

Вопросы ихтиологии, 2020, T. 60, № 1, стр. 95-105
Вкусовые рецепторы ротовой полости у молоди карпа Cyprinus carpio и беломорской трески Gadus morhua marisalbi
Г. В. Девицина 1, *, Т. В. Головкина 1
1 Московский государственный университет
Москва, Россия
* E-mail: gdevicyna@mail.ru
Поступила в редакцию 24.12.2018
После доработки 26.02.2019
Принята к публикации 19.04.2019
Аннотация
Методом сканирующей электронной микроскопии проведено сравнительное исследование особенностей морфологии интраорального вкусового аппарата у одновозрастной молоди карпа Cyprinus carpio и беломорской трески Gadus morhua marisalbi. Показано, что молодь этих двух видов рыб обладает сходной системой зонального распределения вкусовых рецепторов в ротовой полости на три взаимосвязанных отдела. Взаимодействие этих отделов обеспечивает сложную многоступенчатую систему вкусового анализа пищевых объектов. Особенности морфологии вкусового рецепторного аппарата в одноимённых сенсорных зонах ротовой полости у молоди карпа и трески отражают сходство их адаптации к однотипному характеру питания как планктонными, так и бентосными объектами. Выявлены видоспецифичные особенности в морфологии сенсорного поля вкусовых почек. Ряд структурных различий в морфологии одноимённых сенсорных зон ротовой полости молоди карпа и трески имеют адаптивную направленность в связи с видовой спецификой процессов вкусового тестирования.
В сенсорном обеспечении рыб как обитателей водной среды особенно важную роль играют системы химической рецепции. Рыбы обладают тремя хемосенсорными системами – обонятельной, системой общего химического чувства и вкусовой, включающей, в свою очередь, две субсистемы – наружную и внутриротовую (Finger, Morita, 1990; Павлов, Касумян, 1990; Devitsina, Chervova, 1994). Наружная вкусовая субсистема присутствует не у всех видов рыб. Она участвует в поисках пищевых объектов (Atema, 1971). Внутриротовая вкусовая субсистема в отличие от наружной присутствует у всех видов рыб; её роль заключается в определении пригодности пищевого объекта. Степень её развития и особенности морфологии связаны с характером питания вида (Певзнер, 1985; Касумян, 1997). Каждый вид имеет специфические вкусовые предпочтения (Kasumyan, Døving, 2003). Избирательность питания у особей каждого вида связана с морфологическими и функциональными особенностями внутриротовой вкусовой субсистемы. Структурная организация вкусового рецепторного аппарата в ротовой полости рыб видоспецифична (Western, 1969; Devitsina, 2005).
В морфологии внутриротовой вкусовой субсистемы у рыб проявляются возрастные изменения, связанные с характером питания молоди (Андрияшев, 1954; Касумян, Пономарёв, 1990; Девицина, Головкина, 2011; Devitsina et al., 2011). В процессе онтогенеза у рыб меняется характер питания. Каждый этап характеризуется своей спецификой пищевых отношений и своим кормовым коэффициентом (Суворов, 1948; Никольский, 1971). При высокой видовой специфичности питания у рыб на определённых этапах индивидуального развития может проявляться сходство в выборе объектов питания. Это происходит одновременно с морфологическими и функциональными изменениями в различных сенсорных системах, в том числе и вкусовой. Видовая и возрастная специфика в характере питания тесно связана с особенностями морфологии и функциональных свойств внутриротовой вкусовой рецепции у рыб. Проблема межвидовой и возрастной специфики в структуре вкусового рецепторного аппарата ротовой полости у рыб до последнего времени остаётся мало изученной и представляет значительный интерес.
Цель настоящей работы ‒ сравнительное исследование особенностей структурной организации вкусового рецепторного аппарата в ротовой полости у одновозрастной молоди двух видов рыб, населяющих придонные биотопы в водоёмах разного характера, − обыкновенного карпа Cyprinus carpio (Cyprinidae) и беломорской трески Gadus morhua marisalbi (Gadidae).
МАТЕРИАЛ И МЕТОДИКА
Молодь карпа (11 особей длиной 8–9 см, возраст 1+) приобрели во Всероссийском научно-производственном объединении по рыбоводству (ВНИИПРХ, Московская область). Рыб содержали в аквариумах при температуре 18–20°С и кормили мотылём (Chironomidae). Молодь трески (10 особей длиной 10–11 см, возраст 1+) отловили в прибрежной зоне Кандалакшского залива Белого моря. Рыб содержали в сетчатом садке, установленном в море на глубине 1.0–1.5 м в течение 5–6 сут, при температуре 7–8°С и подкармливали пескожилом Arenicola marina.
