

Генетика, 2023, T. 59, № 4, стр. 437-447
Мониторинг генетической структуры популяции соболя (Martes zibellina Linnaeus, 1758) Среднего Приамурья
Л. В. Фрисман 1, *, А. Л. Брыкова 1
1 Институт комплексного анализа региональных проблем Дальневосточного отделения
Российской академии наук
679014 Биробиджан, Россия
* E-mail: l.frisman@mail.ru
Поступила в редакцию 09.06.2022
После доработки 25.08.2022
Принята к публикации 30.08.2022
- EDN: AVCRVZ
- DOI: 10.31857/S0016675823040033
Аннотация
На основе анализа микросателлитных локусов Ма3 и Mer041 проведен мониторинг генетической структуры популяции соболя Среднего Приамурья по материалам охотничьих сборов зимних сезонов с 2011 по 2021 г. Рассмотрены три географические субпопуляции соболя Буреинского нагорья и одна с восточных склонов хребта Сихотэ-Алинь. Показано, что группа специфических аллелей, характеризующих различия соболиного населения Буреинского нагорья и Сихотэ-Алиня, представлена четырьмя аллелями. Проведенный мониторинг двух микросателлитных локусов в субпопуляциях Буреинского нагорья показал более высокое аллельное разнообразие в северной автохтонной по сравнению с более южными, возникшими на основе расселения северной в первой половине XX в. Показана относительная однородность популяционной структуры соболя Буреинского нагорья, как правило не приводящая к статистически значимому уровню дифференциации субпопуляций ни в пространстве, ни во времени. Обнаружен различающийся характер динамики аллельного разнообразия в субпопуляциях левобережья (Буреинское нагорье) и правобережья (хребет Сихотэ-Алинь) р. Амур. Выдвинута гипотеза, что временная гетерогенность в Сихотэ-Алинской субпопуляции может указывать на существование на восточных склонах Сихотэ-Алиня пространственно неоднородной популяционной структуры, сформированной из потомков и возможных гибридов Сихотэ-Алинской автохтонной и интродуцированных форм. Рассматривается влияние антропогенного фактора на динамику генетической структуры.
Соболь (Martes zibellina Linnaeus, 1758) – промысловый пушной вид млекопитающих, заселяющий хвойные и смешанные леса северной Евразии. Его ареал протяжен от Урала до Тихого океана по территории России и частично захватывает сопредельные страны: Монголию, Китай, Корею и Японию. В первой половине XX в. высокая ценность пушнины и бесконтрольный отлов привели к практически полному исчезновению соболя на российской части ареала. Запрет промысла и последующая интродукция животных на угнетенные территории из сохранившихся популяций отдаленных участков ареала восстановили численность, но сформировавшаяся структура популяций отдельных регионов и вида в целом должна была, несомненно, отличаться от исходной, что привлекает внимание к ее описанию и изучению.
Исследование генетической изменчивости и структуры популяций соболя было начато на основе анализа маркеров митохондриальной ДНК на материале северо-восточной части ареала [1, 2] и затем расширено на пределы всего ареала [3, 4]. Для исследования генетической структуры природных популяций соболя в настоящее время активно и успешно применяется анализ микросателлитов – кодоминантно наследуемых и адаптивно нейтральных ядерных генетических маркеров. Проведено исследование соболя пространственно разобщенных территорий (Урал, Сибирь, хребет Сихотэ-Алинь и п-ов Камчатка) [5, 6]. В настоящее время все более пристальное внимание уделяется генетическому исследованию соболиного населения отдельных географических районов [7–9]. Такие работы позволяют рассмотреть влияние глобального эксперимента, проведенного в прошлом столетии, на генетическую структуру соболя конкретного региона.
Интерес к сравнению современной популяционной структуры соболей Буреинского нагорья (левобережье Амура) и восточной части хребта Сихотэ-Алинь (правобережье Амура) обусловлен рядом факторов. Наиболее значимые из них: история формирования ареала в данном регионе и генетическая специфика различных интродукций в ХХ в. Горный рельеф, долготное направление, приокеаническое положение и отсутствие покровного оледенения позволили Сихотэ-Алиню даже в самые холодные времена плейстоцена сохранить лесные рефугиумы [10]. К.Г. Абрамов первым предположил, что после последнего плейстоценового оледенения именно c этой территории происходило расселение соболя [11]. Исследование митохондриальной и ядерной ДНК подтвердило это предположение [6, 12, 13].
