

Вопросы ихтиологии, 2020, T. 60, № 3, стр. 261-270
Обнаружение Photonectes uncinatus на Китовом хребте (Юго-Восточная Атлантика) и переописание вида (Melanostomiidae)
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии им. П.П. Ширшова РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 30.07.2019
После доработки 08.08.2019
Принята к публикации 08.08.2019
Аннотация
В сборах меланостомовых рыб с Китового хребта, выполненных И.А. Труновым в 1976 г., обнаружен второй экземпляр редкого вида Photonectes uncinatus, ранее известного только по голотипу с возвышенности Риу-Гранди (Юго-Западная Атлантика). Приведено переописание вида с учётом новых данных, уточнена индивидуальная изменчивость ряда диагностически значимых морфологических признаков. Установлено, что особенности расположения зубов на palatinum имеют диагностическое значение при выделении групп видов внутри рода Photonectes. На основании литературных данных P. uncinatus указывается также для вод Новой Зеландии (плато Баунти). Обсуждается тип ареала рассматриваемого вида.
В составе рода Photonectes Günther, 1887 в настоящее время признается 28 валидных видов (Прокофьев, 2019; Koeda, Ho, 2019; Prokofiev, Klepadlo, 2019), некоторые из которых известны науке по единичным экземплярам. Одним из таких редких видов является P. uncinatus Prokofiev, 2015, описанный по единственному экземпляру из вод Юго-Западной Атлантики. При первоописании (Прокофьев, 2015) он был отнесен к подроду Trachinostomias Parr, 1927 на основании наличия у него толстой кожи на вертикальных плавниках. После переоценки признаков, положенных в основу внутриродовой классификации Photonectes (Прокофьев, 2019), P. uncinatus был перемещен в состав видовой группы “brauer” номинотипического подрода, включающей помимо рассматриваемого вида также P. braueri (Zugmayer, 1913), P. dinema Regan et Trewavas, 1930, P. distichodon Prokofiev, 2019 и P. leucospilus Regan et Trewavas, 1930. Из этих видов только P. braueri известен по нескольким десяткам поимок, опубликованных в литературе, прочие виды исключительно редки в коллекциях, а P. dinema и P. leucospilus до сих пор известны только по молоди.
Поэтому представляет большой интерес нахождение нового экземпляра P. uncinatus, на этот раз в водах Юго-Восточной Атлантики (Китовый хребет), изучение которого позволяет существенно расширить пределы изменчивости некоторых признаков и уточнить диагностическую значимость ряда из них. Кроме того, голотип рассматриваемого вида является нерестовой самкой, тогда как новый экземпляр – зрелым самцом, что позволяет охарактеризовать у этого вида половой диморфизм в строении посторбитального органа, который у меланостомовых рыб может быть весьма значительным (Бородулина, 2009), в то время как особенности строения посторбитального органа являются наиболее значимыми диагностическими признаками P. uncinatus. Помимо этого анализ литературных данных позволил установить, что P. uncinatus также уже был однажды выловлен в юго-западной части Тихого океана, поскольку из новозеландских вод был указан (с кратким, но достаточно содержательным морфологическим описанием и фотографией рыбы) экземпляр Photonectes, оставленный без видового определения (Aizawa, 1990). Настоящая статья посвящена переописанию данного вида с учетом полученных новых сведений о нём.
МАТЕРИАЛ И МЕТОДИКА
Изученный материал по P. uncinatus приведён при видовом описании, перечень сравнительного материала по другим видам рода указан в предыдущих публикациях (Прокофьев, 2015, 2019; Prokofiev, Klepadlo, 2019). Методика изучения и терминология соответствуют Гиббсу (Gibbs, 1968) с дополнениями Прокофьева (2019). В видовом описании первыми приведены значения счётных и пластических признаков голотипа, за ними в скобках – отличающиеся значения у экземпляра с Китового хребта. В случае отличий в счётных значениях одних и тех же признаков на разных сторонах тела одной рыбы показатели разделены знаком “/”. Морфометрия в первоописании P. uncinatus была выполнена по голотипу, переведенному из 4–10%-ного формалина в 70–75%-ный этанол непосредственно перед исследованием. Измерения голотипа, выполненные после пяти лет хранения в спирте, показали изменения некоторых пропорций, в настоящей статье все пропорции указаны по текущему состоянию экземпляра. Измерения второго экземпляра выполнены через полгода после его перевода из формалина в спирт. В работе использованы следующие сокращения: D, A, P, V – соответственно спинной, анальный, грудные и брюшные плавники; vert. – общее число позвонков, v.c. – число хвостовых позвонков (т.е. с замкнутыми гемальными дугами); БМРТ – большой морозильный рыболовный траулер, НИС – научно-исследовательское судно, ст. – океанографическая станция; ЗММУ – Зоологический музей Московского государственного университета; ИО РАН – Институт океанологии РАН, Москва; MNHN – Национальный музей естественной истории, Париж (Франция). Используется стандартная номенклатура фотофоров стомиеобразных рыб (Morrow, Gibbs, 1964).