Для смыва с поверхности эпителия кожной слизи каждую из опытных рыб в течение 10 мин выдерживали в охлаждённом (9–11°С) 9%-ном растворе хлористого натрия, затем отмывали охлаждённой чистой водой, плотно обёртывали мокрой марлей и рассекали спинной мозг позади черепа. Отпрепарированный эпителий ротовой полости фиксировали в охлаждённом 2.5%-ном растворе глютарового альдегида, приготовленном на 0.1 М фосфатном буфере (рН 7.4). Фиксатор отмывали тем же буфером с добавлением сахарозы. Особенности применения этого метода для тканей рыб описаны ранее (Девицина, 1990). Материал обезвоживали в ряду этанол–ацетон, высушивали при критической точке на аппарате Hitachi HCP–1 (Япония), затем монтировали на металлические столики и напыляли смесью золото–палладий. На препаратах измеряли площадь сенсорного поля вкусовых почек (в мкм2), а также плотность распределения вкусовых почек в эпителии вкусовых зон (шт/мм2) с помощью компьютерной программы Image-Pro. Фотографии препаратов для измерения площадей делали при ориентации их перпендикулярно окуляру микроскопа. Морфологию вкусового рецепторного аппарата в слизистой выстилке ротовой полости изучали с помощью электронных сканирующих микроскопов CamScan S–2 (“Cambridge Intstruments”, Великобритания) или JSM–6380 LA (“Jeol”, Япония).
У опытных рыб изучали морфологию и ультраструктуру сенсорного поля вкусовых почек, особенности их распределения в слизистой выстилке ротовой полости. Проводили сравнительный анализ распределения комплексов различных эпидермальных структур (гребни, сосочки, папиллы), которые обеспечивают оптимальные условия функционирования вкусовых рецепторов. Для статистической обработки результатов применяли непараметрический критерий Манна–Уитни через программу Statistica-7.
РЕЗУЛЬТАТЫ
Анализ распределения вкусовых рецепторов в слизистой выстилке ротовой полости рыб показал, что у обоих исследованных видов рецепторы распределяются сходным образом по зональному принципу.
Карп имеет в ротовой полости девять вкусовых зон, различающихся особенностями морфологии: внутренней поверхностью верхней и нижней губных складок; премаксиллярной и максиллярной зонами; зонами твёрдого и мягкого нёба, языком, гулярной (горловой) и дентальной (Девицина, Головкина, 2007). Дорсальный свод ротовой полости имеет вкусовые зоны верхней губы, премаксиллярную, максиллярную зоны, зоны твёрдого и мягкого нёба.
Верхняя губа на внутриротовой поверхности несёт многочисленные вкусовые поры, в которых находятся вкусовые почки погружённого типа (рис. 1, 2). Вкусовые почки в эпителии верхней губы имеют сенсорное поле ниже поверхности эпителия (рис. 1а, 1б). Плотность распределения вкусовых почек на верхней губе (370 шт/мм2) достоверно выше, чем в других зонах (за исключением зоны мягкого нёба) (рис. 2б). Площадь сенсорного поля вкусовых почек составляет 27–34 (в среднем 29) мкм2 (рис. 2а).
Рис. 1.
Вкусовые зоны верхней челюсти карпа Cyprinus carpio: а – внутренняя поверхность верхней губы с вкусовыми порами, б – вкусовая пора с вкусовой
почкой погружённого типа на верхней губе, в – премаксиллярная зона (1) с крупными вкусовыми сосочками и вкусовыми почками и максиллярная зона (2) с вкусовыми почками, г – вкусовая почка на сосочке премаксиллярной зоны; (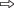 ) – вкусовые почки, (
) – вкусовые почки, ( ) – вкусовые сосочки с вкусовыми почками. Масштаб: а – 100, б – 10, в – 150, г – 15
мкм.
) – вкусовые сосочки с вкусовыми почками. Масштаб: а – 100, б – 10, в – 150, г – 15
мкм.
Рис. 2.
Морфометрические характеристики вкусовых зон ротовой полости карпа Cyprinus carpio (а, б) и беломорской трески Gadus morhua marisalbi (в, г): а, в – площадь сенсорного поля вкусовых почек; б, г – плотность распределения вкусовых почек. Сенсорные зоны: 1 – верхняя губа, 2 – премаксиллярная зона, 3 – максиллярная зона, 4 – твёрдое нёбо, 5 – язык, 6 – гулярная зона, 7 – дентальная зона, 8 – нижняя губа, 9 – мягкое нёбо; *отличия от остальных зон достоверны при p < 0.05.