После антропогенно организованного падения численности начала XX в. очаги автохтонных соболей сохранились на Сихотэ-Алине лишь в недоступных человеку горных участках. На большей части Буреинского хребта соболи были также изведены. Однако на севере Буреинского хребта они сохранились в количестве достаточном для организации в 1952 г. Буреинского соболиного племенного рассадника, откуда зверьки были расселены в различные регионы России, в том числе по Буреинскому нагорью на юг, включая хребет Малый Хинган, и по Сихотэ-Алиню [14]. На Сихотэ-Алине, кроме того, были заселены зверьки из Иркутской области [15]. Исследование географических группировок соболей Среднего Приамурья, разделенных эколого-географическим барьером – р. Амур с прилежащей к ней Средне-Амурской низменностью, позволяет сопоставить популяционную структуру современных обитателей территории Буреинского соболиного рассадника, потомков интродуцентов на территории более южных районов Буреинского хребта и хребта Малый Хинган, а также обитателей восточных районов Сихотэ-Алиня – возможных потомков и гибридов Сихотэ-Алинской автохтонной формы и двух интродуцированных.
В силу большой сложности сбора материала исследование природных популяций соболя обычно опирается на суммарные выборки в пространстве (по ближайшим географическим районам) и во времени (сборы нескольких лет). Такой подход дает усредненные характеристики внутривидового разнообразия обитателей конкретного региона, не позволяя отследить внутрипопуляционные изменения во времени, которые могли бы стать базой для эколого-генетического исследования. В отличие от такого подхода генетический мониторинг охотничьих сборов – возможный источник информации, потенциально позволяющий приблизить нас к пониманию влияния антропогенного изъятия на генетическую структуру данного природного объекта в конкретном регионе в определенный отрезок времени.
Настоящая работа является начальным этапом мониторингового исследования соболя Среднего Приамурья. Цель работы – провести анализ изменения популяционной генетической структуры соболей Среднего Приамурья по материалу охотничьих сборов зимних сезонов 2011/2012–2020/2021 гг. Рассматриваются микросателлиты Ма3 и Mer041, по которым у соболей Сихотэ-Алиня ранее были обнаружены специфичные аллели в сравнении с обитателями как географически удаленных (Сибирь, Урал, Камчатка) [6], так и буреинской [16] частей ареала.
Проводимый мониторинг позволит оценить изменения в генетической структуре животных отдельных районов Среднего Приамурья, а также значимость выявленных маркеров для характеристики различий в популяциях соболя левого и правого берега р. Амур.
МАТЕРИАЛЫ И МЕТОДЫ
Материал от 565 особей из четырех географических районов предоставлен охотниками, имеющими лицензии на отлов соболей. Ни одно животное не было отловлено специально для данной работы. Полученный материал охватывает охотничьи сезоны от 2011/2012 до 2020/2021 гг. Исследуемые субпопуляции, локализованные на Буреинском нагорье, расположенном по левую сторону от р. Амур, условно обозначены нами как “Маган”, “Сутырь” и “Каменушка”; по правую сторону – “Сихотэ-Алинь”. Субпопуляция “Маган” расположена на территории ранее существовавшего Буреинского соболиного племенного рассадника. На рис. 1 и в табл. 1 представлено географическое происхождение, посезонное и численное распределение собранного материала.

Таблица 1.
Географическая локализация и количество исследуемого материала соболя по сезонам
| Район сбора материала | Общее число животных | Количество животных в отдельные охотничьи сезоны | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2011–2012 | 2012–2013 | 2013–2014 | 2014–2015 | 2015–2016 | 2017–2018 | 2019–2020 | 2020–2021 | ||
| Северо-западный склон Буреинского хребта, район верховьев р. Бурея – “Маган” | 148 | – | 14 | 21 | 51 | – | – | 43 | 19 |
| Западный склон Буреинского хребта в его средней части, район рек Тырма, Сутырь – правых притоков р. Бурея – “Сутырь” | 146 | – | 51 | 30 | 35 | 30 | – | – | – |
| Южная оконечность Буреинского хребта и северо-восточный склон хребта Малый Хинган, район р. Бира – “Каменушка” | 148 | 19 | 15 | 49 | 34 | – | – | 31 | |
| Западный склон хребта Сихотэ-Алинь в его средней части, район рек Манома, Хар – “Сихотэ-Алинь” | 123 | – | 30 | 28 | – | 16 | 23 | 26 | – |
| Суммарно | 565 | 19 | 110 | 128 | 86 | 80 | 23 | 69 | 50 |
ДНК для исследования полиморфизма микросателлитных локусов выделяли из фиксированных в спирте тканей с использованием стандартного метода солевой экстракции [17]. Режим амплификации и последовательности примененных праймеров взяты из работы С.Н. Каштанова с соавт. [18]. Участки ДНК, соответствующие выбранным локусам, амплифицировали с помощью прямого и обратного праймеров производства ООО “Бигль” (Санкт-Петербург). Один из них был флуоресцентно помечен по 5'-концу красителем 6-FAM. Анализ длин фрагментов проводили на восьмиканальном генетическом анализаторе АВ-3500 в гель-полимере РОР 7 и в присутствии маркера молекулярной массы LIZ 600. Идентификацию локуса проводили с помощью программы Gene Mapper, версия 4.1 (Applied Biosystems, США). Аллели рассматриваемых локусов в тексте обозначены цифрами их длины с нижним подчеркиванием.
