

Генетика, 2022, T. 58, № 6, стр. 678-684
Географическая изменчивость инверсионного полиморфизма малярийного комара Anopheles messeae Fall. (Diptera: Culicidae) в Томской области
А. К. Сибатаев 1, И. Э. Вассерлауф 1, К. Е. Усов 1, С. С. Алексеева 1, Г. М. Абылкасымова 2, В. Н. Стегний 1, *
1 Томский государственный университет
634050 Томск, Россия
2 Институт генетики и физиологии
050060 Алматы, Казахстан
* E-mail: stegniy@res.tsu.ru
Поступила в редакцию 25.11.2021
После доработки 11.01.2022
Принята к публикации 03.02.2022
- EDN: BQABWO
- DOI: 10.31857/S0016675822060091
Аннотация
Исследовали хромосомную изменчивость комаров Anopheles messeae Fall. в шести населенных пунктах Томской обл. (Западная Сибирь). Определены частоты хромосомных инверсий в локальных выборках An. messeae. Установлено преобладание инверсионных вариантов аутосом: 2R0, 3R0, 3L0 в гомозиготном состоянии. В исследованных нами выборках по половой хромосоме XL отмечен избыток гетерозигот по инверсии XL01. По-видимому, направление отбора в сторону хромосомных вариантов XL01, 2R00, 3R00 и 3L00 в выборках Томской области An. messeae можно объяснить климатическими изменениями последних десятилетий, создающих оптимальные условия для роста и развития комаров.
Важным механизмом видообразования в отряде Diptera являются хромосомные перестройки, инверсии. Именно поэтому виды отличаются по фиксированным инверсиям [1, 2]. В то же время большинство видов, адаптируясь к определенным климатическим условиям, имеют флуктуирующие инверсии, приводящие к инверсионному полиморфизму хромосом [3–7]. Роль хромосомных инверсий в адаптации и эволюции различных видов двукрылых изучается в течение нескольких десятилетий [8–10]. Многолетний мониторинг сезонной цикличности и клинального распределения частот инверсий по ареалам полиморфных видов двукрылых дают полную картину динамики инверсионного полиморфизма в популяциях определенных групп Diptera в ответ на климатические изменения в природе [10–13]. За последнее время произошли климатические изменения во многих областях земного шара. В связи с этим расширились ареалы многих видов насекомых, особенно эпидемически опасных, среди которых переносчики малярии – комары рода Anopheles [14, 15]. Согласно последним оценкам экспертов, потенциальный ареал малярии в XXI в. при потеплении климата расширяется в основном к северу. На территориях, где малярия человека эндемична, увеличивается длительность сезона передачи. Значительно возрастает число людей, проживающих в зонах с большим риском заражения малярией. В последние 15–20 лет существенно изменилась структура завоза малярии в Россию. Изменения климата, произошедшие в ХХ в. на территории стран СНГ и Балтии, сказались на ареалах распространения переносчиков и условиях развития возбудителей в организме переносчиков. Северные границы ареалов малярийных комаров имеют тенденцию к продвижению на север, предполагают возможность замещения северных популяций южными. Для отдельных территорий страны влияние потепления климата может сказаться на частоте заболеваний малярией [16–18]. В связи с этим мониторинг популяций малярийных комаров является актуальным в настоящее время. Малярийные комары рода Anopheles представляют собой удобные модельные объекты для популяционно-генетического анализа. В клетках некоторых тканей комаров содержатся политенные хромосомы, на которых хорошо идентифицируются фиксированные и флуктуирующие перестройки [4, 10–12].