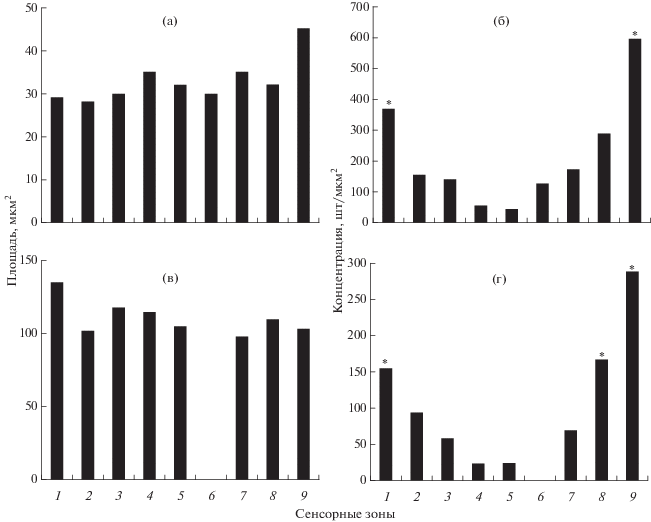
Премаксиллярная зона представлена эпителиальными покровами premaxillare. Средняя плотность распределения вкусовых почек в этой зоне составляет 155 шт/мм2. Эта зона отличается крупными вкусовыми сосочками с плоской вершиной, которые образуют правильный рисунок (рис. 1в). Каждый сосочек площадью 0.03–0.10 мм2 несёт от 5 до 15 вкусовых почек с размером сенсорного поля 20–32 (28) мкм2. Сенсорное поле этих почек находится на уровне или выше поверхности эпидермиса. Сенсорное поле вкусовых почек образовано рецепторными клетками поливиллярного и моновиллярного типов (рис. 1г). Моновиллярные клетки имеют рецепторную микровиллу разной длины.
Максиллярная зона расположена на maxillare и несёт разновеликие удлинённые поперечно ориентированные вкусовые сосочки (рис. 1в). На каждом сосочке несколько вкусовых почек, соразмерных вкусовым почкам премаксиллярной зоны. Средняя плотность распределения вкусовых почек − 140 шт/мм2.
Зона твёрдого нёба примыкает к максиллярной зоне и имеет продольно-ребристую поверхность. Вкусовые почки равномерно распределяются вдоль тонких нёбных рёбер по одной на небольших эпидермальных папиллах. Средняя плотность распределения вкусовых папилл в этой зоне составляет 55 шт/мм2; средняя площадь сенсорного поля отдельных вкусовых почек − 35 мкм2 (рис. 2а, 2б).
Зона мягкого нёба представляет собой специфическое скопление крупных вкусовых сосочков (рис. 3). Для неё характерна бугристая структура, образованная плотно упакованными крупными вкусовыми сосочками сферической формы, несущими по 8–12 вкусовых почек (рис. 3а). В этой зоне преобладают вкусовые почки непогружённого типа, морфологически сходные с почками твёрдого нёба (рис. 3б, 3г). В эпителии мягкого нёба много одиночных сенсорных клеток, несущих пучок длинных апикальных отростков (рис. 3в). В разных участках этой зоны площадь сенсорного поля почек и их концентрация варьируют в широких пределах, составляя в среднем соответственно 45 мкм2 и 600 шт/мм2 (рис. 2б, 2а).
Рис. 3.
Нёбные зоны карпа Cyprinus carpio: а – вкусовые сосочки мягкого нёба с вкусовыми почками, б – вкусовая почка мягкого нёба, в – поверхность эпителия мягкого нёба с одиночными сенсорными клетками, г – вкусовая почка твёрдого нёба; (Δ) – вершины одиночных мультивиллярных клеток; ост. обозначения см. на рис. 1. Масштаб: а – 600, б – 40, в – 10, г – 60 мкм.
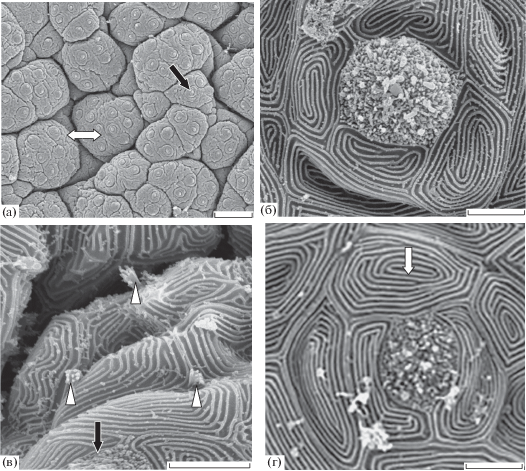
Вентральный свод ротовой полости карпа включает нижнюю губу, дентальную, гулярную зоны и язык (рис. 4а). Нижняя губа на внутриротовой поверхности имеет вкусовые почки непогружённого типа. Плотность их распределения (290 шт/мм2) меньше, чем на верхней губе. Размеры сенсорного поля вкусовых почек составляют 23−34 (32) мкм2. На поверхности нижней губы карпа многочисленны вершины одиночных сенсорных клеток (рис. 4б).
Рис. 4.
Нижняя челюсть карпа Cyprinus carpio: а – расположение сенсорных зон, б – вершины одиночных хемосенсорных клеток и вкусовые почки на поверхности эпителия нижней губы, в – поверхность вкусового сосочка гулярной зоны с вкусовыми почками погружённого типа, г – вкусовая почка погружённого типа в гулярной зоне; 3 – нижняя губа, 4 – дентальная зона, 5 – гулярная зона; ост. обозначения см. на рис. 1, 3. Масштаб: а – 300, б – 50, в – 100, г – 10 мкм.