Оценка результатов изучения полиморфизма была проведена на основе анализа генотипических и аллельных частот. Оценки частот аллелей в географических выборках вычислялись по формуле [19]:
Если в выборке состояние равновесия нарушено, то
Оценки неравновесности сцепления генов, наблюдаемой и ожидаемой гетерозиготности, соответствия распределению Харди–Вайнберга, а также подсчет дистанций на основе попарных FST проводили в программе Arlequin (версия 3.5 2020) [20].
РЕЗУЛЬТАТЫ
Рассматриваемые локусы Ма3 и Mer041 не показали неравновесного сцепления (при уровне значимости 0.05), что указывает на их локализацию в различных хромосомах.
В суммарной выборке из 565 экземпляров обнаружено пять аллелей локуса Ма3: 125, 129, 131, 133 и 135. В выборках охотничьих сезонов географических субпопуляций Буреинского нагорья обнаружено по 3–4 аллеля и по 2–4 аллеля в сезонных выборках хребта Сихотэ-Алинь (табл. 2). В каждой из географических субпопуляций и во все сезоны основным (частота встречаемости >0.5) является аллель 131 (рис. 2). Второй по встречаемости аллель 133 также обнаружен во всех рассматриваемых субпопуляциях и во все сезоны. В субпопуляциях Буреинского нагорья сезонные изменения частоты этих аллелей лежат в пересекающейся области их стандартных ошибок. Исключение составляет субпопуляция “Маган”, где по частотам наиболее представленного аллеля 131 наблюдается разделение сезонных сборов на две группы, в каждой из которых частоты близки между собой, но обнаруживается падение частоты этого аллеля между сезонами 2013/2014 и 2014/2015. Однако согласно критерию хи-квадрат Пирсона нуль-гипотеза равенства частот аллеля 131 в этих сезонных выборках не отклоняется (уровень значимости различий 0.05 < р ≤ 0.1).
Таблица 2.
Генетические характеристики исследованных микросателлитных локусов
| Сезон | N | Ма3 | Мer041 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | НO | HE | pH-W | n | НO | HE | pH-W | ||
| Северо-западный склон Буреинского хребта, район верховьев р. Бурея – “Маган” | |||||||||
| 2012–2013 | 14 | 3 | 0.429 | 0.373 | 1 | 8 | 0.714 | 0.852 | 0.35 |
| 2013–2014 | 21 | 4 | 0.334 | 0.362 | 0.67 | 9 | 0.857 | 0.825 | 0.11 |
| 2014–2015 | 51 | 3 | 0.471 | 0.512 | 0.12 | 8 | 0.588 | 0.757 | 0.001 |
| 2019–2020 | 43 | 3 | 0.488 | 0.569 | 0.02 | 8 | 0.721 | 0.828 | 0.06 |
| 2020–2021 | 19 | 3 | 0.579 | 0.542 | 1 | 8 | 0.842 | 0.818 | 0.61 |
| Западный склон Буреинского хребта в его средней части, район рек Тырма, Сутырь – правых притоков р. Бурея – “Сутырь” | |||||||||
| 2012–2013 | 51 | 3 | 0.490 | 0.547 | 0.07 | 9 | 0.824 | 0.844 | 0.16 |
| 2013–2014 | 30 | 3 | 0.534 | 0.542 | 0.39 | 6 | 0.633 | 0.813 | 0.002 |
