маркеров у молодых русских мужчин // Генетика. 2020. Т. 56. № 9. С. 1075–1086. https://doi.org/10.31857/S0016675820080093
Драгович А.Ю., Боринская С.А. Генетическая и геномная основа агрессивного поведения человека // Генетика. 2019. Т. 55. № 12. С. 1381–1396.
Waltes R., Chiocchetti A.G., Freitag C.M. The neurobiological basis of human aggression: A review on genetic and epigenetic mechanisms // Am. J. Med. Genet. Part B. Neuropsychiatric Genetics. 2015. V. 171. № 5. P. 650–675. https://doi.org/10.1002/ajmg.b.32388
Martin E.M., Fry R.C. Environmental influences on the epigenome: Exposure-associated DNA methylation in human populations // Annu Rev. Public Health. 2018. V. 39. P. 309–333. https://doi.org/10.1146/annurevpublhealth-040617-014629
Tielbeek J.J., Johansson A., Polderman T.J.C. et al. Genome-wide association studies of a broad spectrum of antisocial behavior // JAMA Psychiatry. 2017. V. 74. № 112. P. 1242. https://doi.org/10.1001/jamapsychiatry.2017.3069
Chistiakov D.A., Chekhonin V.P. Early-life adversity-induced long-term epigenetic programming associated with early onset of chronic physical aggression: Studies in humans and animals // World J. Biol. Psychiatry. 2019. V. 20. № 4. P. 258–277. https://doi.org/10.1080/15622975.2017.1322714
Dintsova V.V., Roetman P.J., Ip H.F. et al. Genomics of human aggression: Current state of genome-wide studies and an automated systematic review tool // Psychiatr. Genet. 2019. V. 29. № 5. P. 170–190. https://doi.org/10.1097/YPG.0000000000000239
Ip H.F., van der Laan C.M., Brikell I. et al. Genetic Association Study of Childhood Aggression across raters, instruments and age // Transl. Psychiatry. 2021. V. 11. № 1. P. 413. https://doi.org/10.1038/s41398-021-01480-x
van Dongen J., Hagenbeek F.A., Suderman M. et al. DNA methylation signatures of aggression and closely related constructs: A meta-analysis of epigenome-wide studies across the lifespan // Mol. Psychiatry. 2021. V. 26. P. 1–15. https://doi.org/10.1038/s41380-020-00987-x
Shah S., Bonder M.J., Marioni R.E. et al. Improving phenotypic prediction by combining genetic and epigenetic associations // Am. J. Hum. Genet. 2015. V. 97. № 1. P. 75–85. https://doi.org/10.1016/j.ajhg.2015.05.014
McCartney D.L., Hillary R.F., Stevenson A.J. et al. Epigenetic prediction of complex traits and death // Genome Biol. 2018. V. 19. № 1. P. 136. https://doi.org/10.1186/s13059-018-1514-1
Мустафин Р.Н., Казанцева А.В., Еникеева Р.Ф. и др. Эпигенетика агрессивного поведения // Генетика. 2019. Т. 55. № 9. С. 987–997.
Mitjans M., Seidel J., Begemann M. et al. Violent aggression predicted by multiple pre-adult environmental hits // Mol. Psychiatry. 2019. V. 24. P. 1549–1564. https://doi.org/10.1038/s41380-018-0043-3
Pidsley R., Zotenko E., Peters T.J. et al. Critical evaluation of the Illumina Methylation EPIC BeadChip microarray for whole-genome DNA methylation profiling // Genome Biol. 2016. V. 17. № 1. P. 208. https://doi.org/10.1186/s13059-016-1066-1
Wu M.C., Kuan P.F. A guide to Illumina bead chip data analysis // DNA Methylation Protocols. Methods in Molecular Biology / Ed. Tost J. N.Y.: Humana Press, 2018. V. 1708. P. 303–329.
EWAS Atlas [Электронный ресурс]. Режим доступа: https://ngdc.cncb.ac.cn/ewas/atlas/index (дата обращения: 05.07.2021).
Cecil C.A.M., Walton E., Pingault J.B. et al. DRD4 methylation as a potential biomarker for physical aggression: An epigenome-wide, cross-tissue investigation // Am. J. Med. Genet. B. Neuropsychiatr Genet. 2018. V. 177(8). P. 746–764. https://doi.org/10.1002/ajmg.b.32689
Silva C.P., Kamens H.M. Cigarette smoke-induced alterations in blood: A review of research on DNA methylation and gene expression // Exp. Clin. Psychopharmacol. 2021. V. 29. № 1. P. 116–135. https://doi.org/10.1037/pha0000382
Cecil C.A.M., Zhang Y., Nolte T. Neurosci childhood maltreatment and DNA methylation: A systematic review // Biobehav. Rev. 2020. V. 112. P. 392–409. https://doi.org/10.1016/j.neubiorev.2020.02.019
Raine A. Biosocial studies of antisocial and violent behavior in children and adults: A review // J. Abnorm. Child Psychol. 2002. V. 30. № 4. P. 311–326. https://doi.org/10.1023/a:1015754122318
Fazel S., Gulati G., Linsell L. et al. Schizophrenia and violence: Systematic review and meta-analysis // PLoS Med. 2009. V. 6. № 8. P. e1000120. https://doi.org/10.1371/journal.pmed.1000120
Tuvblad C., Baker L.A. Human aggression across the lifespan: Genetic propensities and environmental moderators // Adv. in Genetics. 2011. V. 75. P. 171–214. https://doi.org/10.1016/B978-0-12-380858-5.00007-1
Ehrenreich H. The impact of environment on abnormal behavior and mental disease // EMBO Rep. 2017. V. 18. № 5. P. 661–665. https://doi.org/10.15252/embr.201744197
Дополнительные материалы отсутствуют.