В Западной Сибири обитает два вида малярийных комаров из комплекса “maculipennis” – Anopheles messeae Fall. и An. beklemishevi Stegniy and Kabanova, для которых установлен внутривидовой полиморфизм по хромосомным инверсиям [10–13]. An. messeae является самым распространенным видом в Голарктическом комплексе видов “maculipennis” и населяет всю бореальную область Палеоарктики. Этот вид детально изучается в плане анализа инверсионного полиморфизма на всем протяжении своего огромного ареала в течение 50 лет [4, 5, 10–12, 19]. Выявлены пространственно-временные закономерности распределения инверсионного полиморфизма и его адаптивное значение. Помимо географического анализа генетической структуры An. messeae изучена и локальная изменчивость инверсионного полиморфизма в системах смежных или близких популяций [10, 13, 19]. Обнаружение межпопуляционных отличий по инверсионным генотипам An. messeae в локальных областях ставит вопрос о механизмах генотипической дифференциации. Продолжение наблюдений за географической динамикой инверсионного полиморфизма малярийного комара An. messeae интересно с точки зрения оценки его адаптивного характера в связи с потеплением климата.
Настоящая работа содержит описание географической изменчивости инверсионного полиморфизма малярийного комара Anopheles messeae Fall. в Томской обл. (Западная Сибирь).
МАТЕРИАЛЫ И МЕТОДЫ
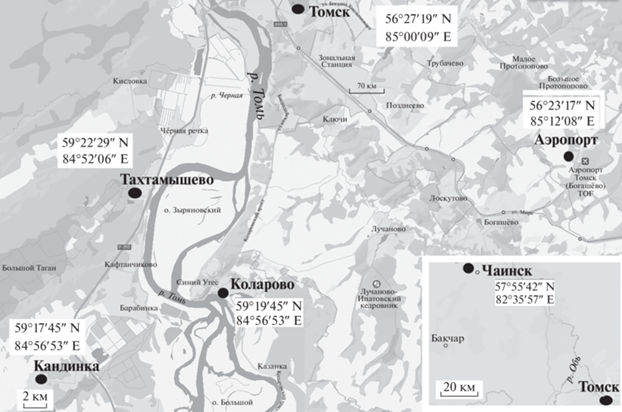
В 2020 г. было собрано 470 личинок малярийных комаров в шести населенных пунктах Томской обл. (Россия): Ботанический сад г. Томск (17.05.2020), п. Коларово (29.05.2020; 21.06.2020; 29.07.2020; 8.08.2020), п. Тахтамышево (15.05.2020), п. Кандинка (21.05.2020; 20.06.2020), п. Аэропорт (12.06.2020), п. Чаинск (26.05.2020; 15.06.2020). Расположение мест выплода малярийных комаров показано на карте (рис. 1). Отлавливали личинок четвертого возраста и фиксировали этанол-уксусной смесью Карнуа в соотношении 3 : 1.
Из личинок выделяли слюнные железы, содержащие политенные хромосомы. Хромосомные препараты готовили стандартным лактоацетоорсеиновым методом [11]. Политенные хромосомы визуализировали с помощью светового микроскопа Zeiss Primo Star и получали микрофотографии политенных хромосом с помощью микроскопа Zeiss Imager A1. Анализ хромосомных инверсий проводили с помощью цитогенетической карты политенных хромосом слюнных желез An. messeae [12, 20]. Частоты хромосомных инверсий подсчитывали в процентном соотношении с помощью формулы закона Харди–Вайнберга, сравнивали частоты инверсий в выборках методом χ2.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Со второй половины 80-х до начала 90-х гг. XX в. было отмечено направленное изменение концентрации хромосомных вариантов на всем изученном ареале в пользу тех, которые доминировали на юге и юго-западе ареала An. messeae [13, 15, 21]. Известно, что для комаров одним из основных абиотических лимитирующих факторов на всех стадиях развития является температурный режим. Это экспериментально показано в 1981 г. на личиночных популяциях п. Коларово, где была определена адаптивная ценность гетерозигот XL01 и 3R01 в результате повышения их частот при экстремально низких и высоких температурах развития личинок. При оптимальных же температурах отбор шел в пользу гомозиготного варианта 3R00 [12]. Поэтому направление отбора в сторону хромосом XL01, 2R00, 3R00 и 3L00 в Коларовской популяции можно объяснить климатическими изменениями последних десятилетий, создающими оптимальные условия для роста и развития комаров.