| 2014–2015 | 35 | 3 | 0.543 | 0.545 | 0.11 | 8 | 0.857 | 0.816 | 0.79 |
| 2015–2016 | 30 | 3 | 0.434 | 0.493 | 0.11 | 8 | 0.500 | 0.823 | 0.002 |
| Южная оконечность Буреинского хребта и северо-восточный склон хребта Малый Хинган, район р. Бира – “Каменушка” | |||||||||
| 2011–2012 | 19 | 3 | 0.474 | 0.482 | 1 | 6 | 0.842 | 0.784 | 0.33 |
| 2012–2013 | 15 | 3 | 0.467 | 0.476 | 0.25 | 8 | 0.933 | 0.860 | 0.89 |
| 2013–2014 | 49 | 4 | 0.33 | 0.49 | 0.002 | 8 | 0.57 | 0.81 | 0.015 |
| 2015–2016 | 34 | 3 | 0.382 | 0.453 | 0.21 | 8 | 0.706 | 0.797 | 0.36 |
| 2020–2021 | 31 | 3 | 0.355 | 0.484 | 0.14 | 9 | 0.774 | 0.821 | 0.20 |
| Западный склон хребта Сихотэ-Алинь в его средней части, район рек Манома, Хар – “Сихотэ-Алинь” | |||||||||
| 2012–2013 | 30 | 3 | 0.434 | 0.389 | 1 | 12 | 0.700 | 0.897 | 0.03 |
| 2013–2014 | 28 | 2 | 0.179 | 0.223 | 0.35 | 9 | 0.643 | 0.873 | 0.02 |
| 2015–2016 | 16 | 2 | 0.063 | 0.063 | 1 | 7 | 0.500 | 0.782 | 0.01 |
| 2017–2018 | 23 | 4 | 0.348 | 0.556 | 0.023 | 11 | 0.478 | 0.906 | 0 |
| 2019–2020 | 26 | 3 | 0.154 | 0.212 | 0.28 | 11 | 0.962 | 0.894 | 0.15 |
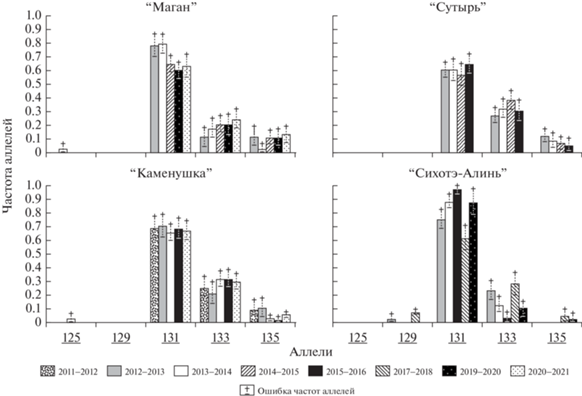
Иная картина представлена в субпопуляции “Сихотэ-Алинь”, где обнаружены резкие статистически значимые колебания частот этих аллелей от сезона к сезону. Достоверность этих различий обнаруживается при сравнении выборок пар сезонов 2015/2016 с 2017/2018 и 2017/2018 с 2019/2020 (уровень значимости равен 0.01). Наблюдаемые различия недостоверны при сравнении материала пар сезонов 2012/2013 с 2013/2014 и 2013/2014 с 2015/2016 (уровень значимости равен 0.1). Аналогичная картина межсезонного изменения частот обнаружена и для аллеля 133 этой субпопуляции.
Аллель 135, третий по встречаемости в субпопуляциях Буреинского нагорья, в более северных субпопуляциях “Маган” и “Сутырь” представлен в материале всех рассматриваемых сезонов с изменениями частоты в основном в пределах стандартной ошибки. В южной субпопуляции “Каменушка” наблюдается статистически достоверное изменение частоты этого аллеля. Он значимо представлен в материале сезонов 2011/2012–2012/2013 и обнаруживается как редкий в выборках последующих сезонов. Данный аллель, определенный как специфичный для субпопуляций Буреинского нагорья по сборам охотничьего сезона 2012/2013 [16], появляется в качестве редкого на Сихотэ-Алине в охотничьи сезоны 2017/2018 и 2019/2020 (рис. 2).
Аллель 129 обнаружен только в Сихотэ-Алинской субпопуляции в сборах двух сезонов, с частотой встречаемости, доходящей до 0.07.
Редкий аллель 125 обнаружен только на Буреинском нагорье в субпопуляциях “Маган” и “Каменушка”.