РЕЗУЛЬТАТЫ
(рис. 1–4)
Рис. 1.
Photonectes uncinatus, общий вид: а – голотип SL 250 мм, самка, возвышенность Риу-Гранди; б – 1 экз., самец SL 222 мм, Китовый хребет.

Рис. 2.
Photonectes uncinatus, детали строения (по голотипу): а – фотофоры IP-серии (1–10 – порядковые номера органов); б, в – скопления мелких поверхностных пятен белой, вероятно, светящееся ткани на дорсальной поверхности тела (б – непосредственно за головой, в – посредине туловища); г – пятна светящейся ткани и вторичные фотофоры на боках средней части туловища. M – миосепта, P – лучи грудного плавника; PV-1 – первый фотофор серии PV; sph – вторичный фотофор; tp – последняя пора темпорального сенсорного канала; wl – пятна белой, вероятно, светящейся ткани. Масштаб: а–в – 3, г – 4 мм.

Рис. 3.
Photonectes uncinatus, окологлазничные светящиеся железы и озубление челюстей: а, е – голотип, левая сторона тела; б, в, ж – экземпляр от Китового хребта, левая сторона тела; г, д – экземпляр от Китового хребта, правая сторона тела, зеркальное отражение. Красными стрелками показаны премаксиллярные зубы наружного ряда, белыми – передненижний выступ окаймления посторбитального органа (б, в) и складка ткани, регулирующая свечение посторбитального органа (г). so – суборбитальный орган; cr – кольца коллагеновой ткани в основаниях зубов. Масштаб: 5 мм (линейка общая для е и ж).
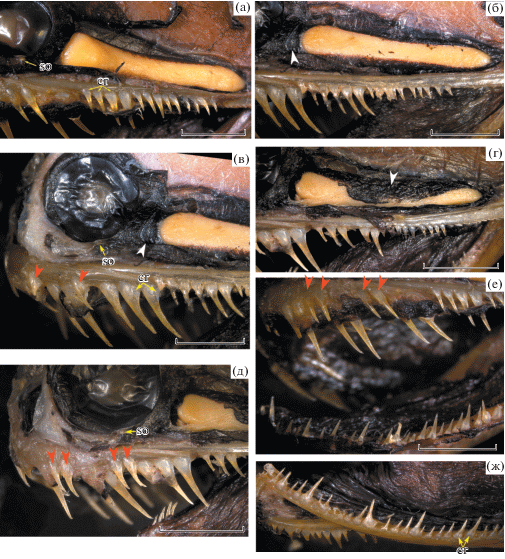
Рис. 4.
Photonectes uncinatus, детали строения, по голотипу (б–д) и экземпляру с Китового хребта (а, е, ж): а – зубы на языке, б – пятковидные выступы retroarticulare, в – сейсмосенсорная система (схема); г – поперечные гребешки, несущие ряды свободных невромастов, на дорсальной поверхности головы; д, е – подбородочный усик (красной стрелкой показан BR-1); ж – грудной плавник. c.io – подглазничный сенсорный канал, c.so-t – супраорбитально-темпоральный сенсорный канал, fcr – костные гребни frontalia; N1, N2 – соответственно передняя и задняя ноздря, so – суборбитальный орган. Масштаб: а, ж – 5; б, д – 4; в – 3; г, е – 2 мм.

Photonectes sp.: Aizawa, 1990. P. 122 (краткое описание, фотография, плато Баунти).