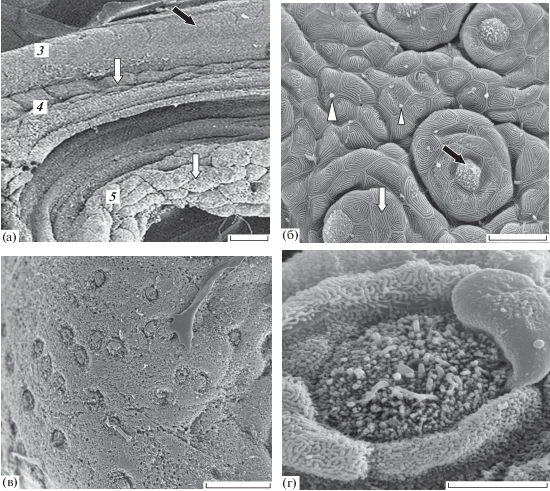
Дентальная зона образована цепью крупных вкусовых сосочков в слизистой выстилке dentale. Каудальный край дентальной зоны образует эпителиальный тяж с вкусовыми почками, площадь которых в среднем составляет 35 мкм2, плотность распределения − 172 шт/мм2 (рис. 2а, 2б).
Гулярная зона хорошо выражена и имеет вид изолированной, латерально ориентированной широкой полосы крупных плотно сжатых вкусовых сосочков сферической формы (рис. 4а, 4в). Каждый сосочек несёт 8–12 вкусовых почек; средняя площадь их сенсорного поля составляет 30 мкм2, плотность распределения − 125 шт/мм2. Вкусовые почки этой зоны имеют сенсорное поле погружённого типа (рис. 4в, 4г).
Язык представляет крупную вкусовую зону, которая имеет перистое расположение тонких дермальных гребней с высокими вкусовыми сосочками. Здесь вкусовые почки располагаются на вершине столбовидных эпидермальных сосочков. Средняя плотность распределения вкусовых почек на языке 43 шт/мм2; наиболее велика их концентрация на кончике языка. Преобладают сравнительно мелкие вкусовые почки. Сенсорное поле лежит выше поверхности эпителия, его площадь варьирует в пределах 20−40 (32) мкм2. Слизистая выстилка корня языка продолжается в глоточной зоне. Она имеет уплощённую поверхность и редкие небольшие вкусовые почки.
Морфология сенсорного поля вкусовых почек карпа сходна во всех сенсорных зонах ротовой полости. Сенсорное поле образовано микровиллами поливиллярных и моновиллярных рецепторных клеток.
Треска имеет в ротовой полости хорошо развитый вкусовой рецепторный аппарат (рис. 5), характеризующийся особенно крупными вкусовыми почками (~ 100 мкм2). Вкусовой аппарат представляют восемь морфологически различных сенсорных зон: внутренняя поверхность верхней губы, поверхность нижней губы, премаксиллярная и максиллярная зоны, дентальная зона, зоны твёрдого и мягкого нёба, язык (Девицина, Головкина, 2008). Характерной чертой вкусовых зон на верхней и нижней челюсти является наличие многочисленных мелких зубов, связанных с костями челюстной дуги – premaxillare и dentale (рис. 5а, 5в). Дорсальный свод ротовой полости трески включает верхнюю губу, премаксиллярную и максиллярную зоны, твёрдое и мягкое нёбо.
Рис. 5.
Вкусовые зоны рострального отдела ротовой полости трески Gadus morhua marisalbi: а – верхнечелюстные зоны: премаксиллярная, максиллярная и верхняя губа (6); б – вкусовая почка на сосочке премаксиллярной зоны; в – нижнечелюстные зоны: нижняя губа и дентальная зона; г – вкусовая почка на нижней губе; (☆) – зубы; ост. обозначения см. на рис. 1, 3. Масштаб: а – 400, б – 50, в – 600, г – 20 мкм.
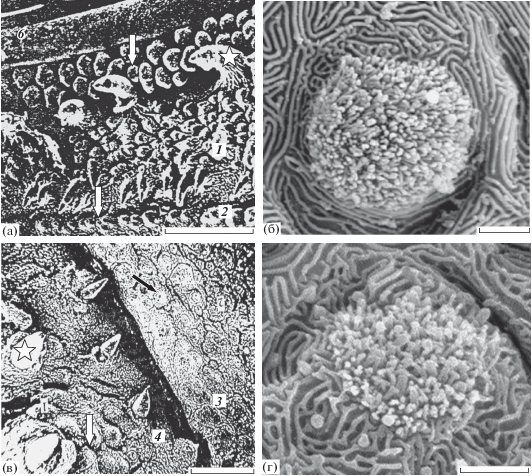
Верхняя губа несёт многочисленные разноразмерные вкусовые почки непогружённого типа. Эта зона обладает высокой плотностью распределения вкусовых почек (155 шт/мм2), большинство из них имеют крупные размеры сенсорного поля − до 140 (в среднем 135) мкм2 (рис. 2в, 2г), которое возвышается над поверхностью эпителия (рис. 5б, 5г).