В суммарной выборке из 565 экземпляров обнаружено 13 аллельных вариантов локуса Меr041: 138, 140, 142, 144, 146, 148, 150, 152, 154, 156, 158, 160 и 162 (рис. 3, 4). В сезонных выборках соболя Буреинского нагорья представлено от шести до девяти аллелей (в среднем по 8.1 на субпопуляцию за сезон). В Сихотэ-Алинских сезонных сборах встречается от 7 до 12 аллелей (в среднем 10). Данный локус нередко показывает значимые колебания частот аллелей от сезона к сезону. Но практически во всех сезонных выборках севера (“Маган”) и центральной части (“Сутырь”) Буреинского нагорья наиболее часто встречающимся является аллель 146. В южной субпопуляции (“Каменушка”) этот аллель наиболее часто встречается в сезонах 2013/2014, 2015/2016, 2020/2021 г. В другие сезоны преобладающими являются аллели 140 (2011/2012) и 150 (2012/2013). Иная картина наблюдается в Сихотэ-Алинской субпопуляции. Наиболее часто встречающийся аллель здесь, как правило, меняется от сезона к сезону.

Рис. 4.
Распределение частот аллелей локуса Mer041 по сезонам в субпопуляциях “Каменушка” и “Сихотэ-Алинь”.

По данным исследования соболей охотничьего сезона 2012/2013 редкие аллели 138, 160 и 162 являлись уникальными для Сихотэ-Алинской субпопуляции. По результатам мониторинга обнаружено присутствие аллеля 138 в Сихотэ-Алинской субпопуляции в четырех из пяти рассматриваемых сезонов. Однако этот аллель встречен и у соболей севера Буреинского хребта в сезоны 2019/2020 и 2020/2021. Редкий аллель 160 был обнаружен в Сихотэ-Алинской субпопуляции в составе материала трех сезонов: 2012/2013, 2017/2018 и 2019/2020. Также он был обнаружен в один из сезонов и на севере Буреинского хребта. По данным проведенного мониторинга лишь один аллель этого локуса 162 остался маркерным для Сихотэ-Алинской субпопуляции и был повторно встречен здесь в сезон 2019/2020 гг.
Сравнение величин наблюдаемой и ожидаемой гетерозиготности (табл. 2) показывает некоторый недостаток гетерозигот в большинстве из 19 рассматриваемых географических сезонных выборок. Но для локуса Ма3 наблюдаемый недостаток гетерозигот, как правило, незначителен и большая часть сезонных географических выборок (16 из 19) показывает соответствие распределения генотипов соотношению Харди–Вайнберга (на уровне значимости 0.05). Нарушение равновесия по этому локусу наблюдается на севере Буреинского хребта в сезон 2019/2020. Для популяции юга Буреинского хребта неравновесными оказываются сборы сезона 2013/2014, который представлен наибольшим количеством локалитетов. Здесь резко падает частота аллеля 135 и появляется аллель 125. Для Сихотэ-Алинской субпопуляции неравновесной оказывается выборка сезона 2017/2018, характеризуемая появлением аллеля 135. Для локуса Меr041 в субпопуляциях Буреинского нагорья наблюдаемый недостаток гетерозигот, как правило, также незначителен, а большая часть сезонных географических выборок (10 из 14) показывает соответствие распределения генотипов соотношению Харди–Вайнберга (на уровне значимости 0.05). Иная картина наблюдается в Сихотэ-Алинской субпопуляции. Для сборов четырех сезонов (с 2012/2013 по 2017/2018) в районе р. Манома обнаружен значимый недостаток гетерозигот, приведший к отклонению от распределения Харди–Вайнберга. Можно предположить, что наблюдаемое отклонение определяется случайностью выявления аллелей при относительно небольших сезонных выборках и высоком аллельном разнообразии. Однако необъяснимо в таком случае почему оно затрагивает именно гетерозиготные генотипы, и более того, на р. Хар – в соседнем локалитете с р. Манома (сезон 2019/2020) при таком же аллельном разнообразии значимого недостатка гетерозигот не наблюдается.
В табл. 3 указан размах значений генетических дистанций, полученных на основе попарных сравнений – FST: для сезонных сборов каждой субпопуляции, для сезонных сборов субпопуляций Буреинского нагорья между собой и с субпопуляцией “Сихотэ-Алинь”. Минимальные показатели FST отражают нулевой уровень дифференциации сезонных сборов всех групп сравнений. FST для сравнения сезонных сборов различных географических субпопуляций Буреинского нагорья не превышают 0.051. При этом практически для всех таких сравнений не отвергается гипотеза нулевого уровня их дифференциации. Максимальные значения показателей генетической дифференциации сборов различных сезонов Сихотэ-Алинской субпопуляции (0.08) и попарных сравнений субпопуляций Буреинского нагорья с Сихотэ-Алинской субпопуляцией (0.102–0.138) отражают достоверно отличный от нуля уровень их генетической дифференциации.