Проведенный в настоящей работе цитогенетический анализ личинок комаров An. messeae выявил инверсионный полиморфизм по половой хромосоме XL и аутосомам (обозначения инверсий согласно карте хромосом An. messeae) [4].
Во всех изученных выборках Томской обл. в кариотипах преобладали хромосомные варианты 2R0, 3R0, 3L0 аутосом в гомозиготном состоянии (табл. 1). В исследованных нами выборках инверсионный полиморфизм проявлялся по половой хромосоме XL. Были выявлены следующие хромосомные варианты: XL00, XL11, XL01, XL12. Также хотелось бы отметить, что в выборке п. Коларово была обнаружена у одной особи новая инверсия XL15 в гетерозиготном состоянии, захватывающая район 2BC–3BC (рис. 2).
Таблица 1.
Инверсионный полиморфизм в изученных выборках An. messeae Томской обл.
| Вариант хромосом | Частоты хромосомных вариантов в выборках (f ± sf), % | |||||
|---|---|---|---|---|---|---|
| Коларово | Тахтамышево | Томск | Кандинка | Чаинск | Аэропорт | |
| Cамцы (гоносомы) | ||||||
| XL0 | 22.7 ± 4.8 | 50.0 ± 17.7 | 35.7 ± 12.8 | 21.4 ± 6.3 | 30.4 ± 9.6 | 6.9 ± 4.6 |
| XL1 | 77.3 ± 4.8 | 50.0 ± 17.7 | 64.3 ± 12.8 | 78.6 ± 6.3 | 69.6 ± 9.6 | 93.1 ± 4.6 |
| n | 75 | 8 | 14 | 28 | 23 | 29 |
| Самки (гоносомы) | ||||||
| XL00 | 11.9 ± 2.4 (12) | 15.8 ± 8.4 (16) | 16.7 ± 8.8 (17)* | 18.8 ± 4.7 (19)* | 6.7 ± 3.2 (7) | 14.8 ± 4.8 (14) |
| XL01 | 58.9 ± 3.6 (46) | 42.1 ± 11.3 (48) | 27.7 ± 10.6 (48)* | 26.1 ± 5.3 (49)* | 45.0 ± 6.4 (38) | 51.9 ± 6.8 (48) |
| XL11 | 27.6 ± 3.3 (42) | 42.1 ± 11.3 (36) | 55.6 ± 11.7 (35)* | 55.1 ± 6.0 (32)* | 46.6 ± 6.4 (55) | 33.3 ± 6.4 (38) |
| XL12 | 1.1 ± 0.8 | 0 | 0 | 0 | 1.7 ± 1.7 | 0 |
| XL15 | 0.5 ± 0.5 | 0 | 0 | 0 | 0 | 0 |
| n | 185 | 19 | 18 | 69 | 60 | 54 |
| Самцы и самки (аутосомы) | ||||||
| 2R00 | 99.2 ± 0.5 (98) | 100.0 | 100.0 | 97.9 ± 1.4 (98) | 95.2 ± 2.4 (94) | 98.8 ± 1.2 (98) |
| 2R01 | 0.8 ± 0.5 (2) | 0 | 0 | 2.1 ± 1.4 (2) | 3.6 ± 2.0 (6) | 1.2 ± 1.2 (2) |
| 2R11 | 0 (0.01) | 0 | 0 | 0 (0.01) | 1.2 ± 1.2 (0) | 0 (0.01) |
| 3R00 | 75.8 ± 2.7 (74) | 74.1 ± 8.4 (74)* | 62.5 ± 8.6 (61)* | 76.3 ± 4.3 (76) | 66.2 ± 5.2 (66)* | 80.7 ± 4.3 (81)* |
| 3R01 | 18.1 ± 2.4 (24) | 18.5 ± 7.5 (24)* | 15.6 ± 6.4 (34)* | 19.6 ± 4.0 (23) | 20.5 ± 4.4 (31)* | 8.4 ± 3.1 (18)* |
| 3R11 | 6.2 ± 1.5 (2) | 7.4 ± 5.0 (2)* | 21.9 ± 7.3 (5)* | 4.1 ± 2.0 (1) | 13.3 ± 3.7 (3)* | 10.8 ± 3.4 (1)* |
| 3L00 | 95.0 ± 1.4 (94) | 92.6 ± 5.0 (92) | 78.1 ± 7.3 (77)* | 91.8 ± 2.8 (92)* | 85.5 ± 3.9 (84) | 96.4 ± 2.0 (96) |
| 3L01 | 5.0 ± 1.4 (6) | 3.7 ± 3.6 (7) | 12.5 ± 5.8 (21)* | 6.1 ± 2.4 (7)* | 12.1 ± 3.6 (15) | 3.6 ± 2.0 (4) |
| 3L11 | 0 (0.09) | 3.7 ± 3.6 (1) | 9.4 ± 5.2 (2)* | 2.1 ± 1.4 (2)* | 2.4 ± 1.7 (1) | 0 0 (0.04) |
| n | 260 | 27 | 32 | 97 | 83 | 83 |
Рис. 2.