Премаксиллярная зона, примыкающая к верхней губе, имеет щётку мелких и острых зубов (рис. 5а). Между зубами находятся высокие эпидермальные сосочки сферической формы с несколькими вкусовыми почками на вершине. Вкусовые сосочки расположены разрозненно. Средние размеры сенсорного поля (102 мкм2) меньше, чем на верхней губе. На сенсорном поле вкусовых почек преобладают тонкие микровиллы поливиллярных клеток (рис. 5б, 5г). Однако плотность распределения вкусовых почек (95 шт/мм2) в этой зоне значительно ниже, чем в зоне верхней губы.
Максиллярная зона расположена каудальнее премаксиллярной. Она не имеет зубов. Вкусовые почки находятся на вершине небольших эпидермальных сосочков (рис. 5а). Вкусовые почки этой зоны подобны почкам премаксиллярной зоны и также характеризуются сравнительно крупным сенсорным полем (118 мкм2). Плотность распределения вкусовых почек сильно варьирует в разных участках этой зоны − 40−70 (60) шт/мм2.
Твёрдое нёбо представляет вкусовую зону в центральной части свода верхней челюсти (рис. 6). Она образована мягкими складками эпителия с редкими одиночными вкусовыми почками непогружённого типа (рис. 6а). Эта зона отличается сравнительно низкой плотностью распределения (10–30 (24) шт/мм2) крупных вкусовых почек (115 мкм2). Морфология сенсорного поля вкусовых почек твёрдого нёба сходна с почками на мягком нёбе и на языке (рис. 6в, 6г). Микровиллы рецепторных клеток на поверхности сенсорного поля вкусовых почек одного размера по высоте (0.70–0.72 мкм), но различаются по диаметру. Диаметр толстых микровилл составляет 0.18 мкм, тонких нитевидных − 0.11 мкм.
Рис. 6.
Нёбные зоны трески Gadus morhua marisalbi: а – поверхность твёрдого нёба, б – сосочек мягкого нёба с вкусовыми почками, в – вкусовая почка на мягком нёбе, г – вкусовая почка на твёрдом нёбе; обозначения см. на рис. 1, 3. Масштаб: а – 500, б – 100, в – 20, г – 50 мкм.
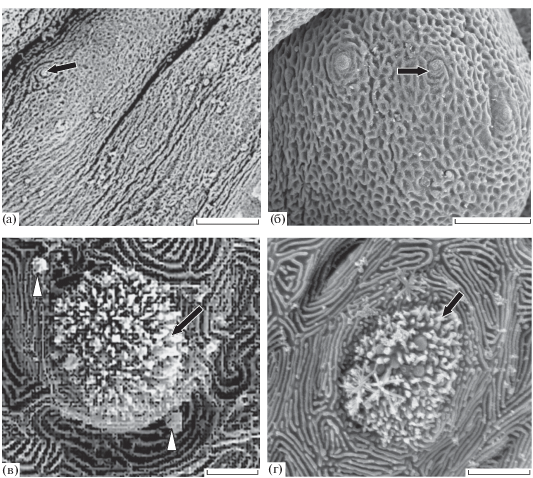
Мягкое нёбо подобно таковому карпа, представляет собой парное скопление крупных, плотно упакованных вкусовых сосочков сферической формы, несущих крупные вкусовые почки. Площадь их сенсорного поля составляет 103 мкм2, плотность распределения − 250−300 (289) шт/мкм2 (рис. 2в, 2г). Сенсорное поле большинства вкусовых почек мягкого нёба образуют тонкие микровиллы (0.11 мкм в диаметре), длина которых достигает 0.9–1.0 мкм. В этой зоне присутствуют и вкусовые почки, сенсорное поле которых несёт как тонкие, так и толстые микровиллы (рис. 6в).
Вентральный свод ротовой полости включает вкусовую зону нижней губы, дентальную зону и язык. Нижняя губа имеет гладкий эпителий с большим количеством погружённых вкусовых почек и кратеров секреторных элементов. Плотность распределения вкусовых почек на нижней губе (168 шт/мм2) несколько выше, чем на верхней (рис. 2г). Здесь, как и на верхней губе, преобладают особенно крупные вкусовые почки с площадью сенсорного поля 100 мкм2.и более, в среднем 110 мкм2 (рис. 2в).
Дентальная зона отделена от губы глубокой бороздой и образована эпителием, покрывающим dentale. Она несёт многочисленые мелкие зубы, подобно премаксиллярной зоне (рис. 5в). Между зубами, направленными назад, возвышаются на эпидермальных папиллах сравнительно мелкие вкусовые почки (98 мкм2); плотность их распределения в этой зоне (71 шт/мм2) значительно ниже, чем на нижней губе (рис. 2г).
Язык имеет вкусовые папиллы округлой формы, каждая из которых несёт одну вкусовую почку непогружённого типа. Площадь сенсорного поля вкусовых почек на языке (105 мкм2) сходна с площадью вкусовых почек на нёбе (рис. 2в). Сенсорное поле вкусовых почек по своей морфологии сходно с сенсорным полем вкусовых почек в других зонах ротовой полости. Средняя плотность распределения вкусовых почек на языке (25 шт/мм2), как и на нёбе, значительно ниже, чем в других зонах (рис. 2г). В эпителии языка и нёба содержится большое количество вершин одиночных сенсорных клеток и кратеров слизистых секреторных клеток.
