

Генетика, 2021, T. 57, № 12, стр. 1403-1414
Реконструкция происхождения гыданских ненцев на основе генетического анализа их родовой структуры с помощью нового набора YSTR-маркеров
В. Н. Харьков 1, *, Л. В. Валихова 1, Е. Л. Яковлева 1, В. Н. Сереброва 1, Н. А. Колесников 1, Т. И. Петелина 2, И. Ю. Хитринская 1, В. А. Степанов 1
1 Научно-исследовательский институт медицинской генетики, Томский национальный исследовательский медицинский центр Российской академии наук
634050 Томск, Россия
2 Тюменский кардиологический научный центр, Томский национальный исследовательский медицинский центр Российской академии наук
625026 Тюмень, Россия
* E-mail: vladimir.kharkov@medgenetics.ru
Поступила в редакцию 20.05.2021
После доработки 17.06.2021
Принята к публикации 28.06.2021
Аннотация
Исследована генетическая структура ненецких родов по маркерам Y-хромосомы с использованием разработанного набора YSTR для определения популяционной принадлежности неизвестного индивида. Результаты анализа частот гаплогрупп и YSTR-гаплотипов свидетельствуют, что ненецкие рода являются родственными объединениями, в большинстве случаев имеющими одного предка по мужской линии. Показано, что Y-хромосомный генофонд ненцев структурирован по родовому принципу. Для подавляющего большинства образцов выявлена тесная генетическая близость представителей одного рода и относительно недавний эффект основателя по мужской линии. Структура гаплотипов гаплогрупп Y-хромосомы в составе ненецкого генофонда демонстрирует генетическую близость ненцев с хантами и энцами. Выявлены значительные различия между двумя ненецкими фратриями. Оценка возраста происхождения отдельных сублиний Y-хромосомы и родовых кластеров гаплотипов полностью согласуется с данными этнографии о формировании родов и фратрий в составе гыданских ненцев. Результаты сравнительного анализа спектра гаплотипов убедительно свидетельствуют о специфичности сублиний Y-хромосомы и кластеров гаплотипов не только на уровне этносов и популяций, но и на уровне фратрий, родов и отдельных фамильных групп. За единичными исключениями каждая фратрия и род имеют свой специфичный кластер гаплотипов, равноудаленных друг от друга.
Ненцы – народ, проживающий в европейской части России и севере Западной Сибири, являются одной из многочисленных групп коренных малочисленных народов. Они расселены на огромной территории, простирающейся с запада на восток на несколько тысяч километров, и делятся на европейских и азиатских (сибирских). Европейские ненцы проживают в Ненецком автономном округе Архангельской области, сибирские – в Ямало-Ненецком автономном округе Тюменской области и в Долгано-Ненецком Таймырском муниципальном районе Красноярского края. Небольшие группы ненцев проживают в Ханты-Мансийском автономном округе, в Мурманской области, а также в Республике Коми. Наряду с нганасанским, энецким и селькупским язык ненцев входит в самодийскую группу уральской языковой семьи. По антропологическим признакам ненцы относятся к монголоидам.
По данным Всероссийской переписи 2010 г. в составе населения Ямало-Ненецкого автономного округа их насчитывалось 29 772 человека (5.7% от общей численности населения) [1]. Самая восточная часть ареала ненцев расположена на территории между Обью и Енисеем. Ненцев, проживающих в Тазовском районе Ямало-Ненецкого автономного округа, можно объединить общим названием “гыданские ненцы”, так как большая часть района расположена на территории Гыданского полуострова, они относятся к группе тундровых [2]. В Тазовском районе находятся национальные поселки – Гыда, Антипаюта, Находка и районный центр Тазовский.
Как и для большинства южносибирских и западносибирских народов, отличительной чертой ненцев является наличие родов, где счет родства ведется по мужской линии. Родовая организация ненцев достаточно хорошо изучена антропологами и этнографами [1]. Браки у ненцев всегда были строго экзогамными. У гыданских, ямальских и тазовских ненцев рода объединялись в две экзогамные фратрии, одна из которых – Харючи (“настоящие ненцы”). Она состояла из родов самодийского происхождения, вторая (Вануйто, “иноплеменники”) – из родов, восходящих к аборигенному генетическому субстрату, а также энецкого и хантыйского происхождения [3]. Иноэтничные группы родов включены ненцами в свою дуально-фратриальную организацию и их представители заключают браки в соответствии с их экзогамными нормами. Со второй половины XVIII в. ненцы стали наращивать поголовье домашних оленей, что привело к постепенному увеличению численности населения и дроблению больших патриархальных родов на малые рода-патронимии [4]. Гыданская группа ненцев является популяцией, наиболее сохранившей традиционную социальную структуру, так как была наименее подвержена изменениям в социально-экономической и культурной сферах.
В XX в. в состав гыданских ненцев влились отдельные представители лесных и европейских ненцев, а также несколько русских и татар. На 2001 год на территории Тазовского района проживали представители 43 ненецких родов и восьми оненечившихся фамилий. На Гыдане проживают представители обеих фратрий. К родам фратрии Харючи относятся Адер, Евай, Лапсуй, Ненянг, Няруй, Окотэтто, Сусой, Сэротэтто, Сюгней, Тогой, Тэсида, Хабдю, Харючи, Хороля, Худи, Хэно, Ядне, Яндо, Яптунай. Во фратрию Вануйто входят рода Вануйто, Вэнго, Ламдо, Пуйко, Сабы, Яр, Яптик, Яунгад. Основу гыданского этнического массива составили ненцы родов Харючи, Яр, Сюгней, Айваседа, Адер.
В формировании восточных ненцев приняли участие также энецкие и хантыйские по происхождению рода, которые вошли в состав фратрии Вануйто. К энецким родам принадлежат Марьик, Аседа, Паровых, Тёр, Оковай. К хантыйским относятся Тибичи, Вэхо, Неркыхы, Няданги, Пурунгуй, Салиндер [5].
Родовую принадлежность у ненцев в большинстве случаев возможно установить по фамилиям, так как практически все современные ненецкие фамилии повторяют название рода либо полностью в исходной форме, либо с небольшими изменениями. Таким образом, подробные данные об этнографии и родовой структуре гыданских ненцев дают богатый материал для исследования генетической структуры родов и фратрий.
Большое число информативных SNP и STR-маркеров Y-хромосомы, используемых в настоящее время для изучения популяционно-генетической структуры различных этносов, позволяют проверить, является ли тот или иной род кровнородственным объединением, имеющим одного родоначальника по мужской линии, или осознание такой общности лишь условное и род является общностью людей, проживающих на одной территории, но не связанных генетическим родством по отцовским генеалогическим линиям. Ранее по маркерам Y-хромосомы уже была охарактеризована генетическая структура родов у казахов [6, 7], башкир [8], хакасов и шорцев [9].
В ходе выполнения проекта по разработке различных тест-систем для ДНК-идентификации и определения популяционной принадлежности неизвестного индивида по образцу его ДНК было показано, что гаплотипы, построенные с помощью генотипирования специально отобранных по уровню их информативности и темпу мутирования STR-маркеров Y-хромосомы, обладают значительной этнической и территориальной специфичностью. Была установлена очень высокая межпопуляционная дифференциация полученных YSTR-гаплотипов [10], что позволило создать эффективно работающий набор YSTR-маркеров для целей ДНК-идентификации.
Цель исследования – охарактеризовать генетическую структуру родов гыданских ненцев с использованием маркеров Y-хромосомы, определить их генетическое родство по мужской линии и установить межэтническое генетическое родство отдельных родов, входящих в состав различных фратрий. В качестве инструмента исследования мы использовали разработанный нами набор YSTR для определения популяционной принадлежности индивидов по образцу их ДНК, решая параллельно задачу тестирования возможностей набора для уточнения степени точности и детализации привязки индивидов не только к этносам и популяциям, но и к конкретному роду и фамилии.
МАТЕРИАЛЫ И МЕТОДЫ
Материал исследования составили образцы ДНК мужчин из различных локальных популяций гыданских ненцев (N = 322). Выборку гыданских ненцев представляет население поселка Тазовский, села Антипаюта, села Гыда, Тазовского района Ямало-Ненецкого автономного округа Российской Федерации. Материал получен в ходе совместных научно-практических медицинских экспедиций в 2019 г. и депонирован в биоресурсную коллекцию “Биобанк населения Северной Евразии”. Забор первичного биологического материала (венозной крови) у доноров проводили с соблюдением процедуры письменного информированного согласия на проведение исследования. На каждого донора составлялась анкета с его родословной, указанием этнической принадлежности и мест рождения предков. В исследование включены только образцы ДНК доноров-мужчин, по результатам анкетирования отрицавших факт метисации по отцовской линии с представителями других этносов минимум в трех поколениях. Индивида относили к данной этнической группе на основании его собственной этнической идентификации, его родителей, места рождения и принадлежности к определенному роду.
Среди всех мужчин, включенных в настоящее исследование, оказались представлены следующие рода (в алфавитном порядке): Адер, Вануйто, Вэлло, Вэнго, Евай, Лапсуй, Лар, Марьик, Ненянг, Няч, Оковай, Окотэтто, Пуйко, Пурунгуй, Салиндер, Сусой, Тёр, Тибичи, Тогой, Тэсида, Худи, Ядне, Яндо, Яптик, Яптунай, Яр, Яунгат.
Для изучения состава и структуры гаплогрупп Y-хромосомы в исследование были включены две системы генетических маркеров: диаллельные локусы, представленные SNP, и полиаллельные высоковариабельные микросателлиты (YSTR). С помощью 103 SNP-маркеров определяли принадлежность образцов к той или иной гаплогруппе. Классификация гаплогрупп дана в соответствии с предложенной Консорциумом по исследованию Y-хромосомы [11], с последующими изменениями [12]. Анализ STR-гаплотипов внутри гаплогрупп проводили с использованием 45 микросателлитных маркеров нерекомбинирующей части Y-хромосомы (YSTR) (DYS19, 385a, 385b, 388, 389I, 389II, 390, 391, 392, 393, 426, 434, 435, 436, 437, 438, 439, 442, 444, 445, 448, 449, 456, 458, 460, 461, 481, 504, 505, 518, 525, 531, 533, 537, 552, 570, 576, 635, 643, YCAIIa, YCAIIb, GATA H4.1, Y-GATA-A10, GGAAT1B07). Генотипирование SNP-маркеров проводили с помощью ПЦР и последующего анализа фрагментов ДНК с применением ПДРФ как описано ранее [10]. STR-маркеры генотипировали с помощью капиллярного электрофореза на генетическом анализаторе ABI Prism 3730 и программного обеспечения GeneMapper как описано ранее [10]. Экспериментальные исследования проведены на базе Центра коллективного пользования научно-исследовательским оборудованием “Медицинская геномика” Томского НИМЦ РАН). Построение медианных сетей гаплотипов Y-хромосомы проводили с использованием программы Network v 10.0.0.0 (Fluxus Technology Ltd.; www.fluxus-engineering.com) по методу медианных сетей Бандельта [13]. Оценку возраста генерации наблюдаемого разнообразия гаплотипов в гаплогруппах проводили методом ASD [14], на основании средних квадратичных отличий в числе повторов между всеми маркерами. Длина поколения принималась равной 30 годам, темп мутирования 0.0021 на локус на поколение. При расчете возраста родов по отдельным гаплогруппам исключались единичные образцы, которые значительно выбивались из общего кластера гаплотипов. Расчеты проводили для родовых групп численностью не менее пяти образцов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате анализа частоты встречаемости использованных SNP-маркеров в исследуемой выборке гыданских ненцев было выявлено шесть гаплогрупп Y-хромосомы. За исключением единичных образцов, практически все исследованные образцы относятся к Y-хромосомной макрогруппе N. Все шесть оставшихся образцов относятся к различным западно-евразийским по происхождению гаплогруппам, что, возможно, свидетельствует о единичных фактах недавней метисации или включения в состав родов отдельных представителей пришлого населения (табл. 1) (рис. 1).
Таблица 1.
Частоты встречаемости гаплогрупп Y-хромосомы у гыданских ненцев
| Род | N1a2b1b1-B170,B171, Z35101 (xB172) | N1a2b1b1b-B172, Z35108 | N1a2b 2a-E- Y3185 | N1a2b1-B478, Z35080 (xVL65) | N1a1a1a1 a2a1c-Y13850 | N1a1a1 a1a2-Z1936 | Другие | Суммарное число |
|---|---|---|---|---|---|---|---|---|
| Адер | 3 (100%) |
– | – | – | – | – | – | 3 |
| Вануйто | – | 46 (100%) |
– | – | – | – | – | 46 |
| Вэлло | 6 (100%) |
– | – | – | – | – | – | 6 |
| Вэнго | 1 (14.29%) |
– | 4 (57.14%) |
– | – | – | 2 (28.57%) |
7 |
| Евай (Явай) | 9 (100%) |
– | – | – | – | – | – | 9 |
| Лапсуй | 37 (94.87%) |
– | – | – | 2 (5.13%) |
– | – | 39 |
| Лар | – | – | – | – | 2 (100%) |
– | – | 2 |
| Марьик | – | – | – | 3 (100%) |
– | – | – | 3 |
| Ненянг | 4 (100%) |
– | – | – | – | – | – | 4 |
| Няч | – | – | – | 6 (100%) |
– | – | – | 6 |
| Оковай | – | – | – | 1 (100%) |
– | – | – | 1 |
| Окотэтто | 2 (66.67%) |
1 (33.33%) |
– | – | – | – | – | 3 |
| Пуйко | – | 2 (100%) |
– | – | – | – | – | 2 |
| Пурунгуй | – | 2 (66.67%) |
– | – | – | 1 (33.33%) |
– | 3 |
| Салиндер | – | – | – | – | 34 (89.47%) |
– | 4 (10.53%) |
38 |
| Сусой | 7 (100%) |
– | – | – | – | – | – | 7 |
| Тёр | – | – | – | 1 (100%) |
– | – | – | 1 |
| Тибичи | 2 (25%) |
– | – | – | 6 (75%) |
– | – | 8 |
| Тогой | 4 (66.67%) |
– | – | – | 2 (33.33%) |
– | – | 6 |
| Тэсида | 11 (100%) |
– | – | – | – | – | – | 11 |
| Худи | 3 (100%) |
– | – | – | – | – | – | 3 |
| Ядне | 35 (87.5%) |
2 (5%) |
– | – | 3 (7.5%) |
– | – | 40 |
| Яндо | 14 (100%) |
– | – | – | – | – | – | 14 |
| Яптик | 1 (25%) |
– | – | 3 (75%) |
– | – | – | 4 |
| Яптунай | 20 (100%) |
– | – | – | – | – | – | 20 |
| Яр | 7 (20%) |
1 (2.86%) |
11 (31.43%) |
– | 2 (5.71%) |
14 (40%) |
– | 35 |
| Яунгат | – | 1 (100%) |
– | – | – | – | – | 1 |
| 166 | 55 | 15 | 14 | 51 | 15 | 6 | 322 |
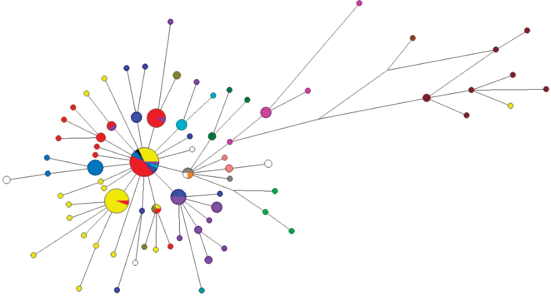
Рис. 1.
Медианная сеть гаплогруппы N1a2b1b1-B170,B171,Z35101 (xB172). Желтым цветом обозначен род Лапсуй, красным – Ядне, светло-голубым – Яр, темно-голубым – Яптик, синим – Тэсида, темно-синим – Яндо, фиолетовым – Яптунай, светло-зеленым – Адер, темно-зеленым – Ненянг, болотным – Тогой, белым – Окотэтто, серым – Худи, светло-розовым – Вэлло, розовым – Сусой, коричневым – Евай, оранжевым – Вэнго.
Фратрия Харючи
В формировании ненцев приняли участие в основном два компонента: южносибирский самодийский и местный аборигенный. В результате их взаимодействия, по мнению Б. Долгих [15], сформировались две фратрии ненцев. Одна восходит к самодийцам – Харючи, а другая к аборигенам – Вануйта (или Вануйто).
Практически все прогенотипированные образцы, относящиеся к фратрии Харючи, относятся (за единичными исключениями) к гаплогруппе N1a2b1b1-B170,B171,Z35101(xB172). Это представители родов Адер, Евай (Явай), Лапсуй, Ненянг, Окотэтто, Сусой, Тогой, Тэсида, Худи, Ядне, Ядно и Яптунай. Все представители рода Вэлло, который относится к лесным ненцам, также принадлежат к этой гаплогруппе, хотя он не входит в состав фратрии Харючи (рис. 1).
Медианная сеть YSTR-гаплотипов демонстрирует ярко выраженную звездообразную филогению гаплогруппы N1a2b1b1-B170, свидетельствующую о сильном эффекте основателя и общем предке для всех родов по мужской линии. Исключением являются все представители рода Евай, которые формируют отдельный кластер гаплотипов. Этому возможно два объяснения: во-первых, род Евай может иметь более древнего общего предка с остальными родами фратрии Харючи и являться потомками параллельно существовавшей с ней группы мужчин; во-вторых, возможно, что различия по числу тандемных повторов сразу в нескольких маркерах являются результатом нескольких одновременных мутаций при гаметогенезе конкретного индивида, который и был родоначальником этого рода. Существование таких выбивающихся из общей сети гаплотипов отдельных субветвей в различных гаплогруппах было показано и ранее, при анализе большого массива данных по различным популяциям и гаплогруппам [16]. С учетом небольшой дисперсии числа повторов в этом кластере гаплотипов у рода Евай наиболее вероятен именно второй вариант. Согласно данным этнографов род Евай выделился из рода Адер в первой половине XX в. [2, 5]. Ветвь гаплотипов рода Евай на медианной сети наиболее близка именно к кластеру, состоящему из представителей родов Адер, Сусой, Вэнго и Ненянг.
Модальный гаплотип этой гаплогруппы обнаружен у представителей семи ненецких родов. При этом все модальные гаплотипы всех крупных родов отличаются от модального гаплотипа гаплогруппы N1a2b1b1-B170 как минимум на один мутационный шаг. Только шесть гаплотипов кроме общего модального повторяются более чем у одного рода. Все они отличаются от модального на один мутационный шаг, и в силу его большой доли от общего числа гаплотипов могут быть результатом повторных параллельных или обратных мутаций отдельных STR. За единичными исключениями каждый род характеризуется собственным спектром гаплотипов, в среднем равноудаленных от модального. Каждый отдельный род также имеет ярко выраженную генетическую близость принадлежащих к нему индивидов и звездообразную филогению гаплотипов на медианной сети, что подтверждает биологическое родство мужчин данного рода по отцовской линии (рис. 2–5).
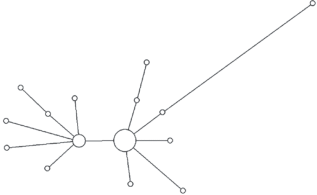
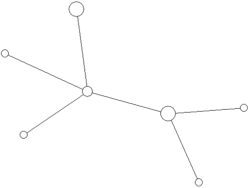
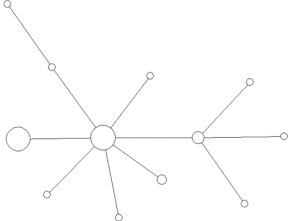
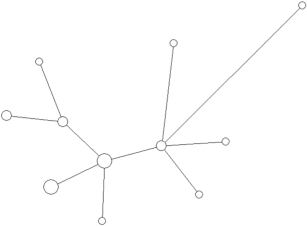
Таким образом, подавляющее число мужчин родов фратрии Харючи принадлежит к одной гаплогруппе и является потомками одного общего предка, что подтверждает данные этнографии о родстве различных родов и выделении их из общих предковых групп. Рода этой фратрии являются патрилинейно родственными друг другу, и в ходе увеличения численности и расселения по новым территориям, за единичными исключениями, не ассимилировали и не включали в свой состав мужчин из других родов. Полученные результаты полностью соответствуют данным о доминировании самодийского исходного компонента во фратрии Харючи и данным этнографии о генеалогической близости всех входящих в нее родов.
Общий возраст для всего массива гаплотипов основной гаплогруппы N1a2b1b1-B170 составил 724 года (SD = 167 лет), для рода Лапсуй – 395 лет (SD = 202 года), для рода Вэлло – 805 лет (SD = = 313 лет), для рода Евай – 814 лет (SD = 266 лет).
Для многих родов, входящих во фратрию Харючи, по различным документам (переписям населения, спискам крестьян и т.д.) известно примерное время их отделения от рода Харючи. Для рода Сусой время генерации гаплотипов составляет 104 года (SD = 73 года). Род Сусой является подразделением рода Окотэтто и по данным этнографии он обособился в первой половине XX в. [5]. Возраст рода Ядне – 285 лет (SD = 146 лет). Родоначальником Ядне является Яур Удолин, который упоминается в ревизских документах в составе рода Харючи начиная с 1763 г. [2, 5]. Возраст рода Яндо – 366 лет (SD = 189 лет). Этот род впервые был отмечен переписью 1897 г. До этого времени сведений о нем не имеется. Однако представители этого рода утверждают, что Яндо является одним из самых древних ненецких родов. Возраст рода Яптунай – 476 лет (SD = 207 лет). Название этого рода впервые упоминается в списках крестьян в конце 30-х гг. XIX в. Возраст рода Тэсида – 100 лет (SD = 74 года). В официальных документах этот род впервые отмечен во второй половине XIX в. Таким образом, результаты оценки возраста генерации гаплотипического разнообразия для перечисленных родов хорошо совпадают с данными этнографии и записей о их обособлении.
Общий возраст гаплогруппы N1a2b1b1-B170 близок по времени к приходу основной миграционной волны самодийцев на территорию северной части Западной Сибири.
Фратрия Вануйто
Во фратрии Вануйто наблюдается совершенно другой состав гаплогрупп и значительная их специфика у разных родов по сравнению с Харючи. В то время как все до одного представителя родов Вануйто (N = 46), Пуйко (N = 2) и Яунгат (N = 1) принадлежат к гаплогруппе N1a2b1b1b-B172, Z35108 (рис. 6), большинство членов рода Вэнго (N = 4) относятся к другой линии – N1a2b2a-E-Y3185 (рис. 7), а рода Яптик – к гаплогруппе N1a2b1-B478-Z35080xVL65 (N = 3) (рис. 8) и N1a2b1b1-B170 (N = 1) (рис. 1).
Рис. 6.
Медианная сеть гаплогруппы N1a2b1b1b-B172. Зеленым цветом обозначен род Вануйто, фиолетовым – Яр, розовым – Пуйко, синим – Вэлло, желтым – Пурунгуй, красным – Вэнго, темно-зеленым – Яунгат.
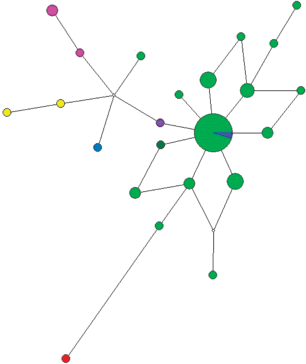
Рис. 7.
Медианная сеть гаплогруппы N1a2b2a-E-Y3185. Синим цветом отмечен род Яр, зеленым – Вэнго, оранжевым – Тайбери, фиолетовым – Тибичи.
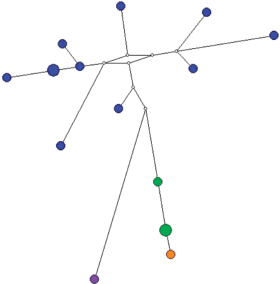
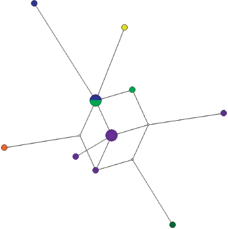
Рис. 8.
Медианная сеть гаплогруппы N1a2b1-B478,Z35080(xVL65). Фиолетовым цветом обозначен род Няч, синим – Яптик, зеленым – Марьик, оранжевым – Тёр, желтым – Оковай.
Досамодийский аборигенный элемент в составе ненцев представлен главным образом в рамках фратрии Вануйто. Ее название Б. Долгих [15] связывал со словом ванг – яма и предполагал, что оно распространялось на “ямных жителей” – жителей землянок, т.е. на аборигенов, вошедших затем в состав фратрии, дав ей свое имя. Сначала название закрепилось за аборигенами, позже став именем фратрии, восходящей по происхождению к аборигенному пласту. Принадлежность большинства представителей части родов этой фратрии к гаплогруппе N1a2b1b1b-B172, а не к параллельной ей N1a2b1b1-B170 хорошо согласуется с этим предположением. При этом наличие других гаплогрупп свидетельствует о большем генетическом разнообразии фратрии Вануйто по сравнению с Харючи и возможно большей гетерогенности аборигенного компонента, вошедшего в состав современных гыданских ненцев.
Для основного рода Вануйто возраст составил 308 лет (SD = 94 года). Общий возраст для всего массива гаплотипов основной гаплогруппы N1a2b1b1b-B172,Z35108 составил 811 лет (SD = = 211 лет), что практически совпадает с возрастом N1a2b1b1-B170 и вероятно свидетельствует о начале демографического роста ненцев после ассимиляции пришлыми самодийцами аборигенного населения Севера. При этом, судя по всему, не происходило вытеснения коренных мужчин и их вклад в генофонд современных гыданских ненцев весьма значителен.
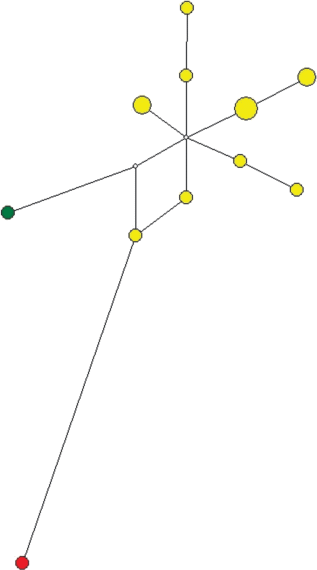
Род Яр. К фратрии Вануйто этнографы также относят род Яр – единственный род тазовских ненцев, из числа самых ранних переселенцев в низовья Таза. Сведений о его происхождении по данным этнографии нет. Из всех исследованных в рамках настоящей работы родов Яр является самым гетерогенным по составу. Представители этого рода относятся к пяти гаплогруппам Y-хромосомы: N1a1a1a1a2-Z1936 (N = 14) (рис. 10), N1a2b2a-E-Y3185 (N = 11) (рис. 7), N1a2b1b1-B170 (N = 7) (рис. 1), N1a1a1a1a2a1c-Y13850 (N = 2) (рис. 9) и N1a2b1b1b-B172 (N = 1) (рис. 6). Даже если исключить единичные образцы двух последних гаплогрупп, то этот род разделяется на три различных по происхождению генетических компонента. Внутри каждой гаплогруппы наблюдается значительное разнообразие гаплотипов и характерная для всех других родов звездообразная филогения медианной сети, что свидетельствует об отсутствии недавней метисации с другими родами или о включении в состав отдельных пришлых мальчиков или мужчин среди представителей этих кластеров гаплотипов. Возможно, что такая гетерогенность объясняется одинаковым наименованием разных по происхождению родов по топонимам, приписыванию общего названия для родов, проживающих на одной территории, или ассимиляции и включении в свой состав мужчин из местного населения при расселении на тазовской территории исходного рода Яр. Для гаплогруппы N1a1a1a1a2-Z1936 возраст составил 551 год (SD = 196 лет). Для гаплогруппы N1a2b1b1-B170 возраст составил 419 лет (SD = 283 года).
Рис. 9.
Медианная сеть гаплогруппы N1a1a1a1a2a1c-Y13850. Оранжевым цветом обозначен род Тибичи, светло-зеленым – Салиндер, темно-зеленым – Лар, синим – Ядне, красным – Яр.
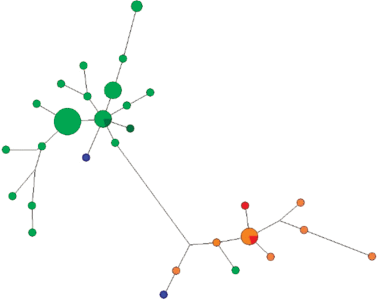
Рис. 10.
Медианная сеть гаплогруппы N1a1a1a1a2-Z1936. Желтым цветом обозначен род Яр, зеленым – Пурунгуй, красным – Аседа.
Хантыйские рода. В начале XX в. стали селиться на Тазу и представители хантыйских по происхождению родов. Они сформировались в XVIII–XIX вв. в низовьях Оби в результате развития ненецкого крупнотабунного оленеводства и вовлечения в него части северных хантов. К этим родам относятся Лар, Няданги, Неркыхы, Пурунгуй, Салиндер и Тибичи [17].
Мужчины, представляющие самый многочисленный род Салиндер (N = 38), полностью принадлежат к совершенно другой гаплогруппе N1a1a1a1a2a1c-Y13850. К этой же гаплогруппе относятся все представители рода Лар (N = 2) (рис. 9). Род Тибичи разделяется на N1a1a1a1a2a1c-Y13850 (N = 6) и N1a2b1b1-B170 (N = 2). Лишь в роде Пурунгуй не выявлена эта основная для хантыйских родов гаплогруппа: два образца принадлежат к гаплогруппе N1a2b1b1b-B172 (рис. 6) и один к N1a1a1a1a2-Z1936 (рис. 10). Гаплогруппа N1a1a1a1a2a1c-Y13850 также обнаружена нами и в выборке казымских хантов, из северной части их ареала.
Общий возраст для всего массива гаплотипов основной гаплогруппы N1a1a1a1a2a1c-Y13850 составил 1752 года (SD = 517 лет), для рода Салиндер – 843 года (SD = 383 года), для рода Тибичи – 607 лет (SD = 342 года). Общий возраст этой гаплогруппы у гыданских ненцев может быть несколько завышен, поскольку между разными родами наблюдаются значительные различия размера аллелей по некоторым YSTR-маркерам, которые могли возникнуть в результате нескольких совместных мутаций у отдельного индивида или накопиться за два–три поколения его потомков.
Данные по гаплогруппам Y-хромосомы полностью подтверждают хантыйское происхождение этих родов, их отличие от фратрии Харючи, других родов фратрии Вануйто и генетическое родство друг другу.
Энецкие рода. Марьик, Няч, Оковай и Тёр, относящиеся к родам энецкого происхождения, полностью состоят из представителей отдельной гаплогруппы N1a2b1-B478,Z35080(xVL65) (рис. 8). Гаплотипы этой линии также образуют звездообразную филогению, с относительно небольшой дисперсией аллелей, что свидетельствует об их тесном генетическом родстве и относительно недавнем общем происхождении.
Вытеснение энцев ненцами из междуречья Таза и Енисея началось с двух “больших” военных столкновений между ними во второй половине XVII в. В следующем столетии ненцы продолжали проникновение на восток для освоения завоеванных территорий, а к XIX в. окончательно закрепились в низовьях Таза и Енисея. Оставшиеся на Тазу энцы были ассимилированы ненцами, а энецкие рода включены в ненецкую двухфратриальную систему. Они стали относиться к фратрии Вануйто, в которую входят потомки аборигенного населения и различные иноплеменники, в отличие от фратрии Харючи, состоящей из потомков самодийцев, выходцев из Южной Сибири [4].
Полученные результаты позволяют более точно взглянуть на историю формирования гыданских ненцев, с одной стороны, на самом современном уровне генотипирования Y-хромосомных маркеров, с другой стороны, полностью учитывая родовую структуру народа, анализируя межэтнические взаимосвязи и родовые различия.
По результатам генотипирования удалось выявить в подавляющем большинстве родов основную гаплогруппу, представителей которой среди исследованной выборки образцов этого рода абсолютное большинство. Генотипирование широкого набора YSTR-маркеров позволило получить достаточно детальные и специфичные кластеры гаплотипов, что позволяет с высокой степенью достоверности определить не только этническую и субпопуляционную принадлежность образцов, но и в большинстве случаев определить их принадлежность к конкретным роду и фамилии. Практически каждый род гыданских ненцев характеризуется полным доминированием одной гаплогруппы со специфичным спектром гаплотипов, подчеркивающим недавний эффект основателя. Анализ гаплотипов убедительно показал, что представители одного рода являются родственниками по мужской линии и имеют родоначальника, жившего в относительно недалеком прошлом. Таким образом, генофонд представителей восточной группы ненецкого этноса, а точнее часть, маркируемая гаплогруппами Y-хромосомы, действительно структурирована прежде всего по родовому принципу. Именно этот уровень организации генофонда гыданских ненцев как единой системы является наиболее точно характеризующим его популяционную структуру. Две фратрии гыданских ненцев значительно различаются по генетической структуре родов по маркерам Y-хромосомы, что подтверждает их формирование на основе различных предковых компонент. Ненецкие рода хантыйского и энецкого происхождения по составу Y-хромосомных гаплогрупп полностью отличаются друг от друга и от самодийской по происхождению фратрии Харючи и аборигенной Вануйто.
Тестирование разработанного набора YSTR для определения этнической принадлежности на примере ненцев продемонстрировало его высокую эффективность и достоверность не только при определении популяционной принадлежности, но и более точную детализацию при работе с широкими и репрезентативными выборками для отдельных родовых и фамильных групп. Это позволяет точнее и эффективнее устанавливать привязку образцов мужской ДНК и открывает значительные перспективы для применения этого набора YSTR в реальной практике.
Работа выполнена в рамках Научно-технической программы Союзного государства “Разработка инновационных геногеографических и геномных технологий идентификации личности и индивидуальных особенностей человека на основе изучения генофондов регионов Союзного государства” (“ДНК-идентификация”), Государственный контракт № 011-17 от 26.09.2017, а также поддержана грантом РФФИ № 18-29-13045 “Популяционная геномика и транскриптомика человека: поиск сигналов не-нейтральной эволюции”.
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Волжанина Е.А. Демографический облик ненцев ямала по материалам всероссийских переписей XXI в. // Вестник археологии, антропологии и этнографии. 2017. № 3(38). С. 120–130.
Квашин Ю.Н. Этнические и демографические процессы у тазовских ненцев в начале XXI в. // Вестник археологии, антропологии и этнографии. 2012. № 3(18). С. 141–152.
Хомич Л.В. Проблемы этногенеза и этнической истории ненцев. Л.: Наука, Ленингр. отд-е, 1976. 187 с.
Васильев В.И. Проблема формирования енисейских ненцев (к вопросу об этнической природе этнографических групп в составе современных народностей Севера) // Этногенез и этническая история народов Севера. М.: Наука, 1975. 89 с.
Квашин Ю.Н. Основные элементы родовой структуры гыданских ненцев // Вестник археологии, антропологии и этнографии. 2001. № 3. С. 1–4.
Abilev S., Malyarchuk B., Derenko M. et al. The Y-chromosome C3*Star-Cluster attributed to Genghis Khan’s Descendants is present at hight frequency in the Kerey Clan from Kazakhstan // Hum. Biol. 2012. V. 84. № 1. P. 79–89. https://doi.org/10.3378/027.084.0106
Zhabagin M.K., Dibizova H.D., Frolova S.A. et al. The relation between the Y-chromosomal variation and the clan structure: The geen pool of the steppe aristocracy and the steppe clergy of the Kazakhs // Moscow Univ. Anthropol. Bul. 2014. V. 1. P. 96–101.
Балановская Е.В., Юсупов Ю.М., Схаляхо Р.А. и др. Генетические портреты семи кланов северо-западных башкир: вклад финно-угорского компонента в генофонд башкир // Вестник Моск. ун-та. Серия XXIII. Антропология. 2017. № 3. С. 94–103.
Харьков В.Н., Новикова Л.М., Штыгашева О.В. и др. Генофонд хакасов и шорцев по маркерам Y-хромосомы: общие компоненты и генетическая структура родов // Генетика. 2020. Т. 56. № 7. С. 826–833. https://doi.org/10.31857/S0016675820070073
Харьков В.Н., Зарубин А.А., Вагайцева К.В. и др. Y‑хромосома как инструмент для ДНК-идентификации и определения популяционной принадлежности // Генетика. 2020. Т. 56. № 9. С. 1065–1074. https://doi.org/10.31857/S0016675820090118
The Y-Chromosome Consortium. A nomenclature system for the tree of human Y-chromosmal binaryhaplogroups // Genome Res. 2002. V. 12. P. 339–348.
Karafet T.M., Mendez F.L., Meilerman M.B. et al. Newbinary polymorphisms reshape and increase resolutionof the human Y chromosomal haplogroup tree // Genome Res. 2008. V. 18. P. 830–838.
Bandelt H.J. Median-joining networks for inferring intraspecific phylogenies // Mol. Biol. Evol. 1999. V. 16. P. 37–48.
Sengupta S., Zhivotovsky L.A., King R. et al. Polarity and temporality of high resolution Y-chromosome distributions in India identify both indigenous and exogenousexpansions and reveal minor genetic influence of central Asian pastoralists // Am. J. Hum. Genet. 2006. V. 78. P. 202–221.https://doi.org/10.1086/499411
Долгих Б.О. Очерки по этнической истории ненцев и энцев. М.: Наука, 1970. 268 с.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Генофонд бурят: клинальная изменчивость и территориальная подразделенность по маркерам Y-хромосомы // Генетика. 2014. Т. 50. № 2. С. 203–213. https://doi.org/10.7868/S0016675813110088
Квашин Ю.Н. Гыданские ненцы: история формирования современной родовой структуры (XVIII–XX вв.). М.: ИЭА РАН, 2003. 186 с.
Дополнительные материалы отсутствуют.