Таблица 3.
Разброс межсезонных значений генетических дистанций (по попарным FST для географических субпопуляций соболя Среднего Приамурья)
| Субпопуляции | “Маган” | “Сутырь” | “Каменушка” | “Сихотэ-Алинь” |
|---|---|---|---|---|
| “Маган” | 0.028–0.002 | |||
| “Сутырь” | 0.051–0.001 | 0.008–0.001 | ||
| “Каменушка” | 0.048–0.001 | 0.021–0.001 | 0.028–0.003 | |
| “Сихотэ-Алинь” | 0.102–0.003 | 0.138–0.007 | 0.132–0.002 | 0.084–0.004 |
ОБСУЖДЕНИЕ
Высокий уровень аллельного разнообразия у соболей Сихотэ-Алиня [6], предполагающий их расселение в постледниковье из Сихотэ-Алинского лесного рефугиума, привлекает внимание к популяционной структуре обитателей левобережья Амура, территории первых шагов такого расселения. Результаты исследования биологического материала одного охотничьего сезона (2012/2013) показали, что вывод о более высоком уровне аллельного разнообразия соболей Сихотэ-Алиня верен и для сравнения субпопуляции западного макросклона Сихотэ-Алиня с субпопуляциями Буреинского нагорья [16]. Однако мониторинг восьми охотничьих сезонов периода 2011–2021 гг. обнаружил более сложную картину. При работе с материалом 2013–2021 гг. у соболей Буреинского нагорья в отдельные сезоны были выявлены редкие аллели (138, 142, 160 локуса Меr041 и 125 локуса Ма3), в общем не изменяющие картины более высокого аллельного разнообразия Сихотэ-Алинской субпопуляции по сравнению с Буреинскими (табл. 2). Но обнаружены эти редкие аллели главным образом в субпопуляции “Маган”. Суммирование для каждой из субпопуляций данных по всем сезонам показывает сопоставимо высокое аллельное разнообразие в субпопуляциях “Маган”, автохтонной для соболей Буреинского нагорья и “Сихотэ-Алинь”, где возможна встреча трех форм – автохтонной Сихотэ-Алинской и двух интродуцированных. Уменьшение аллельного разнообразия в субпопуляциях “Сутырь” и “Каменушка” в сравнении с “Маган” возможно объясняется эффектом основателя при заселении соболей из Верхнебуреинского рассадника в более южные районы Буреинского нагорья.
Проведенный мониторинг показал, что Сихотэ-Алинскую субпопуляцию маркируют два уникальных аллеля – 129 локуса Ма3 и 162 локуса Меr041. Эти аллели обнаружены здесь в выборках нескольких охотничьих сезонов с частотой встречаемости, доходящей до 0.07. Являются ли эти аллели оба, либо какой-нибудь из них характеристикой автохтонной популяции соболей Сихотэ-Алиня, либо занесены (занесен) во время интродукции соболей из Сибири – можно понять при сравнении со зверьками из Иркутской популяции. К сожалению, в нашем распоряжении подобного материала не было. Прямое сопоставление микросателлитных аллелей, полученных на различающейся приборной базе, практически невозможно, особенно для высокополиморфных локусов, к которым относится Меr041. Для наименее полиморфного локуса Ма3 можно попытаться провести аналогию с ранее полученными данными. В промышленной популяции соболя было обнаружено четыре аллеля Ма3 с диапазоном длин их спектра: 131, 133, 135, 137 [21]. У соболей Среднего Приамурья при работе в тех же условиях амплификации нами обнаружены четыре аллеля, определяемые на нашей приборной базе как 129, 131, 133, 135. Кроме того, здесь был обнаружен редкий аллель 125, длина которого на два повтора меньше самого короткого (129) из указанного спектра четырех. Вполне вероятно, что при исследовании промышленной популяции соболя и природных популяций Среднего Приамурья речь идет об одном и том же спектре четырех аллелей. Отсюда следует, что самый длинный аллель выявляемого нами спектра (135) идентичен аллелю 137, являющемуся уникальным для Сихотэ-Алинской популяции по данным С.Н. Каштанова с соавт. [6]. В таком случае характеристикой автохтонной популяции Сихотэ-Алиня является аллель 135. В настоящее время он включен в геном Буреинских и Сихотэ-Алинской субпопуляций. Аллель 129, обнаруженный нами в Среднем Приамурье только на Сихотэ-Алине, мог быть привнесен при интродукции соболей из Иркутской области. Конечно же, эта гипотеза требует инструментальной проверки. Однако если предположение верно, то аллель 129 более чем за полвека пребывания на правобережье Амура не проник на левобережье, что подтверждает значимость Амура с прилежащей Средне-Амурской низменностью в качестве изолирующего эколого-географического барьера даже для активно и далеко мигрирующих соболей. Другим примером, иллюстрирующим подразделенность лево- и правобережных групп соболей, могли бы быть аллели 125 локуса Ма3 и 142 локуса Меr041, встреченные только на Буреинском нагорье. За весь период исследования они обнаружены здесь только у одного либо двух зверьков. Подтверждение их приуроченности только к левобережью Амура требует проведения дальнейшего исследования.
Мониторинг аллельного состава микросателлитов в изымаемом охотниками материале представляет доступный источник информации о динамике генетической популяционной структуры этого природного промыслового вида в каждом из рассматриваемых географических районов региона. Обнаружен различающийся характер динамики популяционной структуры соболя лево- и правобережья р. Амур. Проведенное исследование двух микросателлитных локусов в материале охотничьих сборов 11 лет показывает относительную однородность популяционной структуры соболя Буреинского нагорья, как правило, не приводящую к статистически значимому уровню дифференциации субпопуляций ни в пространстве, ни во времени. По контрасту с ней гетерогенность Сихотэ-Алинской субпопуляции выявляется по частотам аллелей двух рассматриваемых локусов, по количеству этих аллелей в сезонных сборах и постоянно определяемым неравновесным состоянием по локусу Меr041 в локалитете “Манома”. Именно выявленная гетерогенность Сихотэ-Алинской субпопуляции определяет разброс генетических дистанций при сравнении сезонных сборов лево- и правобережья Амура от практически нулевых значений до промежуточного уровня дифференциации по Райту [22].
Существует множество факторов, приводящих к генетической гетерогенности во временных сборах в одной и той же популяции. Прежде всего это изменение численности, которое может быть обусловлено природными факторами, такими как динамика урожайности кедра, миграционной активностью, а также антропогенным давлением, в том числе промыслом соболя в конкретном районе обитания. Учитывая высокую подвижность соболей, можно предположить, что весомый вклад в изменение популяционной структуры мог быть внесен миграционной составляющей. Особенно вероятен такой процесс в районах, находящихся в шаговой доступности от человеческих поселений и потенциально подверженных в какие-то сезоны большой охотничьей активности, приводящей к освобождению мест для новых поселенцев – мигрантов с несколько отличающейся генетической структурой.
Наблюдаемая в локалитете “Манома” межсезонная гетерогенность аллельного состава может указывать на существование на склонах Сихотэ-Алиня пространственно неоднородной популяционной структуры, сформированной из потомков и возможных гибридов автохтонной и двух интродуцированных форм соболя. Предоставленный охотниками материал мог быть собран на стыке обитания этих форм, либо при неравномерном облове различающихся субпопуляций по сезонам. Другое объяснение предполагает возникновение гетерогенности от сезона к сезону за счет потоков мигрантов с различающейся генетической структурой на свободные территории, возможно появляющиеся в том числе в результате промысла.
Рассматриваемые микросателлитные маркеры являются показательными в исследовании дифференциации и мониторинге изменений в популяционной структуре соболя Среднего Приамурья. Продолжение мониторинговых исследований, расширение района исследования и набора маркерных характеристик, как генетических, так и морфологических, позволит воссоздать полную картину эколого-генетических изменений во времени и пространстве.
Работа выполнена в рамках Госзадания ИКАРП ДВО РАН № 075-01570-22-00.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Балмышева Н.П., Соловенчук Л.Л. Генетическая изменчивость гена цитохрома b митохондриальной ДНК соболя (Martes zibellina L.) магаданской популяции // Генетика. 1999. Т. 35. № 9. С. 1252–1257.
Петровская А.В. Генетическая структура популяций соболя (Martes zibellina L.) в Магаданской области по данным об изменчивости митохондриальной ДНК // Генетика. 2007. Т. 43. № 4. С. 530–536.
Рожнов В.В., Пищулина С.Л., Мещерский И.Г. и др. Генетическая структура соболя (Martes zibellina L.) Евразии и анализ распределения митохондриальных линий // Генетика. 2013. Т. 49. № 2. С. 251–258.
Kinoshita Gohta, Sato J.J., Meschersky I.G. et al. Colonization history of the sable Martes zibellina (Mammalia, Carnivora) on the marginal peninsula and islands of northeastern Eurasia // J. Mammalogy. 2015. № 96(1). P. 172–184. https://doi.org/10.1093/jmamma/gyu021
Каштанов С.Н., Свищева Г.Р., Лазебный О.Е. и др. Влияние антропогенных факторов на генетическое разнообразие вида соболь (Martes zibellina L.) // Мол. биология. 2015. Т. 49. № 3. С. 449–454. https://doi.org/10.7868/S002689841503009X
Каштанов С.Н., Свищева Г.Р., Пищулина С.Л. и др. Географическая структура генофонда соболя (Martes zibellina L.): данные анализа микросателлитеых локусов // Генетика 2015. Т. 51. № 1. С. 78–88. https://doi.org/10.7868/S001667581501004X
Каштанов С.Н., Столповский Ю.А., Мещерский И.Г. и др. Таксономический статус и генетическая идентификация соболя Алтая (Martes zibellina averini Bazhanov, 1943) // Генетика. 2018. Т. 54. № 11. С. 1327–1337. https://doi.org/10.1134/S0016675818110073
Монахов В.Г., Модоров М.В., Ранюк М.Н. Генетические последствия реинтродукции соболя (Martes zibellina L.) в Западной Сибири // Генетика. 2018. Т. 54. № 3. С. 361–365. https://doi.org/10.7868/S0016675818030098
Монахов В.Г., Ранюк М.Н., Модоров М.В. Популяционная структура Байкальской горной страны. Анализ генетических и фенотипических характеристик // Экология. 2021. № 2. С. 143–152. https://doi.org/10.31857/S0367059721020074
Крестов П.В., Баркалов В.Ю., Омелько А.М. и др. Реликтовые комплексы растительности современных рефугиумов северо-восточной Азии // Комаровские чтения. Владивосток: Дальнаука, 2009. Вып. 56. С. 5–63.
Абрамов К.Г. Соболь в охотничьем хозяйстве Дальнего Востока. М.: Наука, 1967. 116 с.
Sato J., Hosoda T., Kryukov A. et al. Genetic diversity of the sable (Martes zibellina, Mustelidae) in Russian Far East and Hokkaido inferred from mitochondrial NADH dehydrogenase subunit 2 gene sequences // Mammal. Study. 2011. V. 36. P. 209–222.
Li B., Malyarchuk B., Ma Z. et al. Phylogeography of sable (Martes zibellina L. 1758) in the southeast portion of its range based on mitochondrial DNA variation: highlighting the evolutionary history of the sable // Acta Theriologica. 2013. V. 58. P. 139–148.
Голобокова О.А. История создания Верхнебуреинского соболиного племенного рассадника. 2017. URL: https://komza.khabkrai.ru/Municipalnye-arhivy/Novosti-/890 (дата обращения: 22.11.2019).
Каплин А.А. Пушнина СССР. М.: Внешторгиздат, 1960. 464 с.
Фрисман Л.В., Шлюфман К.В., Брыкова А.Л. Генетическая структура популяций соболя Буреинского хребта по сборам одного охотничьего сезона // Региональные проблемы. 2019. Т. 22. № 4. С. 46–55. https://doi.org/10.31433/2618-9593-2019-22-4-46-55
Aljanabi S.M., Martinez I. Universal and rapid salt-extraction of high quality genomic DNAb for PCR-based techniques // Nucl. Acids Res. 1997. V. 25. № 22. P. 4692–4693.
Каштанов С.Н., Рубцова Г.А., Лазебный О.Е. Исследование генетической структуры промышленной популяции соболя (Martes zibellina Linnaeus,1758) по микросателитным маркерам // Вестник ВОГиС. 2010. Т. 14. № 3. С. 426–431.
Кузнецов В.М. F-статистика Райта: оценка и интерпретация // Проблемы биологии продуктивных животных. 2014. № 4. С. 80–104.
Excoffier L., Laval G., Schneider C. Arlequin. Ver 3.0. An integrated software package for population genetics data analysis // Evolutionary Bioinformatic Online. 2005. V. 1. P. 47–50.
Каштанов С.Н., Афанасьев К.И., Потапов С.Г., Лазебный О.Е. Микросателлитный анализ двух промышленных популяций соболя (Martes zibellina L.) // Генетика. 2011. Т. 47. № 12. С. 1622–1628.
Wright S. Evolution and Genetics of Population. V. 4. Variability Within and Among Natural Populations. Chicago: Univ. Chicago Press, 1978. 590 p.
Дополнительные материалы отсутствуют.