Морфология сенсорного поля вкусовых почек трески во всех сенсорных зонах ротовой полости сходна и видоспецифична. Сенсорное поле образовано преимущественно микровиллами поливиллярных клеток, диаметр которых варьирует от 0.11 до 0.18 мкм, а длина не превышает 0.9–1.0 мкм.
ОБСУЖДЕНИЕ
Карп и треска обладают хорошо развитыми системами зрительной, сейсмосенсорной и химической ориентации, что обеспечивает им надёжную полисенсорную основу для поиска и выбора пищевых объектов. Их вкусовые предпочтения характеризуются высокой видовой специфичностью (Касумян, 1997; Касумян и др., 2009). Морфология и структурная организация вкусового аппарата ротовой полости у взрослых особей карпа и трески различны и соответствуют характеру их питания и пищевого поведения (Western, 1969; Kawakita et al., 1978; Певзнер, 1985; Jakubowski, Whitear, 1990; Марусов, 1997; Девицина, Головкина, 2007, 2008). В то же время молодь этих экологически различных видов имеет сходный характер питания и относится к одной категории полисенсорных бентофагов. Основу их питания составляют личинки насекомых и черви (Суворов, 1948; Никольский, 1971). Проведённое исследование показало сходство структурной организации вкусового рецепторного аппарата в ротовой полости у молоди карпа и беломорской трески. Сходство это проявляется в распределении вкусовых рецепторов в ротовой полости у молоди обоих видов. Особенно хорошо это выражено на внутренней поверхности губ и в прилегающих к ним зонах челюстной дуги (дентальной, премаксиллярной и максиллярной). Отметим, что на поверхности губ у молоди обоих видов, особенно у карпа, наряду с вкусовыми почками присутствуют многочисленные одиночные поливиллярные хемосенсорные клетки, относящиеся к системе общей химической чувствительности (Whitear, 1971; Appelbaum, Riehal, 1997; Девицина, Головкина, 2007, 2008). Эти зоны, расположенные на входе в ротовую полость, образуют ростральный отдел ротовой полости и играют важную роль в первичной оценке пищевого объекта (Касумян, Пономарёв, 1990; Касумян, 1997). Наружная поверхность губ у молоди карповых рыб (карп и карась Carassius auratus) обладает высокой тактильной чувствительностью (Девицина, Лапшин, 2016). У молоди беломорской трески также выявлены морфологические признаки существования тактильной рецепции в околоротовых зонах (Девицина, 1997). Можно полагать, что у молоди обоих исследованных видов рыб эпителий губ, премаксиллярной и максиллярной зон обеспечивает комплексное хемосенсорное и тактильное тестирование пищевых объектов на входе в ротовую полость.
Сходство структурной организации вкусового аппарата ротовой полости у молоди карпа и трески выражается и в том, что они могут использовать всасывающий механизм при захвате пищи (Суворов, 1948; Casinos, 1978). Всасывающий механизм захвата пищевых объектов направляет поток воды с кормом на систему крупных вкусовых сосочков, несущих вкусовые почки, в максиллярной и дентальной зонах ротовой полости. Эти зоны характеризуются высокой плотностью распределения вкусовых почек. Физиологические исследования вкусовой рецепции ротовой полости позвоночных животных показали, что в плотных скоплениях вкусовые почки в процессе качественной оценки пищевого объекта функционально взаимодействуют (Есаков, 1977). Интеграция афферентных сигналов во вкусовой системе играет важную роль при формировании поведенческих реакций.
Значительное сходство структурно-функциональной организации вкусового аппарата у молоди данных видов также выражается в организации сенсорных зон центрального отдела ротовой полости. У молоди как карпа, так и трески этот отдел имеет наиболее простую системную организацию: в сенсорных зонах нёба и языка нет крупных вкусовых сосочков и сравнительно низкая концентрация одиночных вкусовых почек. Очевидно, что при таком распределении отсутствует возможность взаимодействия вкусовых почек при их активации, в отличие от зон рострального отдела. Под контролем рецепторов языка и нёба нужные элементы корма перемещаются в каудальную часть ротовой полости. Комплементарное сходство структурных параметров вкусового аппарата языка с вкусовым аппаратом твёрдого нёба, отмеченное у молоди обоих видов, вероятно, имеет и сходную функциональную обусловленность (Sibbing, Uribe, 1985).
Поведенческие эксперименты показали, что для многих видов рыб характерно многократное оросенсорное тестирование даже одного пищевого объекта с многократными схватыванием и отверганием (Павлов, Касумян, 1990; Касумян, Морси, 1996; Михайлова, Касумян, 2016). Такой характер тестирования, основанный на морфофункциональном взаимодействии рострального и центрального отделов ротовой полости, выявлен у рыб-бентофагов, но отсутствует у хищников (Суворов, 1948; Марусов, 1997; Девицина, Головкина, 2011, 2018).