Photonectes (Trachinostomias) uncinatus: Прокофьев, 2015. С. 136. Рис. 4 (первоописание, возвышенность Риу-Гранди).
Photonectes uncinatus: Прокофьев, 2019. С. 379, 386, 402. Рис. 1у, 2д (изменение подродовой принадлежности, определительная таблица, по голотипу).
Материал. Голотип: ЗММУ № 23280 (из ИО РАН, № 2), SL 250 мм, самка со зрелой икрой (рис. 1а), НИС “Ихтиандр”, рейс 2188.XIII, трал 62, 20 февраля 1989 г., возвышенность Риу-Гранди, глубина 1070 м. Нетиповой экземпляр: ИО РАН, SL 222 мм, самец со зрелыми гонадами (рис. 1б), БМРТ “Салехард”, август–сентябрь 1976 г., Китовый хребет, ~30°–33° ю.ш., ~2°–3° з.д., из тралов № 187–195, сборы И.А. Трунова.
Описание (рис. 2–4). D 16 (18), A 19 (20), P 2, V 7 (8); vert. 54 (53), v.c. 16. Морфометрические показатели приведены в таблице. Фотофоров в сериях: BR 10/11 (10), IP 8 + 2, PV 27 (26), OV 22 (21), VAL 12/14 (13), VAV 14 (16), AC 11 (9). Все органы BR одинаковой величины, расположены на равном расстоянии друг от друга (но у голотипа с левой стороны первые два органа сближены, а у экземпляра с Китового хребта слева BR-2 смещён вверх и также сближен с BR-3). В серии IP первые восемь органов уменьшены в размере (в 1.5 раза мельче двух задних органов и органов серии PV), первые семь из них отстоят друг от друга на величину своего диаметра, восьмой фотофор отстоит от седьмого на промежуток, соответствующий 1.5 его диаметра, а от девятого – на промежуток, равный трём своим диаметрам; девятый и десятый фотофоры IP и первый фотофор PV отстоят друг от друга на равные промежутки (1.5 диаметра органа) (рис. 2а). Расстояние от вершины истмуса до первой пары органов IP слегка превышает суммарную длину восьми передних фотофоров IP и промежутков между ними. Органы PV и VAV образуют прямую линию; 1/2 (1) органа VAL и 2 (3) органа VAV расположены над началом А. У голотипа первый фотофор АС отстоит от последнего фотофора VAV на величину промежутка между последним и предпоследним органами VAV, а у экземпляра с Китового хребта – на два таких промежутка; у голотипа передние органы AC расположены чуть ближе к линии органов VAL, чем VAV, тогда как у экземпляра с Китового хребта – лишь немногим выше линии органов VAV; три последних фотофора АС расположены на хвостовом стебле. На голове и теле рассеяны мелкие вторичные фотофоры, особенно густо сконцентрированные вдоль срединнобрюшной линии и в полосе примерно посредине промежутка между органами OA и PV + VAV. Очень мелкие фотофоры сидят по краю глазного яблока. У голотипа сохранилась кожа межчелюстной перемычки непосредственно за симфизом нижней челюсти, густо покрытая мелкими вторичными фотофорами (далее назад кожа сорвана). Помимо вторичных фотофоров кожа спины на участке непосредственно позади головы (рис. 2б), на верхнебоковой поверхности туловища между миосептами (рис. 2в) и на брюшной поверхности несёт скопления мелких, не контрастных, но чётко отграниченных светлых пятнышек, оконтуренных тёмным пигментом, вероятно, также светящихся (на боках туловища эти пятнышки распределены более рассеянно, чем на дорсальной поверхности: рис. 2г). Контрастных пятен белой светящейся ткани на голове, рыле или над жаберным отверстием нет (у голотипа кожа здесь сохранилась достаточно хорошо). Голубая светящаяся ткань отсутствует. Анус окружен кольцом интенсивно-чёрной ткани (перипрокт), расположен у начала А.