Гетерозигота по инверсии XL15 половой хромосомы An. messeae. Стрелками отмечен район инверсии 2BC–3BC согласно карте [12].

По аутосомам An. messeae во всех шести изученных выборках Томской обл. частотно преобладали “юго-западные” варианты 2R00, 3R00, 3L00. Можно отметить пониженную гетерозиготность по хромосомам 3R01 и 3L01 в выборке п. Аэропорт. В большинстве выборок (п. Коларово, п. Тахтамышево, Ботанический сад г. Томск, п. Кандинка, п. Чаинск) наблюдали низкий процент частот гомозигот XL00 за счет доминирования инверсий XL11 или гетерозигот по инверсии XL01. В выборках Ботанический сад г. Томск и п. Кандинка доминируют хромосомные варианты гомозигот XL11 над гетерозиготами XL01. В выборке п. Чаинск выявлен довольно низкий процент частот гомозигот XL00. Гетерозиготность по XL хромосоме высокая в местах сбора п. Аэропорт и особенно в п. Коларово, где XL01 показывает эффект сверхдоминирования. В то же время в выборке п. Кандинка частота хромосомного варианта XL01 значительно меньше ожидаемой за счет увеличения адаптивной ценности гомозигот XL11 (табл. 1).
Оценивая возможную адаптивность инверсионных вариантов по хромосомам XL, 2R, 3R и 3L, следует отметить, что расстояние между всеми выборками (кроме п. Чаинск) составляет около 15–20 км, что предполагает активную миграцию между ними и сходство в частотах инверсионных вариантов. Однако мы видим достаточно сильные различия между выборками, особенно по частотам генотипов хромосомы XL. Учитывая эти данные, можно сделать вывод, что миграция не является фактором, нивелирующим генетические различия как смежных, так и любых близких выборок. Различия определяются, очевидно, действием локальных экологических факторов естественного отбора. Проведенные нами ранее эксперименты по температурным режимам развития личинок комаров An. messeae подтверждают значение отбора в частотных колебаниях этих инверсионных вариантов [11]. Полное доминирование гомозигот 2R00 во всех изученных выборках, видимо, обусловлено потеплением климата, что было показано в ранних работах [12].
Существенная изменчивость частот хромосомных генотипов между исследованными выборками в регионе Томской обл. не выявляется на диаграммах частот хромосомных вариантов, но проявляются различия между некоторыми выборками комаров при сравнении отдельно по каждой хромосоме (рис. 3).
Рис. 3.
Частоты хромосомных вариантов в выборках An. messeae. Вертикальными линиями указан 95% доверительный интервал. Кол – п. Коларово, Тах – п. Тахтамышево, Аэр – п. Аэропорт, Том – Ботанический сад г. Томск, Чск – п. Чаинск, Кан – п. Кандинка.

Для выявления различий по частотам хромосомных вариантов An. messeae был проведен анализ между всеми выборками с использованием критерия χ2 для таблиц сопряженности, а также было проведено попарное сравнение между собой исследуемых выборок (табл. 2). В данном методе сравнивалось значение критерия $\chi _{{{\text{эмп}}}}^{2}$ с критическими значениями $\chi _{{{\text{кр}}}}^{2}.$ На основании применения критерия χ2 Пирсона нулевая гипотеза может быть отвергнута, при условии $\chi _{{{\text{эмп}}}}^{2}$ > $\chi _{{{\text{кр}}}}^{2}$ [22].
Таблица 2.
Значения χ2 для таблиц сопряженности распределения частот хромосомных вариантов в выборках An. messeae Томской обл. (α = 0.05), d.f. = 1
| Места сбора комаров | $\chi _{{{\text{кр}}}}^{2}$ | $\chi _{{{\text{эмп}}}}^{2}$ | |
|---|---|---|---|
| XL0, XL1 | |||
| Коларово | Кандинка | 3.84 | 4.32 |
| Коларово | Чаинск | 3.84 | 5.73 |
| 2R0, 2R1 | |||
| Коларово | Чаинск | 3.84 | 8.6 |
| 3R0, 3R1 | |||
| Коларово | Кандинка | 3.84 | 4.32 |
| Коларово | Чаинск | 3.84 | 5.73 |
| Тахтамышево | Чаинск | 0.69 | 3.84 |
| Тахтамышево | Кандинка | 0.33 | 3.84 |
| 3L0, 3L1 | |||
| Томск | Кандинка | 3.84 | 25.94 |
| Аэропорт | Томск | 3.84 | 16.53 |
| Аэропорт | Чаинск | 3.84 | 7.50 |
| Коларово | Чаинск | 3.84 | 11.72 |
Х-хромосома – XL. По XL хромосоме выявлены различия распределения частот хромосомных вариантов XL0 и XL1 между выборками п. Коларово и п. Кандинка, а также между выборками п. Коларово и п. Чаинск. Наименьшая частота хромосомного варианта XL0 отмечена в выборках п. Чаинск и п. Кандинка. По этим параметрам данные выборки отличаются от выборки п. Коларово (рис. 3, табл. 2).
Хромосома 2 – 2R-плечо. Между выборками п. Коларово и п. Чаинск по хромосоме 2R выявлены различия по распределению частот хромосомных вариантов 2R0 и 2R1 (табл. 2, рис. 3). Эти выборки находятся далеко друг от друга, на расстоянии 213 км, а также п. Чаинск находится намного севернее п. Коларово (табл. 2, рис. 3).
Хромосома 3. По 3R-хромосоме выявлены достоверные различия по распределению частот хромосомных вариантов 3R0 и 3R1 между выборками п. Коларово и п. Чаинск, п. Коларово и п. Кандинка, а также между выборками населенных пунктов п. Тахтамышево и п. Чаинск, п. Тахтамышево и п. Кандинка (табл. 2, рис. 3). По 3L-хромосоме выявлены достоверные различия по распределению частот хромосомных вариантов 3L0 и 3L1 между выборками п. Коларово и п. Чаинск, п. Коларово и п. Кандинка, а также между выборками п. Тахтамышево и п. Чаинск, п. Тахтамышево и п. Кандинка (табл. 2, рис. 3).
На примере популяции An. messeae п. Коларово изучена сезонная изменчивость инверсионной структуры в течение 2020 г. В данной популяции нами проводился мониторинг динамики инверсионной структуры популяции на протяжении более 40 лет. Для выявления изменения в течение сезона частот хромосомных вариантов An. messeae был проведен внутрипопуляционный анализ, используя критерий χ2 для таблиц сопряженности (табл. 3). Расчет критерия χ2 для таблиц сопряженности не выявил различий сезонной изменчивости инверсионной структуры политенных хромосом комаров в течение 2020 г. Выявлено доминирование гетерозиготы хромосомного варианта XL01 в течение сезона 2020 г. (табл. 3).
Таблица 3.
Изменение частот хромосомных вариантов в популяции An. messeae п. Коларово в течение сезона
| Хромосомный вариант | Частоты хромосомных вариантов, % | |||
|---|---|---|---|---|
| 05/2020 | 06/2020 | 07/2020 | 08/2020 | |
| Самцы | ||||
| XL0 | 30.0 ± 14.5 | 33.3 ± 12.6 | 7.1 ± 6.8 | 20.0 ± 10.3 |
| XL1 | 70.0 ± 14.5 | 66.7 ± 12.6 | 92.9 ± 6.8 | 80.0 ± 10.3 |
| n | 10 | 21 | 14 | 15 |
| Самки | ||||
| XL00 | 10.3 ± 4.8 | 12.3 ± 4.7 | 6.5 ± 5.9 | 15.1 ± 6.2 |
| XL01 | 56.4 ± 7.9 | 77.5 ± 5.9 | 38.7 ± 11.8 | 48.6 ± 8.7 |
| XL11 | 33.3 ± 7.5 | 8.2 ± 3.9 | 54.8 ± 12.0 | 33.3 ± 8.2 |
| XL12 | 0 | 0 | 0 | 3.0 ± 3.0 |
| XL14 | 0 | 2.0 ± 2.0 | 0 | 0 |
| n | 39 | 49 | 31 | 33 |
| Самцы и самки | ||||
| 2R00 | 100.0 | 100.0 | 100.0 | 97.9 ± 2.0 |
| 2R01 | 0 | 0 | 0 | 2.1 ± 2.0 |
| 2R11 | 0 | 0 | 0 | |
| 3R00 | 75.5 ± 6.1 | 74.3 ± 5.2 | 75.6 ± 6.4 | 77.1 ± 6.0 |
| 3R01 | 18.4 ± 5.5 | 17.1 ± 4.5 | 22.2 ± 6.2 | 16.7 ± 5.4 |
| 3R11 | 6.1 ± 3.4 | 8.6 ± 3.3 | 2.2 ± 2.2 | 6.2 ± 3.5 |
| 3L00 | 98.0 ± 2.0 | 100 | 91.1 ± 4.2 | 91.7 ± 4.0 |
| 3L01 | 2.0 ± 2.0 | 0 | 8.9 ± 4.2 | 8.3 ± 4.0 |
| 3L11 | 0 | 0 | 0 | |
| n | 49 | 70 | 45 | 48 |
Таким образом, наши исследования выявили географическую изменчивость инверсионных генотипов, которая обусловлена действием локального отбора. Также следует отметить, что структура всех изученных выборок по инверсионному варианту аутосом 2R0, 3R0, 3L0 стала сходной с европейскими популяциями, но отличается от них по изменению инверсионных частот по половой хромосоме в сторону увеличения гомозигот XL1 и гетерозигот по инверсии XL01.
Исследование выполнено в рамках государственного задания Министерства науки и высшего образования Российской Федерации (проект № FSWM-2020-0019), а также при поддержке Программы развития Томского государственного университета (Приоритет-2030).
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Стегний В.H. Эволюционное значение хромосомных инверсий // Журн. общей биологии. 1984. Т. 45. № 1. С. 3–15.
Баклушинская И.Ю. Хромосомные перестройки, реорганизация генома и видообразование // Зоолог. журн. 2016. Т. 95. № 4. С. 376–393. https://doi.org/10.7868/S0044513416040036
Чубарева Л.А. Хромосомный полиморфизм в природных популяциях кровососущих мошек и некоторых других двукрылых насекомых // Цитология. 1974. Т. 6. № 3. С. 267–280.
Стегний В.Н., Новиков Ю.М., Плешкова Г.Н., Кабанова В.М. Инверсионный полиморфизм малярийного комара Anopheles messeae Fall. Сообщение II. Межпопуляционная вариабельность инверсионных частот // Генетика. 1978. Т. 14. № 6. С. 1016–1023.
Русакова А.М., Артемов Г.Н., Стегний В.Н. Инверсионный полиморфизм природных популяций малярийного комара (Anopheles messeae Fall.) восточной части ареала // Вестник Томского гос. ун-та. Биология. 2011. № 2(14). С. 117–121.
Krimbas C., Powell J. Drosophila Inversion Polymorphism. Boca Raton: CRC Press, 1992. 576 p.
Hoffmann A.A., Sgro C.M., Weeks A.R. Chromosomal inversion polymorphisms and adaptation // Trends in Ecol. & Evol. 2004. V. 19. P. 482–488.
Dobzhansky Th. Genetics of Evolutionary Process. N.Y., London: Columbia Univ. Press, 1971. 520 p.
Кикнадзе И.И. Роль хромосомного полиморфизма в дивергенции популяций и видов в роде Chironomus (Diptera) // Зоолог. журн. 2008. Т. 87. № 6. С. 686–701.
Стегний В.Н. Популяционная генетика и эволюция малярийных комаров. Томск: Изд-во Томского ун-та, 1991. 136 с.
Кабанова В.М., Карташова Н.Н., Стегний В.Н. Кариологическое исследование природных популяций малярийного комара в Среднем Приобье I. Характеристика кариотипа Anopheles maculipennis messeae // Цитология. 1972. Т. 14. № 5. С. 613–636.
Стегний В.H., Кабанова В.М., Новиков Ю.М. Кариотипическое исследование малярийного комара // Цитология. 1976. Т. 18. № 6. С. 760–766.
Артемов Г.Н., Русакова А.М., Стегний В.Н. Инверсионный полиморфизм малярийного комара Anopheles messeae в природных популяциях // Вестник Томского гос. ун-та. Бюлл. оперативной научной информации. 2004. № 30. С. 3–7.
Tolle M.A. Mosquito-borne Diseases // Current Problems in Pediatric and Adolescent Health Care. 2009. V. 39. I. 4. P. 97–140. https://doi.org/10.1016/j.cppeds.2009.01.001
Новиков Ю.М. Эффекты глобального потепления: направленная динамика пропорции видов Anopheles и цитогенетической структуры таксона Anopheles messeae Fall. в Западной Сибири // Пробл. эвол. цитогенетики, селекции и интродукции. Матер. научных чтений, посвященных 100-летию профессора В.П. Чехова. 2–5 декабря 1997 г. Томск: 1997. С. 73–86.
Всемирная Организация Здравоохранения. Европейское региональное бюро г. Копенгаген. Региональная стратегия: от борьбы к элиминации малярии в Европейском регионе. ВОЗ 2006–2015 гг. Копенгаген: ВОЗ, 2006. 44 с.
Ревич Б.А. Изменение здоровья населения России в условиях меняющегося климата // Демография. 2008. С. 140–150.
Юрченко Ю.А., Белевич О.Э. Малярия и малярийные комары (Anopheles, Culicidae) Новосибирской области // Сибирский экол. журн. 2010. Т. 17. № 5. С. 733–737.
Стегний В.Н., Пищелко А.О., Сибатаев А.К., Абылкасымова Г. Пространственно-временные изменения частот хромосомных инверсий по ареалу малярийного комара Anopheles messeae Fall. (Culicidae) за 40-летний период мониторинга // Генетика. 2016. Т. 52. № 6. С. 664–671.
Стегний В.Н. Генетические механизмы адаптации и видообразования двукрылых насекомых (на примере малярийных комаров): Дис. … докт. биол. наук. Томск: Томский гос. ун-т, 1983. 323 с.
Гордеев М.И., Сибатаев А.К. Цитогенетическое и фенотипическое изменение в центральных и периферических популяциях малярийных комаров Anopheles messeae Fall. (Diptera, Culicidae) // Генетика. 1996. Т. 32. № 9. С. 1199–1205.
Бондарчук С.С., Бондарчук И.С. Статобработка экспериментальных данных в MS Excel. Томск: Изд-во Томского гос. пед. ун-та, 2018. 433 с.
Дополнительные материалы отсутствуют.