Значительное сходство молоди исследуемых видов отражается и в структуре каудального отдела ротовой полости в зоне мягкого нёба: плотность распределения вкусовых почек в этой сенсорной зоне значительно выше, чем в других зонах ротовой полости. Пищевой объект в этой зоне получает повреждение щётками мелких зубов у трески и глоточными зубами у карпа, что усиливает эффективность действия химического стимула на выходе из ротовой полости. Рецепторы мягкого нёба посылают второй после челюстной дуги мощный сигнал во вкусовые центры мозга. Таким образом, можно полагать, что у молоди данных видов существует морфофункциональное сходство одноимённых сенсорных зон ротовой полости. Это является отражением сходства молоди по характеру оросенсорного тестирования пищевых объектов.
Карп и треска обладают сложной организацией мозговых центров вкусовой системы, которые получают топические проекции отделов ротовой полости (Finger, 1981; Kanwal, Finger, 1992; Reutter, Witt, 1993). Специфические особенности в морфологии вкусового рецепторного аппарата и соответствующих центров мозга у молоди исследованных видов составляют основу организации их пищевого поведения (Brawn, 1969; Шелихов, Наумова, 1976; Marui, 1977).
Результаты настоящей работы показали, что молодь карпа и трески имеет сходную структурную организацию вкусового аппарата в ротовой полости. Сходство молоди этих видов выражается в однотипном дискретном распределении вкусовых почек, скопления которых формируют три отдела ротовой полости. Особенности количественного распределения вкусовых рецепторов показывают однотипное распределение функциональной нагрузки по сенсорным зонам и отделам ротовой полости у одновозрастной молоди обоих исследованных видов. Важную роль в формировании вкусового аппарата ротовой полости играет сходный характер питания молоди этих видов (Никольский, 1980). Структурно-функциональная организация сенсорных зон в ротовой полости у молоди исследованных видов представляет собой сложную адаптивную систему вкусового анализа. Сходство молоди разных видов по структурной организации вкусового аппарата можно рассматривать как проявление параллельной (адаптивной) изменчивости.
Список литературы
Андрияшев А.П. 1954. Рыбы северных морей СССР. М.: Изд-во АН СССР, 566 с.
Девицина Г.В. 1990. Исследование морфологии хемосенсорных систем // Сенсорная физиология морских рыб (методические аспекты). Мурманск: Апатиты. С. 7–10.
Девицина Г.В. 1997. К вопросу о хемосенсорно-тактильном обеспечении пищевого поведения тресковых рыб // Вопр. ихтиологии. Т. 37. № 1. С. 94–100.
Девицина Г.В., Головкина Т.В. 2007. Особенности морфологии вкусовых рецепторов карпа, Cyprinus carpio L. в связи с его характером питания // Матер. Междунар. симп. “Тепловодная аквакультура и биологическая продуктивность водоёмов аридного климата”. Астрахань: Изд-во АГТУ. С. 399–405.
Девицина Г.В., Головкина Т.В. 2008. Морфология вкусового аппарата ротовой полости наваги Eleginus navaga L. и трески Gadus morhua marisalbi Der. (Gadidae, Teleostei) // Сенсорные системы. Т. 22. № 1. С. 70–86.
Девицина Г.В., Головкина Т.В. 2011. Вкусовой аппарат ротовой полости молоди радужной форели Oncorhynchus (=Parasalmo) mykiss // Вопр. ихтиологии. Т. 51. № 1. С. 113–123.
Девицина Г.В., Головкина Т.В. 2018. Структурная организация вкусового аппарата у харациновых рыб (Characidae, Teleostei) // Там же. Т. 58. № 1. С. 53–66. https://doi.org/10.7868/S0042875218010071
Девицина Г.В., Лапшин Д.Н. 2016. Тактильная рецепция наружных покровов головы серебряного карася (Carassius auratus gibelio) // ДАН. Т. 466. № 4. С. 493–495. https://doi.org/10.7868/S0869565216040253
Есаков А.И. 1977. О периферических механизмах регуляции деятельности вкусового рецепторного аппарата // Сенсорные системы: морфофизиологические и поведенческие аспекты / Под ред. Гершуни Г.В. Л.: Наука. С. 60–75.
Касумян А.О. 1997. Вкусовая рецепция и пищевое поведение рыб // Вопр. ихтиологии. Т. 37. № 1. С. 78–93.
Касумян А.О., Морси А.М.Х. 1996. Вкусовая чувствительность карпа к свободным аминокислотам и классическим вкусовым веществам // Там же. Т. 36. № 3. С. 386–399.
Касумян А.О., Пономарёв В.Ю. 1990. Формирование пищевой поисковой реакции на естественные химические сигналы в онтогенезе карповых рыб // Там же. Т. 30. Вып. 3. С. 447–456.
Касумян А.О., Марусов Е.А., Сидоров С.С. 2009. Влияние пищевого запахового фона на вкусовые предпочтения и пищевое поведение карпа Cyprinus carpio и трески Gadus morhua // Там же. Т. 49. № 4. С. 528–540.
