

Генетика, 2022, T. 58, № 5, стр. 562-569
Стабильность популяционной системы В-хромосом корейской мыши Apodemus peninsulae (Mammalia, Rodentia) в Южном Прибайкалье: 35 лет наблюдений
И. А. Жигарев 1, *, И. А. Крищук 2, **, З. З. Борисова 3, Ю. М. Борисов 3
1 Московский педагогический государственный университет
119991 Москва, Россия
2 Научно-практический центр Национальной академии наук Беларуси по биоресурсам
220072 Минск, Беларусь
3 Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
119071 Москва, Россия
* E-mail: i.zhigarev@gmail.com
** E-mail: ikryshchuk@yandex.ru
Поступила в редакцию 02.11.2021
После доработки 26.12.2021
Принята к публикации 30.12.2021
- EDN: DOAYVG
- DOI: 10.31857/S0016675822050125
Аннотация
В течение 35 лет, с 1984 по 2019 гг., в Южном Прибайкалье в пяти районах проведены наблюдения за изменениями системы добавочных хромосом восточноазиатских (корейских) мышей (Apodemus peninsulae). Количество и структура морфотипов дополнительных хромосом статистически оставались постоянными. Анализ систем В-хромосом демонстрирует стабильность и низкую вариабельность статистических показателей за 35-летний отрезок времени, а также отсутствие статистических различий между выборками разных лет.
Многие исследования в области генетики выявили и продолжают выявлять стабильность видовых кариотипов большинства видов животных и растений. Параллельно этим наблюдениям было обнаружено, что микроэволюционные процессы в популяциях некоторых видов формируют определенные и не всегда стабильные полиморфные видовые кариотипы [1]. Наиболее загадочным явлением считается полиморфизм по так называемым дополнительным хромосомам, или В-хромосомам (Bs), присутствующим, помимо основного набора хромосом, в кариотипе многих видов. Например, подобный полиморфизм выявлен у 85 из 4380 кариологически изученных видов млекопитающих (1.9%), среди них чаще В-хромосомы встречаются у видов отряда грызуны (Rodentia) [2]. Еще в 1974 г. B.Н. Орловым [1] была опубликована сводка по полиморфизму В-хромосом для восьми видов млекопитающих. Позже М. Вуйошевич с соавт. [2] обобщили данные о В-хромосомах уже для 85 видов млекопитающих. Дополнительные сведения о млекопитающих с В-хромосомами приведены в работах [3, 4 и др.]. Современные исследования указывают на то, что добавочные хромосомы функционально активны, по крайней мере многие из них содержат гены, кодирующие белки [2, 5–9]. Следовательно представления о том, что В-хромосомы инертны можно считать устаревшими. Однако понимание роли добавочных хромосом в жизнедеятельности клеток и тем более в формировании адаптивных, в том числе микроэволюционных, процессов еще далеко от определенности. Добавочные хромосомы могут быть не у всех особей тех видов, которых относят к видам с В-хромосомами. На этом фоне известны виды, у которых практически все особи имеют В-хромосомы, в частности это восточноазиатские (корейские) мыши (Ароdemus peninsulae, Thomas, 1906) [3, 4, 10–13].
У восточноазиатских мышей выявлен значительный полиморфизм по числу и морфотипам В-хромосом в пределах всего обширного видового ареала [4, 10, 11, 13, 14]. В кариотипе клеток этих грызунов всегда присутствует основной А‑набор хромосом, состоящий из 48 акроцентрических хромосом. Дополнительные В-хромосомы обнаружены в клетках практически всех мышей, исследованных континентальных и островных популяций (на данный момент известны лишь популяции о. Сахалин и о. Стенина, представители которой не имеют Bs) [13]. Их количество может быть разным и доходить до 30, таким образом у этого вида встречаются особи, имеющие в клетках от 48 до 78 хромосом, при этом морфотипы В‑хромосом также могут существенно отличаться [4, 13–15]. По размеру и морфологии В-хромосомы группируются в пять классов: крупные метацентрические, средние метацентрические, мелкие метацентрические, мелкие акроцентрические и точечные, или микро-В-хромосомы. Почти у каждой особи из сибирских популяций восточноазиатских мышей эти классы В-хромосом образуют индивидуальные комбинации, различающиеся по их числу и соотношению морфотипов. На сегодняшний день не ясны механизмы наследования добавочных хромосом. Учитывая тот факт, что большинство комбинаций морфотипов представлены уникальными наборами (64.7 ± 1.3%), по крайней мере в выборке исследованных животных (598 особей), можно предположить случайный характер наследования В-хромосом. Однако нужно отметить и другое: в пределах всего ареала восточноазиатской мыши более 35% вариантов – это повторяющиеся комбинации, и они чаще всего приходятся на комбинации относительно некрупных хромосом. Кроме этого установлено, что в популяциях доля животных с большим количеством B‑хромосом постепенно уменьшается, пропорционально этому количеству. Одна из гипотез объясняющая этот феномен связана с усложнением процесса наследования большого количества крупных добавочных хромосом в клетках [4].
Можно предположить, что существенная изменчивость добавочных хромосом с их нерегулярным наследованием, предположительным отсутствием гомологичности и аккумуляцией в мейотических клетках [16, 17], вероятно приводят к изменчивости и стохастичности их числа и морфотипов в популяции мышей в различные годы. Для проверки этой гипотезы необходимы ряды многолетних наблюдений. На сегодняшний день в научной литературе не так много публикаций, оценивающих не только анализ временных рядов, но и простую регистрацию динамики систем В-хромосом в конкретных популяциях. Существует лишь незначительное количество подобных исследований [11, 13, 17–21]. Эти работы демонстрируют разноплановость тенденций: как вариабельность, так и постоянство систем В-хромосом в конкретных популяциях. Это в свою очередь требует дополнительных материалов и, желательно, полученных за значительные промежутки времени.
Цель настоящего исследования – определить степень изменчивости популяционной структуры вариантов системы добавочных хромосом восточноазиатских (корейских) мышей (Apodemus peninsulae) в Южном Прибайкалье за 35-летний период наблюдений (1984–2019 гг.).
МАТЕРИАЛЫ И МЕТОДЫ
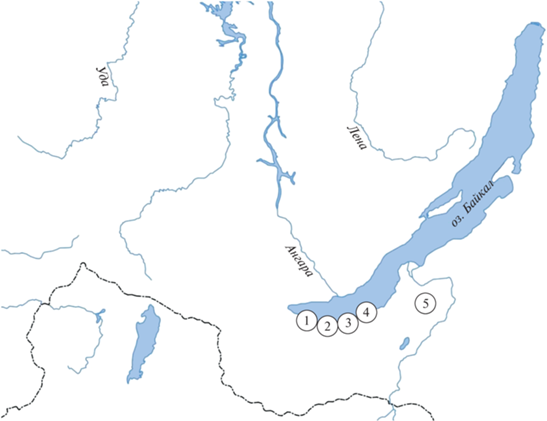
Отловы восточноазиатских мышей в Южном Прибайкалье осуществляли в разные годы. С 1984 по 2019 гг. – в пяти районах: пункт № 1 – окрестности г. Байкальск, Иркутской обл. (N 51°30′33.7′′ E 104°09′40.4′′); пункт № 2 – окрестности пос. Выдрино, Кабанский р-н, Республика Бурятия (N 51°26′6.7′′ E 104°38′13.0′′); пункт № 3 – окрестности пос. Танхой, Кабанский р-н, Республика Бурятия (N 51°29′18.5′′ E 104°50′57.0′′); пункт № 4 – окрестности г. Бабушкин, Кабанский р-н, Республика Бурятия (N 51°38′35.3′′ E 105°31′20.4′′); пункт № 5 – окрестности пос. Иволгинск, Республика Бурятия (N 51°44′45.3′′ E 107°16′31.9′′) (рис. 1).
Рис. 1.
Пункты отлова Apodemus peninsulae в Южном Прибайкалье. 1 – окрестности г. Байкальск, Иркутской обл.; 2 – окрестности пос. Выдрино, Кабанский р-н, Республика Бурятия; 3 – окрестности пос. Танхой, Кабанский р-н, Республика Бурятия; 4 – окрестности г. Бабушкин, Кабанский р-н, Республика Бурятия; 5 – окрестности поселка Иволгинск, Республика Бурятия.
В пределах 35-летнего периода мы выделили три временных отрезка: 1984–1989 гг. (материалы пунктов № 1, 4, 5), 2002–2003 гг. (№ 2, 3) и 2019 г. (№ 1, 2, 3, 4).