Морфометрические признаки Photonectes uncinatus
| Признак | Риу-Гранди (голотип) | Китовый хребет |
|---|---|---|
| SL, мм | 250 | 222 |
| В % SL | ||
| Длина головы | 12.0 | 12.2 |
| Длина рыла | 2.0 | 1.6 |
| Горизонтальный диаметр глаза | 2.4 | 3.2 |
| Ширина костного межглазничного промежутка | 4.0 | 4.5 |
| Длина посторбитального органа | 6.0 | 6.1 |
| Длина подбородочного усика | 1.6 | 1.0 |
| Длина верхней челюсти | 12.0 | 12.2 |
| Длина Р | 4.0 | 7.7 |
| Длина V | 9.6 | 12.2 |
| Расстояние: | ||
| ̶ антедорсальное | 82.0 | 83.3 |
| ̶ антевентральное | 64.8 | 60.8 |
| ̶ антеанальное | 82.0 | 82.4 |
| ̶ пектовентральное | 44.0 | 51.4 |
| ̶ вентроанальное | 17.6 | 20.3 |
| Максимальная высота тела | 13.6 | 13.5 |
| Минимальная высота тела | 2.0 | 1.8 |
| Длина хвостового стебля | 5.2 | 5.9 |
Суборбитальный фотофор небольшой, одинаково развитый у обоих экземпляров. Посторбитальный орган очень длинный, в 2.5 (2.0) раза превышает горизонтальный диаметр глаза, составляет 50% длины верхней челюсти, его фотогенная железа несколько сужена в средней части (рис. 3а–3г). Высота переднего края, середины и заднего конца фотогенной железы соответственно в 5.0 (5.4), 10.0 (13.5) и 7.5 (8.8) раза меньше её длины. Передненижний край интенсивно-чёрной ткани, окаймляющей фотогенную железу, образует короткий закруглённый выступ, не заходящий за вертикаль заднего края глаза (рис. 3б, 3в). Фотогенная железа посторбитального органа может (частично?) закрываться (сверху вниз) складкой чёрной ткани: экземпляр с Китового хребта зафиксирован с частично закрытой железой с правой стороны (рис. 3г). Половой диморфизм в размерах и форме посторбитального органа не выражен.
Озубление praemaxillare гетерогенное бисериальное (рис. 3в, 3д, 3е), число зубов во внутреннем ряду 7/8 (8/9), в наружном – 4 (2+2/2+2) (4 (2+2)/2 (1+1)). Во внутреннем ряду непосредственно за симфизом сидят 1/2 (2) мелких зуба с простыми вершинами, последующие зубы гораздо более длинные, менее различающиеся по величине, с отчётливыми зубчиками у вершин (barbed). У голотипа они сгруппированы попарно, а у экземпляра с Китового хребта как 1/2–2–3. У голотипа все внутренние премаксиллярные зубы подвижные, включая короткие присимфизные зубы (у экземпляра с Китового хребта они неподвижны). В основаниях всех челюстных зубов сильно развито кольцо коллагеновой ткани. Зубы наружного ряда praemaxillare у голотипа неподвижны, но кольцо коллагеновой ткани у них также хорошо развито; это позволяет предполагать, что их неподвижность может быть обусловлена условиями фиксации. У экземпляра с Китового хребта все зубы в наружном ряду подвижные, коллагеновое кольцо в их основаниях выражено столь же хорошо, как у других зубов во внутреннем ряду и на других челюстных костях. Наружные зубы со слабыми зубчиками у вершин (у голотипа с правой стороны они заметно более выражены). На maxillare 13/16 (14/16) резко различающихся по величине зубов (передние 4/5 (5/6) из них заметно длиннее последующих), за которыми следует гребёнка из 15(16) мелких зубов, сильно наклонённых и обращённых вершинами в каудальном направлении (рис. 3а, 3б, 3г). Озубление dentale гетерогенное, зубы не слишком резко различаются по величине, число зубов 32/33 или 34 (~37/~38) (рис. 3е, 3ж). Вершины максиллярных и большей части нижнечелюстных зубов с более или менее выраженными дополнительными зубчиками, в целом развитыми слабее, чем у премаксиллярных зубов внутреннего ряда. На сошнике 3+3 (2+2) зуба, прогрессивно увеличивающихся от внутреннего к наружным, у самого наружного зуба (у голотипа – у двух более наружных) вершина слабо изогнута или с зачаточным зубчиком. На нёбных костях 3/3 (5/3) зуба, из них второй (второй и третий/второй) мелкие и скрыты в коже11; первый нёбный зуб гораздо длиннее последующих и заметно отстоит от них; вершины всех зубов простые. На языке две группы зубов (рис. 4а), в каждой из которых они организованы в два параллельных ряда, числом 3+3 + 3+3 (3+3 + 2+2); все зубы с простыми вершинами. Жаберные тычинки представлены острыми зубчиками. Жаберные лепестки на epibranchiale-1 длинные, в самой верхней части ceratobranchiale-1 вдвое, далее примерно до половины его длины приблизительно в 1.5 раза короче лепестков epibranchiale-1 и нижней половины ceratobranchiale-1; лепестки последующих жаберных дуг очень длинные, выступают наружу. Retroarticulare образует длинный пятковидный выступ, направленный назад (рис. 4б).
На дорсальной поверхности головы хорошо выражены крупные поры сейсмосенсорной системы, представляющие собой фонтанели в костях крыши черепа (каналы проходят внутри кости). Супраорбитально-темпоральный канал представлен 11 порами: медиальнее ноздрей расположены две поры, медиальнее орбиты – пять, причём первые две отделены от трёх последующих острым костным гребнем; позади орбиты открываются три темпоральные и одна супратемпоральная пора (рис. 4в). С каждой стороны на крыше черепа медиальнее орбиты имеется по два продольных костных гребня. Кожа верха головы между супраорбитально-темпоральными порами имеет поперечные эпидермальные гребешки, несущие очень мелкие свободные невромасты (рис. 4г). Инфраорбитальных пор две; первая открывается книзу и кзади от ноздрей, а вторая – вниз, непосредственно перед суборбитальным фотофором (рис. 4в). Края пор и ноздрей окаймлены чёрным. В dentale имеется шесть фонтанелей; сохранность материала не позволяет оценить строение предкрышечно-нижнечелюстного канала более детально.
Подбородочный усик очень маленький, с тупой вершиной, у голотипа загнутой, а у экземпляра с Китового хребта прямой. У экземпляра с Китового хребта (самец) подбородочный усик, несмотря на его несколько меньшие размеры, короче, чем у голотипа (самка), будучи прижатым назад, не заходит своим концом за BR-1 (у голотипа это отношение оценить нельзя из-за особенностей фиксации), покрыт пигментированной кожей, без луковицы и/или придатков (рис. 4д, 4е).
Кожа сильно подвижная (подкожный студенистый слой при жизни, по-видимому, был сильно развит), полностью (до кончиков) покрывает лучи D и A у обоих экземпляров. Лучи Р тонкие, гибкие, равной длины (рис. 4ж), сравнительно короткие (3.0 (1.6) раза в длине верхней челюсти). Концы лучей V далеко не достигают ануса. Кожа при жизни была, по-видимому, интенсивно-чёрная (в настоящее время там, где она сохранилась, цвет на большей площади выцвел). Ротожаберная полость чёрная.
ОБСУЖДЕНИЕ
Photonectes uncinatus принадлежит к группе видов “braueri” номинотипического подрода, характеризующейся гетерогенным озублением челюстей и бисериальным озублением praemaxillare, наличием двух слабых равновеликих лучей Р, вертикальными плавниками, сплошь покрытыми чёрной кожей у половозрелых рыб и редукцией подбородочного усика на поздних стадиях онтогенеза (Прокофьев, 2019). От всех видов группы он хорошо отличается очень длинным посторбитальным органом (5.8–6.1 против 3.3–3.6% SL у обоих полов)22. В отличие от P. distichodon, также, по-видимому, связанного с южносубтропическими водами, подбородочный усик у половозрелых особей P. uncinatus лишён терминальной луковицы и филамента. От P. dinema и P. leucospilus, известных только по молоди, рассматриваемый вид может быть легко отличён по отсутствию пары контрастных пятен белой светящейся ткани у вершины рыла (Прокофьев, 2019). Выявленная изменчивость нивелирует ранее предполагавшиеся (Прокофьев, 2019) отличия P. uncinatus от P. braueri и P. distichodon по числу нёбных зубов. Передние один–два нёбных зуба у P. uncinatus отделены от последующих заметным промежутком (рис. 5а–5в), однако наличие такого промежутка, по-видимому, является специфической чертой группы видов “braueri” в целом, поскольку он обнаружен мной у большинства половозрелых экземпляров представителей этой группы (только у голотипа P. distichodon он не выражен) (рис. 5а–5и). У молоди P. braueri SL 43–50 мм нёбные зубы обычно неравновелики и неравномерно расставлены, но чёткое разделение на две группы выявлено только у одного из трёх изученных экземпляров (рис. 5е, 5ж). В пределах рода у видов подрода Trachinostomias подразделения зубов на две группы не наблюдается (рис. 5к–5м), а у видов из групп “caerulescens” и “albipennis” число нёбных зубов уменьшено и они сидят одной компактной группой (рис. 5н, 5о). Таким образом, расположение зубов palatinum в двух разобщённых группах является специфической особенностью видов группы “braueri”, но полностью сформировывается она, по-видимому, только на поздних стадиях онтогенеза.
Рис. 5.
Photonectes spp., варианты расположения зубов palatinum, схема (а, б, г, е–о – левая сторона тела; в, д – правая сторона): а – P. uncinatus, голотип SL 250 мм, Риу-Гранди; б, в – P. uncinatus SL 222 мм, Китовый хребет; г, д – P. braueri SL 225 мм, банка Сложная; е – P. braueri SL 50 мм, НИС “Петр Лебедев”, ст. IV-123A; ж – P. braueri SL 43 мм, НИС “Академик Курчатов”, ст. 835; з – P. distichodon, голотип SL 167 мм; и – P. distichodon, паратип SL 175 мм; к, л – P. margarita ЗММУ № 23286 (к – SL 200 мм, л – SL 70 мм); м – P. parvimanus SL 140 мм, НИС “М. Келдыш”, ст. 2332; н – P. caerulescens MNHN № 2003-1449 SL 140 мм; о – P. albipennis SL 170 мм, НИС “Витязь”, ст. 7400.
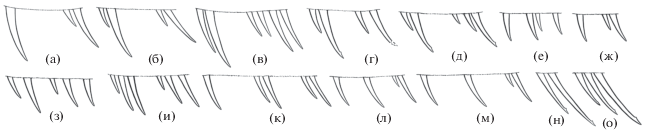
Новый экземпляр P. uncinatus с Китового хребта показывает существенную внутривидовую изменчивость по числу зубов на vomer, palatinum и в наружном ряду на praemaxillare, длине рыла и подбородочного усика, величине глаза, длине Р и числу фотофоров АС, но особенно неожиданным оказалось присутствие у него восьми лучей V. Число лучей V у видов Photonectes очень постоянно и равно семи (Morrow, Gibbs, 1964; собственные данные – см. список материала в: Прокофьев, 2019). Как очень редкая вариация, шесть лучей V приводится для 1 экз. P. dinema (Morrow, Gibbs, 1964) и 1 экз. P. margarita (Прокофьев, 2015); восемь лучей для видов данного рода до сих пор отмечено не было. Характер изменчивости по данному признаку нуждается в выяснении на более представительном материале. У голотипа имеется семь лучей V, такое же их число указано и для экземпляра с плато Баунти (Aizawa, 1990).
Половой диморфизм в размерах посторбитального органа у P. uncinatus не выражен – половозрелые самец и самка близких размеров имеют сходные значения его длины (таблица). Что же касается связи выявленных отличий между голотипом и экземпляром с Китового хребта с половым диморфизмом, то такая возможность требует подтверждения на более представительном материале – все отмеченные выше различия могут быть просто следствием индивидуальной изменчивости.
Голотип P. uncinatus был пойман в юго-западной части Атлантического океана на подводной возвышенности Риу-Гранди, новый экземпляр происходит с Китового хребта в Юго-Восточной Атлантике, однако данный вид, по-видимому, распространён циркумглобально в южносубтропических водах, потому что ему, несомненно, конспецифичен экземпляр SL 246 мм, пойманный в водах Новой Зеландии (плато Баунти, глубина 735 м) и указанный как Photonectes sp. (Aizawa, 1990). Для этого экземпляра приведено D 17, A 20, P 2, V 7, vert. 54, v.c. 14; BR 10, IP 8 + 2, PV 25, VAV 15, OV 21, VAL 15, AC 11; зубов на vomer 3+3, на palatinum 4, на praemaxillare 11 (расположение не описано), на maxillare 15; D и A, покрытые кожей; отсутствие пятен светящейся ткани на рыле, очень длинный посторбитальный орган и отсутствие подбородочного усика (Aizawa, 1990). Не исключено, что усик был просто пропущен из-за его очень малых размеров, хотя полностью отрицать возможность его полной редукции у отдельных экземпляров вида также нельзя.
Нахождения P. uncinatus в Южной Атлантике относятся к субтропическим водам, однако в Тихом океане он отмечен несколько южнее – к югу от южной субтропической конвергенции (ЮСК) (рис. 6). В понимании Гиббса (Gibbs, 1968), этот вид следует относить к особой ихтиофауне ЮСК (Subtropical Convergence species), хотя, как позднее отметили Гиббс и Маккинни (Gibbs, McKinney, 1988), трактовка ихтиофауны ЮСК довольно расплывчата из-за того, что ряд видов связан как с водами ЮСК, так и с водами преимущественно к северу от нее (semisubtropical species), либо к югу от нее (subantarctic species). В качестве полусубтропических видов Гиббс и Маккинни рассматривают виды, распространённые преимущественно между 30° и 40° ю. ш., и южноатлантические нахождения P. uncinatus относятся к субтропической водной массе. Однако новозеландское нахождение лежит даже несколько южнее границы ЮСК. Гиббс и Маккинни характеризуют специфическую ихтиофауну ЮСК видами, основной ареал которых лежит в пределах 35°–45° ю.ш. с отдельными нахождениями к северу и югу от этих границ; и в такой трактовке P. uncinatus может быть отнесён, с определённой долей условности связанной с единичностью находок, к ихтиофауне ЮСК. Для выяснения, насколько далеко этот вид проникает на север, конечно, необходимы дополнительные находки. Нужно однако отметить, что поимки рыб со зрелыми гонадами, готовыми к нересту, под 30°–33° ю.ш. свидетельствуют о том, что нерест, по крайней мере в Южной Атлантике, происходит в субтропических водах, то есть основа ареала включает данную водную массу и, таким образом, по крайней мере в Южной Атлантике P. uncinatus может представлять “полусубтропические виды” в понимании Гиббса и Маккинни (Gibbs, Mac-Kinney, 1988).

Список литературы
Бородулина О.Д. 2009. Внешнее строение посторбитального органа некоторых представителей семейства Melanostomiidae (Stomiiformes) // Вопр. ихтиологии. Т. 49. № 5. С. 717–720.
Прокофьев А.М. 2015. Материалы к ревизии подрода Trachinostomias (Melanostomiidae: Photonectes). Морфологическая изменчивость P. margarita с описанием двух новых видов // Там же. Т. 55. № 2. С. 131–138.
Прокофьев А.М. 2019. Материалы к ревизии рода Photonectes (Melanostomiidae) // Там же. Т. 59. № 4. С. 375–404.
Aizawa M. 1990. Photonectes sp. Fishes collected by the r/v Shinkai Maru around New Zealand / Eds. Amaoka K. et al. Tokyo: JAMARC. P. 122.
Ekman S. 1953. Zoogeography of the sea. Dublin: Sidgwick and Jackson, 417 p.
Gibbs R.H. 1968. Photonectes munificus, a new species of melanostomiatid fish from the south Pacific subtropical convergence, with remarks on the convergence fauna // Contrib. Sci. Los Angeles County Mus. № 149. P. 1–6.
Gibbs R.H., McKinney J.F. 1988. High-count species of the stomiid fish genus Astronesthes from the Southern Subtropical Convergence region: two new species and redescription of Cryptostomias (=Astronesthes) psychrolutes // Smiths. Contr. Zool. № 460. P. 1–25.
Koeda K., Ho H.-C. 2019. Photonectes banshee, a new species of the subgenus Photonectes (Stomiiformes: Stomiidae: Melanostomiinae) from Taiwan // Zootaxa. V. 4586. № 1. P. 180–186.
Morrow J.E., Gibbs R.H. 1964. Melanostomiatidae // Fishes of the Western North Atlantic. V. 1. Pt. 4. New Haven: Sears Found. P. 351–522.
Prokofiev A.M., Klepadlo C. 2019. Two new species of Photonectes with blue luminous tissue on body, and a re-examination of P. mirabilis (Teleostei: Stomiidae) // Zootaxa. V. 4590. № 2. P. 270–282.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии