

Генетика, 2022, T. 58, № 2, стр. 177-186
Филогеография Lacerta media Lantz et Cyrén, 1920 (Lacertidae: Sauria) по результатам анализа митохондриального гена цитохрома b
М. А. Доронина 1, И. В. Доронин 1, *, **, С. А. Луконина 2, Л. Ф. Мазанаева 3, А. В. Барабанов 1
1 Зоологический институт Российской академии наук
199034 Санкт-Петербург, Россия
2 Пензенский государственный университет
440026 Пенза, Россия
3 Дагестанский государственный университет
367025 Махачкала, Россия
* E-mail: igor.doronin@zin.ru
** E-mail: ivdoronin@mail.ru
Поступила в редакцию 24.05.2021
После доработки 24.08.2021
Принята к публикации 07.09.2021
- EDN: ZEVCTU
- DOI: 10.31857/S0016675822020035
Аннотация
Проведен анализ cytb митохондриальной ДНК (1143 пн) у представителей Lacerta media. Проанализированы 34 образца из 19 локалитетов. Особое внимание уделено ранее неисследованным популяциям на северной границе ареала в пределах Северного Кавказа и Западного Закавказья. Реконструировано два типа дендрограмм филогенетических отношений, показавших идентичную топологию – байесовская (BA) и максимального правдоподобия (ML). Филогенетическая реконструкция выделяет четыре поддержанные клады. Аналогично на медианной сети гаплотипов обособляются четыре гаплогруппы. На более ранних этапах процесса филогенеза L. m. media выделяются популяции восточной части Армянского нагорья и Северного Загроса. В дальнейшем происходит обособление популяций центральной части Армянского нагорья, Большого Кавказа, а на завершающем этапе – Центрального Загроса и восточной части Малоазиатского нагорья. Полученные данные не поддерживают ранее предложенный Ф. Ахмадзаде с соавт. сценарий расселения номинативного подвида с запада на восток и северо-восток. Взамен его мы рассматриваем противоположное направление – с востока на север, юг и запад. Выявлено две географические области, где обитают представители нескольких клад/гаплогрупп – Армянское нагорье и горы Загроса.
Средняя ящерица, Lacerta media Lantz et Cyrén, 1920 – представитель надвидового комплекса Lacerta (trilineata), находящегося в фокусе исследований филогении и филогеографии рода Lacerta Linnaeus, 1758 [1]. В ряде публикаций последних лет [2–6] популяции с территории Кавказа, где обитает номинативный подвид (по месту сбора лектотипа – долина р. Квабисхеви, окр. г. Боржоми Грузии [7]), были представлены выборкой лишь из пяти локалитетов с Армянского нагорья. Однако не были исследованы ящерицы с Северного Кавказа и Западного Закавказья – Дагестана и Краснодарского края, т.е. с северной границы видового ареала [8, 9]. В ряде работ эта территория вовсе отсутствует на картах распространения L. media [2, 3]. Обозначенные дагестанские и черноморские периферические популяции изолированы от закавказских и представляют интерес с позиции исследования микроэволюционных процессов.
В настоящее время отсутствует ясность с определением времени появления и расселения представителей комплекса Lacerta (trilineata). Обнаружение в северокавказском позднемиоценовом местонахождении “Солнечнодольск” ископаемых костных фрагментов L. trilineata [10] ставят под сомнение существующие сценарии филогенеза комплекса, рассматривающие в качестве областей дивергенции основных филогенетических клад Малоазиатское нагорье (Анатолию) и район Эгейского моря в плиоцене–раннем плейстоцене [3, 6] или в позднем миоцене–раннем плиоцене [5]. В отношении L. media также имеется противоречивая информация: на основе сведений по палеогеографии региона время вселения средней ящерицы в горный Дагестан относят к сармату (поздний миоцен) [11], а в Южную Осетию – к концу миоцена–середине плиоцена [12], тогда как по молекулярно-генетическим датировкам вид появился примерно 3.5 млн лет назад в позднем плиоцене [4, 6] или примерно 7.12 млн лет назад в позднем миоцене [5].
Вышесказанное свидетельствует о необходимости более обстоятельного исследования филогении и филогеографии средней ящерицы; на Кавказе мы можем ожидать обнаружения рефугиума, свидетельствующего о древней колонизации региона, либо подтверждение относительно недавнего расселения вида с территории Анатолии – предполагаемого центра ее происхождения и радиации. Подчеркнем, что палеонтологические свидетельства, которые могут служить реперными точками при реконструкции филогении вида, ограничиваются упоминанием средней ящерицы в мустьерских слоях (верхний плейстоцен) пещеры Швалиети в Грузии [13].
Немаловажным является то обстоятельство, что L. m. media включена в региональные [14, 15] и национальные [16, 17] списки охраняемых таксонов животных (Красные книги); это подчеркивает актуальность данного исследования с позиции изучения и сохранения генетического разнообразия.
МАТЕРИАЛЫ И МЕТОДЫ
Для выделения геномной ДНК использовали части регенерированных хвостов или фаланги пальцев передних конечностей ящериц, фиксированные в 96%-ном этаноле, от пяти экземпляров, собранных в трех локалитетах Большого Кавказа и одном локалитете Загроса (рис. 1). Ваучерные экземпляры хранятся в коллекции Зоологического института РАН (ZISP) и Зоологическом музее МГУ (ZMMU). В филогенетический анализ были включены 29 последовательностей L. m. media [2, 3], взятых из GenBank (http:// www.ncbi.nlm.nih.gov/gene/) (табл. 1).
Рис. 1.
Пункты сбора экземпляров Lacerta media, использованных для молекулярно-генетического анализа. Обозначение клад: А – желтые, B – синие, C – красные, D – зеленые. Нумерация и буквенное обозначение соответствуют таковым в табл. 1.
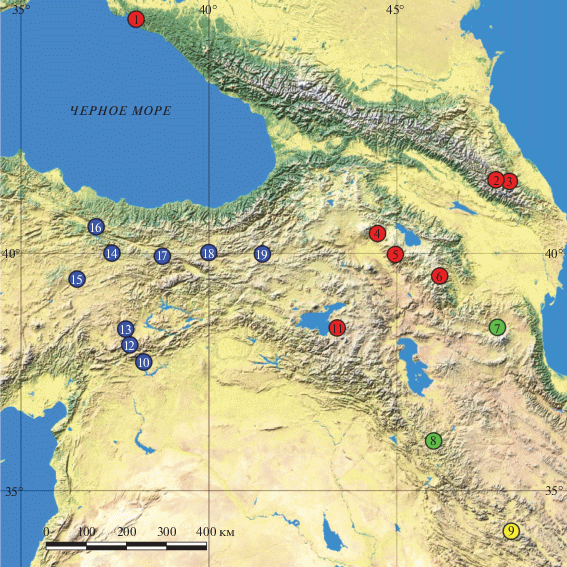
Таблица 1.
Коллекционные экземпляры Lacerta media, использованные при молекулярно-генетическом анализе
| Номера локалитетов на рис. 1 | Локалитеты | Коллекционные номера | Номера в Генбанке | Клада | Даты сбора | Географические координаты | Высота, м н. у. м. | Коллектор(-ы) | Источник информации |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Россия, Краснодарский край, окр. г. Геленджик, Ю склон Маркотхского хребта | ZISP 23571.1-2 | MZ065557–MZ065558 | C | 28.04.2006 | 38.0624, 44.5992 | 160 | С.В. Островских, К.Д. Мильто | Данная публикация |
| 2 | Россия, Дагестан, Ахтынский р-н, с. Кака | ZISP TS 2979 | MZ065555 | C | 07.07.2020 | 47.6328, 41.4790 | 1130 | З.С. Исмаилова | » |
| 3 | Россия, Дагестан, Ахтынский р-н, окр. с. Ахты, склон Ю экспозиции г. Ухиндаг | ZMMU 13972 | MZ065556 | C | 03–04.05.2013 | 47.6916, 41.4478 | 1450 | А.А. Аскендеров | » |
| 4 | Армения, Арагацотнская обл., гора Аралер (=Аралер, Ара лер, Ара, Араилер) | – | U88603 | C | – | 44.4833, 40.4000 | 2150 | – | [2] |
| 5 | Армения, Араратская обл., заповедник “Хосровский лес” | – | KC896976 | C | – | 44.9579, 39.9763 | 1500 | – | [4] |
| 6 | Армения, Сюникская обл., трасса М-2 между с. Шаки и г. Горис | – | KC896977 | C | – | 46.1359, 39.5358 | 2010 | – | » |
| 7 | Иран, Ардебиль, с. Араблу | – | KC896978–KC896983 | D | – | 47.6720, 38.4753 | 1110 | – | » |
| 8 | Иран, Курдистан, 35 км ЮЗ г. Секкес, около трассы Секкес–Бане | ZISP 29894 | MZ065554 | D | 08.08.2018 | 45.9746, 36.0727 | 2070 | А.Н. Барабанов | Данная публикация |
| 9 | Иран, Лурестан, окр. г. Нурабад | – | KC896984– KC896988 | A | – | 48.0397, 34.1097 | 1800 | – | [4] |
| 10 | Турция, Адыяман, г. Адыяман | – | KC897002– KC897003 | B | – | 38.2667, 37.7500 | 640 | – | [4] |
| 11 | Турция, Ван, окр. г. Ван | – | KC896993, KC896995 | C | – | 43.4107, 38.4624 | 1810 | – | » |
| 12 | Турция, Малатья, СВ г. Доганшехир | – | KC896989 | B | – | 37.8947, 38.1093 | 1180 | – | » |
| 13 | Турция, Малатья, перевал Карахан | – | KC896990– KC896991 | B | – | 37.7855, 38.3393 | 1870 | – | » |
| 14 | Турция, Сивас, дорога Доганшар–Хафик | – | KC897000 | B | – | 37.4197, 40.0029 | 1660 | – | » |
| 15 | Турция, Сивас, окр. д. Казанджик | – | KC897001 | B | – | 36.4942, 39.4717 | 1630 | – | » |
| 16 | Турция, Токат, между г. Никсар и г. Башчифтлик | – | KC896994 | B | – | 37.0004, 40.5374 | 640 | – | » |
| 17 | Турция, Эрзинджан, д. Кюрелик | – | KC896996– KC896998 | B | – | 38.7608, 39.9397 | 1730 | – | » |
| 18 | Турция, Эрзинджан, окр. д. Кючюкотлукбели | – | KC896999 | B | – | 39.9939, 40.0147 | 2070 | – | » |
| 19 | Турция, Эрзурум, 20 км СВ г. Эрзурум | – | KC896992 | B | – | 41.4220, 39.9864 | 2020 | – | » |
Выделение тотальной ДНК проводили стандартным солевым методом с лизированием протеиназой К [18]. Амплификацию участка гена cytb осуществляли с использованием ПЦР-амплифиикатора T100TM Thermal Cycler (Bio-Rad) и праймеров 5'-GCC CCA AAA TAA GGA GAT GG-3' и 5'-TAG TGA TGG GGG ATT AAA GC-3' [19, 20] по следующей схеме: начальная денатурация 95°C (3 мин), затем 32 цикла (95°C – 30 с, 56°С – 30 с, 72°С – 90 с) и конечная элонгация (72°С – 5 мин). Реакционная смесь для ПЦР (25 мкл) содержала 50–100 нг ДНК, 0.5 мкМ каждого праймера, 0.2 мМ dNTPs, 1.5 мМ MgCl2, 2.5 мкл 10× ПЦР буфера (10 мМ Tris-HCl, pH 8.3, 50 мМ KCl) и 2 ед. Taq-полимеразы (Thermo Scientific). Секвенирование проводили на генетическом анализаторе ABI PRIZM 3500xL (Applied Biosystems). Окончательная длина амплифицированного фрагмента ДНК составила 1143 пн. Полученные последовательности (MZ065554–065558) депонировали в GenBank NCBI.
Выравнивание последовательностей проводили в программах Geneious Prime 2021.0.1 (https:// www.geneious.com) и Ali View 1.6 [21]. Филогенетические отношения были реконструированы с использованием метода Байеса (Bayesian analysis, BA) в программе Mr. Bayes 3.1.2 [22–24] и методом максимального правдоподобия (Maximum Likelihood, ML) в программном пакете MEGA X [25]. Для BA использовали 5 млн генераций; статистическую надежность узлов ML-деревьев оценивали путем бутстреп-анализа (1500 псевдореплик). Выбор модели эволюции для нуклеотидных последовательностей проводили в программе MrModeltest 2.4 [26] c использованием информационного критерия Акаике (AIC). Была выбрана модель GTR (General Time Reversible) с параметрами I = = 0.3909 и G = 0.7440. Графические изображения деревьев получены с помощью программы FigTree 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree). Построение сети гаплотипов осуществляли методом TCS в программе PopART [27].
Для изучения внутривидовой структуры L. media в анализ были включены последовательности из работы Ф. Ахмадзаде с соавт. [3] – L. m. ciliciensis Schmidtler, 1975 (KC896967–896974), L. m. israelica Peters, 1964 (KC896975) и L. m. wolterstorffi Mertens, 1922 (KC897005–897007). Для укоренения дендрограмм филогенетических отношений в качестве внешней группы были выбраны последовательности L. strigata Eichwald, 1831 (LN835024.1), L. agilis Linnaeus 1758 (AY616285, AY616398) и Eremias stummeri Wettstein, 1940 (NC029878.1) из GenBank. При анализе генетической изменчивости с использованием программы DnaSP v.5.10.01 [28] вычисляли следующие параметры: общее количество полиморфных позиций (S), общее количество замен (η), количество гаплотипов (H), разнообразие гаплотипов (h), разнообразие нуклеотидов на сайт (π), среднее количество нуклеотидных замен (K), значение теста Таджимы (Tajima’s D) и тест на нейтральность Фу (Fu’s Fs). Генетические дистанции (p‑дистанции) вычисляли в программе MEGA X.
Для проведения анализа таксономических границ по молекулярно-генетическим признакам нами были использованы алгоритмы ASAP (Automated simultaneous analysis phylogenetics) [29] и ABGD (Automatic barcode gap discovery) [30]. Они позволяют выявить условные молекулярные операционные таксономические единицы (Molecular phylogenetic taxonomic units, MOTU), представляющие собой монофилетические клады неопределенного ранга. Анализ проводили с использованием следующих параметров: Pmin (минимальная априорная дистанция) = 0.01, Pmax (максимальная априорная дистанция) = 0.1, X (относительная ширина интервала) = 0.5; была использована матрица попарных К80-дистанций.
РЕЗУЛЬТАТЫ
34 последовательности гена cytb L. m. media образуют 24 гаплотипа. Выявлены 54 (4.7% от общей длины фрагмента) вариабельные позиции, из них – 44 (3.8%) парсимони информативные. Соотношение транзиций к трансверсиям составило 4.84. Нуклеотидный состав участка на L-цепи смещен в сторону уменьшения доли гуанина (A – 26.7, T – 31.3, C – 30, G – 12%). Показатели генетического разнообразия в выборке: S = 54, η = 57, h = 0.964 (±0.021), π = 0.0160 (±0.0010), K = = 14.01, Tajima’s D = 0.02 (p > 0.1), Fu’s Fs = –1.80 (p > 0.1). Показатель Fs-тест незначим. Недостаточность значительных отрицательных величин D свидетельствует об отсутствии в прошлом внезапного роста популяции (экспансии) и/или положительного отбора, а положительных – об отсутствии недавнего сокращения численности популяции (т.н. бутылочного горлышка), фрагментации и/или балансирующего отборa.
По результатам реконструкции филогенетических взаимоотношений представителей L. m. media были получены дендрограммы с идентичной топологией, на которых можно выделить четыре митохондриальные клады (рис. 2). Последовательности с территории Большого Кавказа вошли в кладу С, внутри которой нет значимой генетической гетерогенности (h – 0.889, K – 1.67). Для сравнения наибольшую структурированность мы наблюдаем в кладе B (h – 0.961, K – 6.08). Аналогичное прослеживается при анализе медианной сети гаплотипов: выделяются четыре гаплогруппы (I–IV), наиболее удаленная из которых – IV (рис. 3). От II группы она отдалена на 25 нуклеотидных замен, от III – на 22, а от I – на 29.
Рис. 2.
Дендрограмма филогенетических отношений представителей Lacerta media по результатам анализа фрагмента гена cytb (1143 пн). В узлах указаны байесовы апостериорные вероятности и бутстреп-поддержки (BA/BS). Буквенное обозначение клад соответствует таковому в табл. 1 и 2. В двух столбцах разными цветами указаны выделенные молекулярные операционные таксономические единицы (MOTU).

Рис. 3.
Медианная сеть гаплотипов cytb представителей Lacerta m. media (1143 пн). Каждый цветной кружок – уникальный гаплотип, чей размер пропорционален встречаемости в выборке; цвет кружка соответствует кладе на рис. 2: А – желтые, B – синие, C – красные, D – зеленые. Соединительные линии – вероятные эволюционные связи, засечки – нуклеотидные замены, черные кружки в узлах линий – предсказанные гаплотипы. Цветной заливкой обозначены географические регионы: оранжевой – Иранское нагорье, желтой – Малоазиатское нагорье, бирюзовой – Армянское нагорье, розовой – Большой Кавказ. Пунктирной линией ограничены группы гаплотипов.

При расчете p-дистанций максимальное значение было получено при сравнении клад A/D, В/D, а минимальное – B/С (табл. 2).
Таблица 2.
Средние генетические p-дистанции (%) (под диагональю) и их стандартные отклонения (над диагональю) между кладами Lacerta media по данным анализа последовательностей фрагмента гена cytb
| Клады | A | B | C | D |
|---|---|---|---|---|
| A | 0.35 | 0.38 | 0.51 | |
| B | 1.58 | 0.33 | 0.48 | |
| C | 1.63 | 1.39 | 0.49 | |
| D | 2.79 | 2.59 | 2.45 |
Анализ ASAP/ABGD показал наличие четкого разрыва между значениями популяционной и подвидовой изменчивости (treshold distance = 3.27%, barcoding gap = 2.20%). Применение данных алгоритмов позволило выявить в объеме L. media соответственно 4 и 5 монофилетических MOTU (рис. 2). Если в первом случае молекулярные операционные таксономические единицы строго соответствуют признаваемым подвидам средней ящерицы, то во втором дополнительно была делимитирована клада D.
ОБСУЖДЕНИЕ
Согласно топологии полученной дендрограммы, первой на подвидовом уровне обособляется пара L. m. israelica и L. m. wolterstorffi, в дальнейшем – L. m. ciliciensis и L. m. media. Согласно предшествующим публикациям [3, 4], Малоазиатское нагорье может рассматриваться как исходная территория, откуда происходило множественное независимое расселение средней ящерицы на Армянское нагорье (включая самые восточные турецкие популяции в районе оз. Ван), в горы Эльбурса на северо-западе Ирана и Загроса на западе Ирана; именно популяции Анатолии характеризуются наибольшим генетическим разнообразием внутри номинативного подвида. В процитированной работе [3] локалитет Араблу на северо-западе Ирана, расположенный на северном склоне потухшего вулкана Сабалан (=Савалан, Савелан или Кухе-Себелан), был отнесен к “Эльбурскому” рефугиуму. С этим мы не можем согласиться, поскольку хребет Савелан-Даг, в который входит вулкан, выступает восточной оконечностью Армянского вулканического нагорья [31]. Кроме того, L. media традиционно включают в восточносредиземноморскую зоогеографическую группу видов рептилий, в то время как Эльбурс – рефугиум гирканской герпетофауны [32]. Представители последней не обитают в районе Сабалана [33, 34].
Безусловно предковая форма L. m. media первоначально проникла на обозначенную территорию с запада, где происходила основная радиация вида [3, 4]. Однако согласно полученной нами дендрограмме (рис. 2) на более ранних этапах процесса филогенеза L. m. media выделяются популяции восточной части Армянского нагорья и Северного Загроса (клада D). В дальнейшем происходит обособление популяций центральной части Армянского нагорья и Большого Кавказа (С), а на завершающем этапе – Центрального Загроса (A) и восточной части Малоазиатского нагорья (B). Взамен сценария расселения номинативного подвида с запада на восток и северо-восток мы предлагаем противоположное направление – с востока на север, юг и запад. Наши выводы о становлении ареала L. m. media соответствуют схеме, которую предложил в 1924 г. О. Сирен [35]. Уместно вспомнить, что И.С. Даревский [36] и Т.А. Мусхелишвили [37] считали Армянское нагорье “центром формирования” средней ящерицы.
На Армянском нагорье обнаружены представители трех клад (B, С, D) и трех гаплогрупп (II, III, IV), что наглядно продемонстрировано на сети гаплотипов (рис. 3). Еще одна область, где выявлены несколько клад (А и D) и гаплогрупп (I и IV), – горы Загроса. Эти клады/гаплогруппы генетически максимально удалены друг от друга в выборке L. m. media. Обособление популяций, составляющих клады А, B и C, согласно датировкам Ф. Ахмадзаде с соавт. [3], произошло около 650 тыс. лет назад, а D – около 1 млн лет назад. Вероятно, это было обусловлено климатическими флуктуациями плейстоцена, оказавшими влияние на ландшафты этой горной территории [38] и хорологию средней ящерицы. Кладе A генетически наиболее близки гаплотипы из Адыямана, входящие в кладу B. Это отражает тесную географическую связь юго-востока Малоазиатского (Восточный Тавр) и севера-запада Иранского (Загрос) нагорий [39]. Отметим, что популяции с этой территории (остан Керманшах) были обозначены Е.Ф. Шмидтлером [40] как самостоятельная форма “L. m. media–Zagrosform”. Полученные данные по определению границ между таксонами (по алгоритму ABGD) указывают на сравнительно высокий таксономический статус популяций, полученные последовательности от которых образовали в нашем исследовании кладу D.
Обращает на себя внимание отсутствие значимой генетической дистанции в выборке с оконечностей Большого Кавказа – точки 1–3 на рис. 1, расстояние между которыми по прямой составляет примерно 850 км. Это можно интерпретировать как свидетельство быстрого расселения вида с юга в двух направлениях в обход Главного Кавказского хребта. Однако нельзя исключать и существование единого циркумкавказского ареала. Он мог распасться во время последнего ледникового (гляциального) максимума (около 22 тыс. лет назад) либо сформироваться (вторично?) в ксеротермический период голоцена (около 4500–2500 лет назад) и разделиться в связи с последующей бореализацией климата. На такой сценарий может указывать наличие изолированных северокавказских популяций другого представителя средиземноморской зоогеографической группы рептилий – оливкового полоза, Platyceps najadum (Eichwald, 1831), имеющего в регионе чрезвычайно сходные с L. media очертания ареала [41]. Снижение в этот период области вероятного распространения (пригодности местообитаний) для обитания средней ящерицы на северо-западе Большого Кавказа было показано при ГИС-моделировании [42].
В дальнейшем необходимо включить в анализ ящериц с пограничных между кладами территорий, что позволит детализировать историю расселения вида. Так, новые находки L. media в Талышских горах [43] могут свидетельствовать либо о существовании здесь малочисленных реликтовых популяций, относящихся к кладе D, либо недавнем проникновении представителей клады C в горнолесной пояс Талыша; область распространения клады B может простираться до северо-западного склона Малого Кавказа в пределах Грузии. Наконец, глубокого анализа требует изучение филогеографии вида в Загросе – все еще малоизученной территории, выступающей с одной стороны барьером между Месопотамией и нагорьем, с другой – коридором для распространения “северных” видов герпетофауны на юг региона [33].
Авторы признательны Н.Б. Ананьевой, О.А. Ермакову, В.Ф. Орловой, А.А. Кидову, Б.С. Туниеву и Н.А. Шаповалу за ценные консультации и помощь в работе.
Поддержание коллекции ZISP осуществлено в рамках проекта Министерства науки и высшего образования РФ (№ 075-15-2021-1069); исследования выполнены при финансовой поддержке гранта РНФ (№ 22-24-00079).
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Kornilios P., Thanou E., Lymberakis P. et al. A phylogenomic resolution for the taxonomy of Aegean green lizards // Zool. Scripta.2019. V. 49. № 1. P. 14–27. https://doi.org/10.1111/zsc.12385
Godinho R., Crespo E.G., Ferrand N. Harris D.J. Phylogeny and evolution of the green lizards, Lacerta spp. (Squamata: Lacertidae) based on mitochondrial and nuclear DNA sequences // Amphibia–Reptilia. 2005. V. 26. № 3. P. 271–285. https://doi.org/10.1163/156853805774408667
Ahmadzadeh F., Flecks M., Carretero M.A. et al. Rapid lizard radiation lacking niche conservatism: ecological diversification within a complex landscape // J. Biogeography. 2013. V. 40. № 9. P. 1807–1818. https://doi.org/10.1111/jbi.12121
Ahmadzadeh F., Flecks M., Rödder D. et al. Multiple dispersal out of Anatolia: biogeography and evolution of oriental green lizards // Biol. J. Linnean Society. 2013. V. 110. № 2. P. 398–408. https://doi.org/10.1111/bij.12129
Sagonas K., Poulakakis N., Lymberakis P. et al. Molecular systematics and historical biogeography of the green lizards (Lacerta) in Greece: Insights from mitochondrial and nuclear DNA // Mol. Phylogenet. Evol. 2014. V. 76. P. 144–154. https://doi.org/10.1016/j.ympev.2014.03.013
Kornilios P., Thanou E., Lymberakis P. et al. Genome-wide markers untangle the green-lizard radiation in the Aegean Sea and support a rare biogeographical pattern // J. Biogeography. 2019. V. 46. P. 552–567. https://doi.org/10.1111/jbi.13524
Доронин И.В., Доронина М.А. Обзор типовых экземпляров Lacerta media Lantz et Cyrén, 1920 (Sauria: Lacertidae) // Тр. Зоол. ин-та РАН. 2019. Т. 323. № 2. С. 85–92. https://doi.org/10.31610/trudyzin/2019.323.2.85
Nettmann H.-K., Rykena S. Lacerta trilineata Bedriaga 1886 – Riesensmaragdeidechse // Handbuch der Reptilien und Amphibien Europas. Bd. 2/I. Echsen (Sauria) II. (Lacertidae II: Lacerta) / Ed. Böhme W. Aula-Verlag (Wiesbaden), 1984. P. 100–128.
Agasyan A., Tuniyev B., Cogalniceanu D. et al. Lacerta media // The IUCN Red List of Threatened Species. 2009. (publ. online). https://doi.org/10.2305/IUCN.UK.2009.RLTS.T164749A5922769
Čerňanský A., Syromyatnikova E.V. The first Miocene fossils of Lacerta cf. trilineata (Squamata, Lacertidae) with a comparative study of the main cranial osteological differences in green lizards and their relatives // PLoS One. 2019. V. 14. № 8. (publ. online). https://doi.org/10.1371/journal.pone.0216191
Мазанаева Л.Ф., Туниев Б.С. Зоогеографический анализ герпетофауны Дагестана // Совр. герпетология. 2011. Т. 11. Вып. 1/2. С. 55–76.
Лотиев К.Ю., Туниев Б.С. Возможные пути и этапы формирования герпетофауны Южной Осетии // Горные экосистемы и их компоненты: Материалы VI Всерос. конф. с международным участием, посвященной Году экологии в России и 100-летию заповедного дела в России. Нальчик: Алеф, 2017. С. 159–160.
Векуа А.К. История животного мира – позвоночные // Грузия в антропогене. Развитие компонентов ландшафта и палеогеографические реконструкции. Тбилиси: Изд. Сакартвело, 1991. С. 308–381.
Туниев Б.С., Островских С.В. Ящерица средняя Lacerta media (Lantz et Cyren, 1920) // Красная книга Краснодарского края. Животные. III изд. Краснодар: Адм. Краснодар. края, 2017. С. 492–493.
Мазанаева Л.Ф., Аскендеров А.Д. Средняя ящерица Lacerta media (Lantz et Cyren, 1920) // Красная книга Республики Дагестан. Махачкала: Типография ИП Джамалудинов М.А., 2020. С. 532–533.
Туниев Б.С., Лотиев К.Ю. Ящерица средняя Lacerta media Lantz et Cyren, 1920 // Красная книга Республики Южная Осетия. Нальчик: Издательство М. и В. Котляровых (ООО “Полиграфсервис и Т”), 2017. С. 226–227.
Об утверждении Перечня объектов животного мира, занесенных в Красную книгу Российской Федерации. Приказ Минприроды РФ от 24.03.2020 № 162. Зарегистрировано в Минюсте РФ 02.04.2020 № 57940. https://minjust.consultant.ru/documents/45937
Aljanabi S.M., Martinez I. Universal and rapid salt-extraction of high genomic DNA for PCR-based techniques // Nucl. Acids Res. 1997. V. 25. № 22. P. 4692–4693. https://doi.org/10.1093/nar/25.22.4692
Marzahn E., Mayer W., Joger U. et al. Phylogeography of the Lacerta viridis complex: Mitochondrial and nuclear markers provide taxonomic insights // J. Zool. Syst. and Evol. Res. 2016. V. 54. № 2. P. 85–105. https://doi.org/10.1111/jzs.12115
Кукушкин О.В., Ермаков О.А., Иванов А.Ю. и др. Филогеография прыткой ящерицы в Крыму по результатам анализа гена цитохрома b: древний рефугиум на полуострове, поздняя экспансия с севера и первые свидетельства гибридизации подвидов Lacerta agilis tauridica и L. a. exigua (Lacertidae: Sauria) // Тр. Зоол. ин-та РАН. 2020. Т. 324. № 1. С. 56–99. https://doi.org/10.31610/trudyzin/2020.324.1.56
Larsson A. AliView: A fast and lightweight alignment viewer and editor for large data sets // Bioinformatics. 2014. V. 30. № 22. P. 3276–3278. https://doi.org/10.1093/bioinformatics/btu531
Huelsenbeck J., Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees // Bioinformatics. 2001. V. 17. № 8. P. 754–755. https://doi.org/10.1093/bioinformatics/17.8.754
Ronquist F., Huelsenbeck J.Ph. MrBayes 3: Bayesian phylogenetic inference under mixed models // Bioinformatics. 2003. V. 19. № 12. P. 1572–1574. https://doi.org/10.1093/bioinformatics/btg180
Altekar G., Dwarkadas S., Huelsenbeck J.P., Ronquist F. Parallel metropolis coupled markov chain Monte Carlo for Bayesian phylogenetic inference // Bioinformatics. 2004. V. 20. № 3. P. 407–415. https://doi.org/10.1093/bioinformatics/btg427
Kumar S., Stecher G., Li M. et al. MEGA X: Molecular evolutionary genetics analysis across computing platforms //Mol. Biol. Evol. 2018. V. 35. № 6. P. 1547–1549. https://doi.org/10.1093/molbev/msy096
Nylander J.A.A. Mr Modeltest version 2. Program distributed by the author. Evolutionary Biology Centre, Uppsala Univ., 2004.
Leigh J.W., Bryant D. PopART: Full-feature software for haplotype network construction // Methods Ecol. Evol. 2015. V. 6. № 9. P. 1110–1116. https://doi.org/10.1111/2041-210X.12410
Librado P., Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data // Bioinformatics. 2009. V. 25. № 11. P. 1451–1452. https://doi.org/10.1093/bioinformatics/btp187
Sarkar I.N., Egan M.G., Coruzzi G. et al. Automated simultaneous analysis phylogenetics (ASAP): An enabling tool for phlyogenomics // BMC Bioinformatics. 2008. V. 9. № 103. P. 1–5. https://doi.org/10.1186/1471-2105-9-103
Puillandre N., Lambert A., Brouillet S., Achaz G. ABGD, Automatic Barcode Gap discovery for primary species delimitation // Mol. Ecol. 2012. V. 21. № 8. P. 1864–1877. https://doi.org/10.1111/j.1365-294X.2011.05239.x
Прохоров А.М. (гл. ред.). Себелан // Большая Советская Энциклопедия (в 30 томах). Т. 23. Сафлор – Соан. Изд. 3-е. М.: БСЭ, 1976. С. 101.
Tuniyev B.S., Orlov N.L., Ananjeva N.B., Aghasyan A.L. Snakes of the Caucasus: Taxonomic diversity, distribution, conservation. St. Petersburg; M.: KMK Scientific Press, 2019. 276 p.
Anderson S.C. The Lizards of Iran // Society for the Study of Amphibians and Reptiles. Oxford: Ohio, 1999. V. 15. 442 p.
Kidov A.A., Litvinchuk S.N. Distribution and conservation status of the Hyrcanian wood frog (Rana pseudodalmatina) in Azerbaijan // Rus. J. Herpetol. 2021. V. 28. № 2. P. 97–107. https://doi.org/10.30906/1026-2296-2021-28-2-97-107
Cyrén O. Klima und Eidechsenverbreitung. Eine Studie der geographischen Variation und Entwicklung einiger Lacerten, in sbesondereunter Berücksichtigung der klimatischen Faktoren // Meddel and enfran Göteborgs Musei Zoologiska Avdelning. 1924. V. 27. № 3. P. 1–82 + 13 p. plates.
Даревский И.С. Фауна пресмыкающихся Армении и ее зоогеографический анализ. Автореферат дис. … канд. биол. наук. Ереван, 1957. 28 с.
Мусхелишвили Т.А. О распространении полосатой ящерицы (Lacerta strigata Eichwald) и средней ящерицы (Lacerta trilineata media Lantr et Cyren) в Восточной Грузии // Сообщ. Акад. наук Груз. ССР. 1966. Т. XLIII. № 3. С. 753–756.
Kehl M. Quaternary climate change in Iran – the state of knowledge // Erdkunde. 2009. V. 63. № 1. P. 1–17. https://doi.org/10.3112/ERDKUNDE.2009.01.01
Atalay I., Efe R., Soykan A. Mediterranean Ecosystems of Turkey: Ecology of Taurus Mountains // Environment and Culture in the Mediterranean Region / Eds Efe, Cravins, Öztürk, Atalay. Newcastle, UK: Cambr. Scholars Publ., 2008. P. 3–37. https://doi.org/10.5848/csp.1087.00001
Schmidtler J.F. Orientalische Smaragdei dechsen: 2. Über Systematik und Synökologie von Lacerta trilineata, L. media und L. pamphylica // Salamandra. 1986. V. 22. № 2/3. P. 126–146.
Tuniyev B.S. On the Mediterranean influence on the formation of herpetofauna of the Caucasian isthmus and its main xerophylous refugia // Rus. J. Herpetology. 1995. V. 2. № 2. P. 95–119. https://doi.org/10.30906/1026-2296-1995-2-2-95-119
Доронин И.В., Мазанаева Л.Ф., Доронина М.А. Использование ГИС-моделирования для анализа распространения средней ящерицы, Lacerta media Lantz et Cyrén, 1920, на территории Дагестана (Россия) // Тр. Зоол. ин-та РАН. 2018. Т. 322. № 4. С. 463–480. https://doi.org/10.31610/trudyzin/2018.322.4.463
Кидов А.А., Доронин И.В., Пыхов С.Г., Доронина М.А. О новой находке Lacerta media Lantz et Cyrén, 1920 (Reptilia, Lacertidae) в Талыше // Совр. герпетология. 2019. Т. 19. Вып. 1/2. С. 68–73. https://doi.org/10.18500/1814-6090-2019-19-1-2-68-73
Дополнительные материалы отсутствуют.