Расстояния между пунктами отловов составляют 20–70 км, между крайними точками – 200 км. Они располагаются на территории, где отсутствуют природные или техногенные преграды для перемещения мышей, что позволяет считать этот район географически единым.
Исследования характеристик популяционных систем добавочных хромосом в этой сейсмически активной зоне Южного Прибайкалья и в зоне аэропромвыбросов целлюлозно-бумажного комбината г. Байкальска (функционировал до 2013 г.), проведенные на начальном этапе, в 1984–2003 гг., отражены в наших предыдущих работах, там же приведены кариотипы мышей [22–24].
В 2019 г. дополнительно изучены кариотипы 42 особей Apodemus peninsulae, отловленных в окрестностях г. Байкальск, пос. Выдрино, пос. Танхой, и пос. Бабушкин (табл. 1).
Таблица 1.
Формулы изменчивости вариантов системы B-хромосом (Bs) восточноазиатских (корейских) мышей Южного Прибайкалья (2019 г.)
| № п/п | Пол | Количество хромосом | Формула вариантов Вs | Пункт отлова | |
|---|---|---|---|---|---|
| (2n = 48) + Bs | Вs | ||||
| 1 | ♀ | 53 | 5 | 0.1.0.0.04 | Окр. г. Байкальска |
| 2 | ♀ | 56 | 8 | 0.1.1.0.06 | То же |
| 3 | ♂ | 55 | 7 | 0.1.2.0.04 | » |
| 4 | ♂ | 53 | 5 | 0.2.0.0.03 | » |
| 5 | ♂ | 55 | 7 | 0.1.1.0.05 | » |
| 6 | ♀ | 56 | 8 | 0.1.0.0.07 | » |
| 7 | ♀ | 59 | 11 | 0.1.1.0.09 | » |
| 8 | ♀ | 59 | 11 | 0.1.0.0.10 | » |
| 9 | ♂ | 60 | 12 | 0.1.0.0.11 | » |
| 10 | ♀ | 60 | 12 | 0.0.0.0.12 | » |
| 11 | ♀ | 60 | 12 | 0.2.1.0.09 | » |
| 12 | ♀ | 61 | 13 | 0.1.1.0.11 | » |
| 13 | ♀ | 55 | 7 | 0.1.1.0.05 | » |
| 14 | ♂ | 57 | 9 | 0.0.2.0.07 | » |
| 15 | ♂ | 58 | 10 | 0.1.1.0.08 | » |
| 16 | ♀ | 58 | 10 | 0.1.0.0.09 | » |
| 17 | ♂ | 58 | 10 | 0.2.0.0.08 | » |
| 18 | ♀ | 59 | 11 | 0.0.3.0.08 | » |
| 19 | ♂ | 59 | 11 | 0.0.3.0.08 | » |
| 20 | ♀ | 59 | 11 | 0.1.1.0.09 | » |
| 21 | ♀ | 54 | 6 | 0.2.3.0.01 | » |
| 22 | ♀ | 60 | 12 | 0.2.0.0.10 | » |
| 23 | ♂ | 60 | 12 | 0.2.0.0.10 | » |
| 24 | ♀ | 52 | 4 | 0.1.1.0.02 | Окр. пос. Танхой |
| 25 | ♂ | 52 | 4 | 0.3.0.0.01 | То же |
| 26 | ♀ | 59 | 11 | 0.2.1.0.08 | Окр. пос. Выдрино |
| 27 | ♂ | 56 | 8 | 0.2.0.0.06 | То же |
| 28 | ♀ | 54 | 6 | 0.0.1.0.05 | Окр. пос. Танхой |
| 29 | ♂ | 57 | 9 | 0.0.1.0.08 | То же |
| 30 | ♂ | 58 | 10 | 0.0.1.0.09 | » |
| 31 | ♀ | 56 | 8 | 1.0.0.1.06 | » |
| 32 | ♀ | 54 | 6 | 0.1.0.0.05 | » |
| 33 | ♂ | 59 | 11 | 0.1.1.0.09 | » |
| 34 | ♀ | 54 | 6 | 0.1.1.0.04 | Окр. пос. Бабушкин |
| 35 | ♀ | 55 | 7 | 0.2.0.0.05 | То же |
| 36 | ♂ | 55 | 7 | 0.1.1.0.05 | » |
| 37 | ♀ | 57 | 9 | 0.2.1.0.06 | » |
| 38 | ♂ | 59 | 11 | 0.1.0.0.10 | » |
| 39 | ♂ | 59 | 11 | 0.0.1.0.10 | » |
| 40 | ♂ | 59 | 11 | 0.1.0.0.10 | » |
| 41 | ♀ | 60 | 12 | 0.1.1.0.10 | » |
| 42 | ♂ | 61 | 13 | 0.1.1.0.11 | » |
Хромосомные препараты готовили прямым методом из клеток костного мозга с предварительным введением внутрибрюшинно 0.5 мл 0.04% раствора колхицина [1]. Для характеристики кариотипа каждого животного мы использовали анализ не менее чем 20 метафазных пластинок.
В настоящей работе нами использована система классификации В-хромосом (Bs) и формула их цифрового кодирования по Ю.М. Борисову [22, 23]. В табл. 1 отображены формулы кодирования добавочных хромосом мышей, отловленных в 2019 г. в районе исследования. В формуле первая цифра обозначает количество крупных двуплечих хромосом, равных по размерам 1–8 парам А-хромосом (I класс). Вторая цифра обозначает количество хромосом II класса (двуплечих, средних размеров, равных 17–23 парам А-хромосом); третья – количество хромосом III класса (также метацентрических, мелких, равных по размерам 17–23 парам А-хромосом); четвертая ‒ количество хромосом IV класса (акроцентрических, мелких, равных 17–23 парам А-хромосом); пятая цифра ‒ количество микро-В-хромосом V класса. Последние настолько мелкие, что определить положение центромеры не представляется возможным. На рис. 2 показаны метафазные пластинки Apodemus peninsulae из трех пунктов отлова: Байкальск (№ 13, табл. 1); Выдрино (№ 27, табл. 1); Бабушкин (№ 34, табл. 1).

РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Дополнительные хромосомы обнаружены у всех представителей восточноазиатских мышей, обитающих в Южном Прибайкалье. Их число в клетках мышей варьирует от 4 до 13, в то время как в границах всего ареала наблюдаются предельные вариации от 0 (о. Сахалин) до 30 дополнительных хромосом (Средняя Сибирь) в клетках конкретного зверька [4]. Таким образом, исследуемую популяцию можно отнести к популяциям, представители которых имеют умеренное содержание добавочных хромосом. Это же подтверждается показателем “индекс условной массы В-хромосом” (см. ниже). Напомним, что избыточное содержание В-хромосом (предположительно) усложняет их наследование.
До сих пор нет ясности, от каких факторов зависит то или иное количество дополнительных хромосом в клетках животных, при том, что основной набор из 48 хромосом у этого вида остается неизменным. В многочисленных работах показано разнообразие сочетаний В-хромосом пяти классов [4, 12, 16, 17, 26]. Таким образом, вариабельность сочетаний пяти классов добавочных хромосом носит индивидуальный характер, однако определенные закономерности распределения В-хромосом в пределах географических популяциях восточноазиатских мышей все-таки выявлены [4, 13, 19, 22, 23].
За 35 лет наблюдений в Южном Прибайкалье фиксируется удивительная стабильность основных показателей системы B-хромосом у корейских мышей. Среднее число В-хромосом на всем протяжении наблюдений оставалось статистически постоянной величиной, находясь в пределах от 8.11 ± 0.9 до 9.14 ± 0.4, без достоверных отличий, р ≥ 0.28 (рис. 3, табл. 2). Дисперсионный анализ (ANOVA) также не подтверждает различие дисперсий (F(2; 81) = 0.55; p = 0.57).
Рис. 3.
Динамика статистических показателей среднего количества дополнительных хромосом (Bs) в популяциях Apodemus peninsulae Южного Прибайкалья (1984–2019 гг.).

Таблица 2.
Статистические показатели изменчивости вариантов системы B-хромосом у восточноазиатских мышей Южного Прибайкалья
| Количество хромосом | Годы | ||
|---|---|---|---|
| 1984‒1989 | 2002‒2003 | 2019 | |
| Bs | $\frac{{9.00 \pm 0.49}}{{4 - 13,\,\,n = 33}}$ | $\frac{{8.11 \pm 0.90}}{{4 - 11,\,\,n = 9}}$ | $\frac{{9.14 \pm 0.40}}{{4 - 13,\,\,n = 42}}$ |
| Крупные метахромосомы | 0.0 | $\frac{{0.11 \pm 0.11}}{{0 - 1,\,\,n = 9}}$ | $\frac{{0.02 \pm 0.02}}{{0 - 1,\,\,n = 42}}$ |
| Средние метахромосомы | $\frac{{1.09 \pm 0.14}}{{0 - 2,\,\,n = 33}}$ | $\frac{{1.00 \pm 0.41}}{{0 - 3,\,\,n = 9}}$ | $\frac{{1.07 \pm 0.12}}{{0 - 3,\,\,n = 42}}$ |
| Мелкие метахромосомы | $\frac{{0.82 \pm 0.17}}{{0 - 3,\,\,n = 33}}$ | $\frac{{0.67 \pm 0.24}}{{0 - 2,\,\,n = 9}}$ | $\frac{{0.79 \pm 0.13}}{{0 - 3,\,\,n = 42}}$ |
| Акросомы | 0.0 | $\frac{{0.11 \pm 0.11}}{{0 - 1,\,\,n = 9}}$ | $\frac{{0.02 \pm 0.02}}{{0 - 1,\,\,n = 42}}$ |
| Точечные (микро) хромосомы | $\frac{{7.09 \pm 0.55}}{{0 - 12,\,\,n = 33}}$ | $\frac{{6.22 \pm 0.98}}{{1 - 9,\,\,\,\,n = 9}}$ | $\frac{{7.24 \pm 0.44}}{{1 - 12,\,\,n = 42}}$ |
Структура морфотипов дополнительных хромосом за 35 лет наблюдения практически не менялась. Отсутствуют достоверные отличия, как долей типов хромосом, так и общего распределения, рассчитанного методом χ2 и через индекс сходства распределений Животовского [25]. У зверьков всегда преобладают точечные минихромосомы (72–76% от всех Bs). Их реальное количество колебалось от 0 до 12, однако среднее значение было стабильным, 6–7 хромосом в клетке (табл. 2). Вторые позиции занимали средние (14‒19%) и мелкие (около 10%) метацентрические хромосомы, их среднее количество также было стабильным и не имело статистических отличий в разные годы наблюдений. А вот крупные метацентрики и акросомы встречаются в данной популяции редко и нерегулярно (0–1 хромосома на клетку), причем эта особенность никак не менялась с течением времени.
Г.В. Рослик и И.В. Картавцева ввели удобный показатель, индекс условной массы В-хромосом или индекс Bs, который в целом характеризует весь массив дополнительных хромосом в клетке [26]. Чем он выше, тем “массивнее” этот набор. В Южном Прибайкалье индекс Bs у восточноазиатских мышей в течение 35 лет наблюдений стабилен и статистически не изменяется (рис. 4). Здесь необходимо отметить, что в пределах обширного ареала вида, в разных популяциях, этот показатель достаточно изменчив и принимает значение от 3 до 23 [4, 26]. В исследуемом регионе индекс Bs изменялся от 6 до 17, с незначительными показателями коэффициента вариации (табл. 3) и отсутствием достоверных отличий выборок разных лет (критерий Краскела–Уоллиса H(2; 84) = 0.81, p = 0.67).
Рис. 4.
Динамика среднего значения “индекс условной массы” В-хромосом в популяциях Apodemus peninsulae Южного Прибайкалья (1984–2019 гг.).

Таблица 3.
Статистические показатели изменчивости индекса Bs восточноазиатских мышей Южного Прибайкалья
| Годы | Показатели | ||||||
|---|---|---|---|---|---|---|---|
| объем выборки (n) | M | Min | Max | Std. Dev. | Coef. Var., % | SE | |
| 1984–1989 | 33 | 12.0 | 6 | 17 | 2.83 | 23.57 | 0.49 |
| 2002–2003 | 9 | 11.2 | 7 | 15 | 2.73 | 24.31 | 0.91 |
| 2019 | 42 | 12.2 | 7 | 17 | 2.65 | 21.78 | 0.41 |
| Всего | 84 | 12.0 | 6 | 17 | 2.71 | 22.59 | 0.29 |
Анализ систем добавочных хромосом 84 особей A. peninsulae из Южного Прибайкалья показывает наличие умеренного (в сравнении с другими популяциями) разнообразия вариантов сочетаний В-хромосом, встречающихся у зверьков, их оказалось 46 (в пределах всего ареала на данный момент установлено 292 варианта комбинаций). При этом уникальных наборов В-хромосом (индекс уникальности Bs), встречающихся единожды в пределах всего ареала, только 4.1 ± 0.76% (для выборки из 598 мышей со всего ареала этот показатель существенно выше ‒ 64.7 ± 1.3%) [4].
Если рассмотреть долю мышей, которая обладает уникальными, не встречающимися в других местах ареала, наборами В-хромосом, то она опять же небольшая и составляет всего 14.3% от всех особей популяции Южного Прибайкалья, остальные зверьки имеют наборы, которые уже отмечены нами у других мышей либо этих, либо иных популяций. Нужно обратить внимание на то, что для всего ареала этот показатель существенно выше ‒ почти треть из всех исследованных зверьков (31%) восточноазиатских мышей имеет уникальные наборы В-хромосом. Таким образом, относительно высокая повторяемость комбинаций наборов В-хромосом у мышей на юге Байкальского побережья, может говорить о более слабом влиянии случайных факторов в формировании этих комбинаций.
Конкретные наборы В-хромосом, отмеченные для мышей Южного Прибайкалья, встречаются у особей разных популяций, однако подавляющее большинство из них – это либо внутрипопуляционные комбинации, либо встречающиеся в соседних популяциях Бурятии, Монголии, а также Центральной Сибири и западных популяций (Новосибирская и Кемеровская обл.), реже Северного Китая и Японии (о. Хоккайдо). Любопытно, что ни разу не были отмечены сходные наборы В‑хромосом у зверьков популяций противоположного берега Байкала (Западного и Северного Прибайкалья) и Приморья.
Таким образом, анализ систем В-хромосом в популяции восточноазиатской мыши Южного Прибайкалье показывает стабильность и низкую вариабельность статистических показателей за 35-летний отрезок времени, а также отсутствие статистических различий между выборками разных лет.
Авторы выражают благодарность М.Ю. Борисову, М.В. Борисовой, В.А. Хотилович и И.В. Онищук за оказанную помощь в отлове грызунов.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Орлов В.Н. Кариосистематика млекопитающих. (Цитогенетические методы в систематике млекопитающих). М.: Наука, 1974. 208 с.
Vujosevic M., Rajicic M., Blagojevic J. B chromosomes in populations of mammals revisited // Genes. 2018. V. 9(10). P. 110–136. https://doi.org/10.3390/genes9100487
Борисов Ю.М., Мышлявкина Т.А. В-хромосомы // Усп. совр. биологии. 2018. Т. 138. С. 336–351.
Borisov Yu.M., Zhigarev I.A. B chromosome system in the Korean field mouse Apodemus peninsulae Thomas 1907 (Rodentia, Muridae) // Genes. 2018. V. 9(10). P. 147–158. https://doi.org/10.3390/genes9100472
Trifonov V.A., Perelman P.L., Kawada S.I. Complex structure of B-chromosomes in two mammalian species: Apodemus peninsulae (Rodentia) and Nyctereutes procyonoides (Carnivora) // Chromosome Res. 2002. V. 10. P. 109–116. https://doi.org/10.1023/A:1014940800901
Rubtsov N.B., Karamysheva T.V., Andreenkova O.V. et al. Comparative analysis of micro and macro B chromosomes of Korean field mouse Apodemus peninsulae (Rodentia, Murinae) performed by chromosome microdissection and FISH // Cytogenet. Genome Res. 2004. P. 289–294. https://doi.org/10.1159/000056786
Rubtsov N.B., Borisov Yu.M. Sequence composition and evolution of mammalian B chromosomes // Genes. 2018. V. 9(10). P. 91–109. https://doi.org/10.3390/genes9100490
Makunin A., Romanenko S., Beklemisheva V. et al. Sequencing of supernumerary chromosomes of red fox and raccoon dog confirms a non-random gene acquisition by B chromosomes // Genes. 2019. V. 9(8). P. 201–2015. https://doi.org/10.3390/genes9080405
Trifonov V.A., Dementyeva P.V., Beklemisheva V.R. et al. Supernumerary chromosomes, segmental duplications, and evolution // Russ. J. Genet. 2010. V. 46. P. 1094–1096. https://doi.org/10.1134/S1022795410090206
Hayata I. Chromosomal polymorphism caused by supernumerary chromosomes in the field mouse, Apodemus giliacus // Chromosome. 1973. V. 42. P. 403–414. https://doi.org/10.1007/BF00399408
Bekasova T.S., Vorontsov N.N., Korobitsyna K.V., Korablev V.P. B-chromosomes and comparative karyology of the mice of the genus Apodemus // Genetica. 1980. V. 52–53. P. 33–43.
Картавцева И.В. Кариосистематика лесных и полевых мышей (Rodentia, Muridae). Владивосток: Дальнаука, 2002. 142 с.
Kartavtseva I.V., Roslik G.V. A complex B chromosome system in the Korean field mouse Apodemus peninsulae // Cytogenet. Genome Res. 2004. V. 106. № 2–4. P. 271–278. https://doi.org/10.1159/000079298
Борисов Ю.М., Жигарев И.А., Шефтель Б.И. В-хромосомы восточноазиатских мышей (Apodemus peninsulae Thomas, 1907 (Rodentia, Muridae)) на восточных склонах Цинхай-Тибетского плато (КНР) // Генетика. 2020. Т. 56. № 10. С. 1184–1188. https://doi.org/10.31857/S0016675820090039
Борисов Ю.М., Шефтель Б.И., Сафронова Л.Д., Александров Д.Ю. Устойчивость популяционных систем В-хромосом восточноазиатской мыши Apodemus peninsulae Прибайкалья и Северной Монголии // Генетика. 2012. Т. 48. № 10. С. 1190–1199.
Kolomiets O.L., Borbiev T.E., Safronova L.D. et al. Synaptonemal complex analysis of B-chromosome behavior in meiotic prophase I in the East-Asiatic mouse Apodemus peninsulae (Muridae, Rodentia) // Cytogenet. Cell Genet. 1988. V. 48. № 3. P. 183–187.
Борисов Ю.М. Процесс увеличения числа и вариантов системы В-хромосом мышей Apodemus peninsulae в популяции Горного Алтая за 26-летний период // Генетика. 2008. Т. 44. № 9. С. 1227–1237.
Борисов Ю.М., Бочкарев М.Н., Карамышева Т.В. и др. Феномен увеличения числа В-хромосом у восточноазиатских мышей Apodemus peninsulae (Mammalia, Rodentia) в популяции Горного Алтая // Докл. РАН. 2007. Т. 412. № 1. С. 126–128.
Borisov Y.M., Abramov S.A., Borisova M.Y., Zhigarev I.A. The occurrence of dot-like micro B chromosomes in Korean field mice Apodemus peninsulae from the shore of the Teletskoye Lake (Altai Mountains) // Comparative Cytogenetics. 2020. V. 14(1). P. 97–105. https://doi.org/10.3897/CompCytogen.v14i1.47659
Борисов Ю.М. Нестабильность В-хромосом соматических и герминативных клеток мышей Apodemus peninsulae // Генетика. 2008. Т. 44. № 8. С. 1101–1107.
Гилева Э.А. Система В-хромосом у копытных леммингов Dicrostonyx torquatus Pall., 1779 из природных и лабораторных популяций // Генетика. 2004. Т. 40. № 12. С. 1686–1694.
Борисов Ю.М. Система В-хромосом восточноазиатской мыши – маркер популяций в Прибайкалье // Генетика. 1990. Т. 26. № 12. С. 2215–2224.
Борисов Ю.М. Цитогенетическая дифференциация популяций восточноазиатской мыши в Восточной Сибири // Генетика. 1990. Т. 26. № 10. С. 1828–1839.
Борисов Ю.М., Шефтель Б.И., Сафронова Л.Д., Александров Д.Ю. Устойчивость популяционных систем В-хромосом восточноазиатской мыши Apodemus peninsulae в Прибайкалье и Северной Монголии // Генетика. 2012. Т. 48. №. 10. С. 1189–1198.
Животовский Л.А. Показатель сходства популяций по полиморфным признакам // Журн. общей биологии. 1979. Т. 40. № 4. С. 587–602.
Рослик Г.В., Картавцева И.В. Морфотипы В-хромосом Ароdemus peninsulae (Rodentia) Дальнего Востока России // Цитология. 2012. Т. 54. № 1. С. 66–77.
Дополнительные материалы отсутствуют.