Марусов Е.А. 1997. Особенности реагирования беломорской трески Gadus morhua marisalbi и молоди беломорской кумжи Salmo trutta на корм и пищевые химические стимулы // Там же. Т. 37. № 1. С. 138–142.
Михайлова Е.С., Касумян А.О. 2016. Оросенсорное тестирование пищи у рыб: хронология поведения // Изв. РАН. Сер. биол. № 4. С. 377–388. https://doi.org/10.7868/S000233291604010X
Никольский Г.В. 1971. Частная ихтиология. М.: Высш. шк., 470 с.
Никольский Г.В. 1980. Структура вида и закономерности изменчивости рыб. М.: Пищ. пром-сть, 184 с.
Павлов Д.С., Касумян А.О. 1990. Сенсорные основы пищевого поведения рыб // Вопр. ихтиологии. Т. 3. Вып. 5. С. 720−732.
Певзнер Р.А. 1985. Роль вкусовой сенсорной системы в пищевом поведении рыб // Сигнализация и поведение рыб. Мурманск: Апатиты. С. 63–72.
Суворов Е.К. 1948. Основы ихтиологии. М.: Сов. наука, 580 с.
Шелихов В.Н., Наумова Т.С. 1976. Системный подход к анализу функции полости рта и проблеме переработки сенсорной информации // Афферентная функция полости рта и проблема переработки сенсорной информации / Под ред. Шелихова В.Н. М.: Минздрав РСФСР. С. 5–18.
Appelbaum S., Riehal R. 1997. Scanning electron microscopic observations of the chemo – and mechanoreceptors of carp larvae (Cyprinus carpio) and their relationship to early behavior // Aquat. Liv. Resour. V. 10. P. 1–12.https://doi.org/10.1051/alr:1997001
Atema J. 1971. Structures and functions of the sense of taste in the catfish (Ictalurus natalis) // Brain Bihav. Evol. V. 4. P. 273–294.
Brawn V.M. 1969. Feeding behaviour of cod (Gadus morhua) // J. Fish. Res. Board Can. V. 26. № 3. P. 316–318.
Casinos A. 1978. The comparative feeding mechanism of Gadidae and Macrouridae // Gegenbaurs Morphol. Jahrb. V. 124. № 3. P. 434–449.
Devitsina G.V. 2005. Comparative morphology of intraoral taste apparatus in fish // J. Ichthyology. V. 45. Suppl. 2. P. S286–S306.
Devitsina G.V., Chervova L.S. 1994. Morphological and physiological aspects of fish chemoreception // Adv. Fish Biol. Fish. V. 1. P. 185–191.
Devitsina G.V., Golovkina T.V., Rod’kin M.M. 2011. Feature of gustatory system morphology in early juveniles of Siberian sturgeon Acipenser baerii (Acipenseridae Acipenseriformes) // J. Ichthyol. V. 51. № 11. P. 1104–1116. https://doi.org/10.1134/S0032945211110038
Finger T.E. 1981. Laminar and columnar organization of the vagal lobe in goldfish: possible neural substrate for sorting food from gravel // Soc. Neurosci. Abstr. V. 7. P. 665.
Finger T.E., Morita Y. 1990. Two gustatory systems: facial and vagal gustatory nuclei have different brainstem connections // Science. V. 227. P. 776–778.
Jakubowski M., Whitear M. 1990. Comparative morphology and cytology of taste buds in teleosts // Z. Mikrosk.-Anat. Forsch. V. 104. P. 529–560.
Kanwal J.S., Finger T.E. 1992. Central representation and projections of gustatory systems // Fish Chemoreception. V. 6. L.: Chapman and Hall. P. 79–102.
Kasumyan A.O., Døving K. 2003. Taste preference in fish // Fish Fisheries. V. 4. P. 289–347.
Kawakita K., Marui T., Funakoshi M. 1978. Scanning electron microscope observation on the taste buds of the carp (Cyprinus carpio L.) // Jpn. J. Oral. Biol. V. 20. P. 103–113.
Marui T. 1977. Taste responses in the facial lobe of the carp, Cyprinus carpio // Brain. Res. V. 217. P. 59–68.
Reutter K., Witt M. 1993. Morphology of vertebrate taste organs and their nerve supply // Mechanisms of taste transduction / Eds. Simon S.A., Roper S.D. Boca Raton: CRC Press. P. 29–82.
Sibbing F.A., Uribe R. 1985. Regional specialization in the oro-pharyngeal wall and food processing in the carp (Cyprinus carpio L.) // Netherl. J. Zool. V. 35. P. 377–422.
Western J.R.H. 1969. Studies on the diet, feeding mechanism and alimentary tract in two closely related teleosts, the freshwater Cottus gobio L. and the marine Parenophrys bubalis // Acta Zool. V. 50. P. 185–205.
Whitear M. 1971. Cell specialization and sensory function in fish epidermis // J. Zool. V. 163. P. 237–264.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии