

Известия РАН. Серия биологическая, 2019, № 2, стр. 169-180
Мускулатура покровов пяти представителей отряда Paramphistomatida (Plathelminthes, Trematoda)
Е. Н. Глаголева *, И. В. Ястребова, М. В. Ястребов
Ярославский государственный университет им. П.Г. Демидова
150057 Ярославль, пр-д Матросова, 9, Россия
* E-mail: glagolevaen@gmail.com
Поступила в редакцию 10.05.2017
После доработки 28.05.2018
Принята к публикации 30.01.2018
Аннотация
Описано строение покровов пяти видов трематод отряда Paramphistomatida, принадлежащих к четырем семействам. Установлено, что у всех видов толщина наружной пластинки тегумента минимальна в полостях присосок и постоянна на прочих участках покровов, а также совпадают число и последовательность мышечных слоев – кольцевой, продольный, диагональный. Отмечено, что слои состоят либо из отдельных мышечных волокон, либо из мышечных пучков, а к периферической мускулатуре могут быть ошибочно отнесены продольные паренхимные мышцы, топографически близкие к покровам. Обнаружено, что толщина и разреженность кольцевого и продольного слоев заметно варьируют на разных участках покровов, а для диагональных мышц характерны тупой угол пересечения, высокая регулярность расположения и незначительные изменения толщины.
Покровы трематод и других гельминтов – структура органной природы (Никишин, 2004). Их радикальное отличие от большинства прочих органов – огромная площадь поверхности, подвергающейся прямым воздействиям внешней среды. Естественно, что покровы участвуют в осуществлении многих жизненных отправлений паразитических червей. Среди отправлений важное место занимает двигательная активность, обеспечиваемая, в частности, периферической мускулатурой. Большая площадь поверхности покровов предполагает наличие региональных дифференцировок в их строении, связанных со специализацией отдельных участков, которые неодинаково взаимодействуют с внешней средой. Специализация проявляется в разной толщине наружной пластинки тегумента, в строении и топографии вооружения, если оно имеется, а также в различной архитектонике мускулатуры.
Сведения о пространственном распределении мышечных элементов в покровах трематод, к сожалению, фрагментарны (Ястребов, Ястребова, 2014). В пределах класса имеется ряд отрядов (Strigeidida, Notocotylida, Cyclocoelida, Hemiurida и др.), каждому из которых присущ специфический, легко узнаваемый габитус. Однако ни в одном из них мускулатура покровов не изучалась на представительном материале, позволяющем связать ее архитектонику с внешним строением и образом жизни гельминтов. В приведенный выше ряд можно с полным основанием включить отряд Paramphistomatida, который выделяется редким у трематод терминальным положением прикрепительных органов. Представителям данного таксона “повезло”, так как их присоски и копулятивные аппараты исследованы сравнительно хорошо, потому что строение названных органов лежит в основе системы отряда.
Цель работы – дополнение сведений о мышечной системе парамфистоматид описанием мускулатуры покровов.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве материала использованы мариты пяти видов парамфистоматид, относящихся к четырем семействам: Paramphistomum cervi (Zeder, 1790) и Calicophoron calicophorum (Fishoeder, 1901) (Paramphistоmatidae) от коровы Bos taurus, Stichorchis subtriquetrus (Rudolphi, 1814) (Cladorchidae) от бобра обыкновенного Castor fiber, Gastrodiscoides hominis (Lewis et McConnall) (Gastrodiscidaе) от кабана Sus scropha и Diplodiscus mehrai (Pande, 1937) (Diplodiscidae) от лягушки озерной Rana ridibunda. Для исследования отбирались одноразмерные половозрелые черви каждого вида от одной особи хозяина. Виды определены с помощью данных, опубликованных ранее (Скрябин, 1949; Jones et al., 2005). Микроморфология марит изучена под световым микроскопом МИКМЕД-1 (ОАО ЛОМО, Россия) по полным сериям фронтальных, сагиттальных и поперечных парафиновых срезов толщиной 7–10 мкм, изготовленных по стандартной методике и окрашенных методом Маллори. В совокупности число изученных серий срезов в сагиттальной проекции составило 13, в поперечной – 10, во фронтальной – 5.
Рисунки выполнены с помощью рисовального аппарата РА-4 (завод “Прогресс”, СССР), промеры – с помощью градуированной окулярной линейки. Весь цифровой материал в тексте и таблицах приведен в микрометрах. При измерении диаметров мышечных волокон и мышечных пучков первый диаметр (вертикальный) перпендикулярен базальной пластинке, вдоль которой идет волокно или пучок, второй (горизонтальный) параллелен ей. Если первый диаметр больше второго, форма поперечного сечения мышечного элемента именуется овальной, если наоборот – поперечно-овальной, при равенстве диаметров – округлой. Для диагональных мышц, которые не удается расположить строго перпендикулярно плоскости среза, указана толщина, измеренная в одной плоскости. В мышечных слоях расстояние между соседними мышцами приводится в относительных величинах, а именно в их горизонтальных диаметрах. По вертикальным диаметрам оценивается толщина слоя. Под расстоянием между диагональными мышцами подразумевается расстояние между теми из них, которые параллельны одна другой. Под углом пересечения мышц в этом слое понимается угол, ориентированный к концам тела, а не к его латеральным участкам.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
P. cervi – крупные трематоды с конусовидным, округлым в сечении телом, слегка согнутым на брюшную сторону. Ротовая присоска терминальная, с отчетливой предротовой губой. Брюшная присоска крупная, с вентральным устьем.
Как у всех прочих объектов, покровы данного вида не вооружены и состоят из следующих компонентов: тегумента, базальной пластинки и трех последовательно расположенных слоев мускулатуры – кольцевого, продольного, диагонального (рис. 1). Толщина наружной пластинки тегумента 8.4 мкм почти на всей поверхности тела. В устьях присосок она увеличивается и составляет 14–16.8, а в их полостях, напротив, уменьшается и равна соответственно 4.9–7.4 и 2.5–7.4 мкм. Толщина базальной пластинки составляет 5.6–7.4 и уменьшается до 2.8 мкм лишь в устьях присосок. Слой кольцевой мускулатуры состоит из одного ряда мышечных волокон. На участке тела между присосками они округлые в сечении диаметром 2.8–5.6, а на уровне присосок – в основном поперечно-овальные диаметром 2.8–5.6 × 5.6–8.4 мкм. Расстояние между соседними мышечными волокнами равно одному собственному диаметру на спинной стороне тела, а на брюшной стороне они расположены практически вплотную одно к другому.
Рис. 1.
Покровы Paramphistomum сervi на спинной стороне тела в его средней части. а – срединный сагиттальный срез, б – поперечный срез. 1 – наружная пластинка тегумента, 2 – базальная пластинка, 3 – кольцевые мышцы, 4 – продольные мышцы, 5 – диагональные мышцы, 6 – цитоны тегумента, 7 – продольные паренхимные мышцы, 8 – дорсовентральные мышцы, 9 – лакуны лимфатической системы; для рис. 1–5.
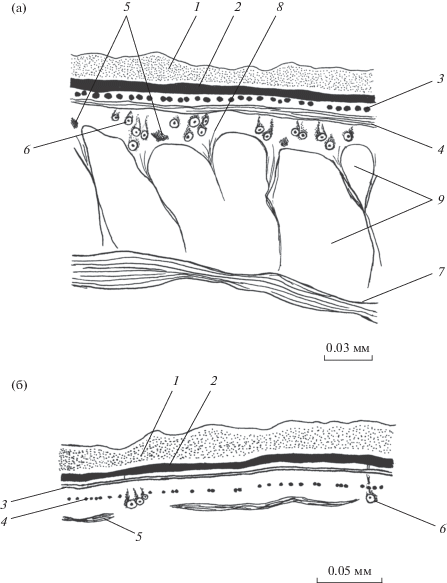
Мышцы продольного слоя по большей части поперечно-овальные в сечении. На уровне ротовой присоски их диаметр составляет на спинной и брюшной сторонах тела 3.7–12.3 × 4.9–17.2, на боковых сторонах – 3.7–7.4 × 4.9–12.3 мкм. Между присосками толщина слоя выравнивается на всей площади покровов, хотя диаметры отдельных волокон изменяются и составляют 4.9–9.8 × 4.9–24.6 на спинной и брюшной сторонах тела и 3.7–9.8 × 3.7–17.2 мкм на боковых сторонах. На уровне брюшной присоски толщина продольного слоя на спинной и боковых сторонах тела остается прежней. На брюшной стороне тела этот слой, напротив, достигает максимальной толщины и состоит из овальных (редко округлых) в сечении волокон диаметром 7.4–17.2 × 4.9–17.2 мкм. Расстояние между соседними продольными мышцами варьирует на всех участках покровов от 1 до 2 собственных диаметров. Диагональные мышцы почти всюду расположены на уровне цитонов тегумента и только на уровне ротовой присоски – под ними. Они пересекаются под тупым углом. Толщина диагональных мышц в передней и задней третях тела 8.4–11.2 мкм. Здесь расстояние между соседними мышечными элементами в 2 раза больше их толщины. В средней трети тела их толщина составляет 14–16.8 мкм, а расстояние между ними соизмеримо с их толщиной.
Тело C. calicophorum грушевидной формы, слегка изогнутое на брюшную сторону, в поперечном сечении округлое или немного сжатое дорсовентрально. Ротовая присоска терминальная, с небольшой предротовой губой вокруг устья. Брюшная присоска напоминает присоску P. cervi, но имеет субвентральное устье.
Толщина наружной пластинки тегумента на поверхности тела 11.2–27.5, в полости ротовой присоски 4.9–12.3, в полости брюшной присоски 12.3–14.8 мкм. Толщина базальной пластинки 2.8–5.6 мкм. Кольцевые мышцы расположены вплотную одна к другой, овальные или округлые в сечении (рис. 2). Их диаметр на уровне ротовой присоски 5.6–14 × 5.6–14, на уровне полового атриума 8.4–16.8 × 2.8–5.6, позади атриума 5.6–8.4 × × 2.8–5.6 мкм. Слой продольных мышц представлен овальными или округлыми в сечении волокнами с редкими вкраплениями поперечно-овальных на боковых сторонах тела (табл. 1). Толщина слоя максимальна на уровне семенников на спинной и брюшной сторонах тела и уменьшается на его концах, причем на переднем более заметно. На боковых сторонах тела продольный слой очень тонкий. Кроме того, есть разница в частоте расположения соседних мышечных элементов: на спинной стороне тела расстояние между ними составляет один собственный диаметр, на брюшной – меньше него, а на боковых сторонах оно может быть на порядок больше диаметра, достигая 56 мкм. Диагональные мышцы располагаются на уровне цитонов тегумента. Угол их пересечения тупой, и на срезах они выглядят почти как кольцевые мышцы. Самые толстые из них (14–16.8 мкм) находятся на уровне полового атриума. На остальных участках тела их толщина 11.2–14 мкм. Соседние мышечные элементы отстоят один от другого на 2–3 собственных толщины.
Рис. 2.
Покровы Calicophoron calicophorum. а, б – поперечный срез в средней части тела на брюшной и боковой сторонах соответственно; в – срединный сагиттальный срез на спинной стороне тела на уровне ротовой присоски.
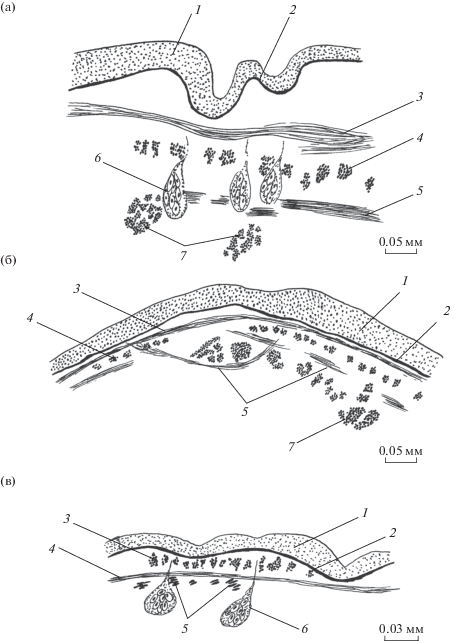
Таблица 1.
Диаметры продольных мышц в покровах Calicophoron calicophorum, мкм
| Уровень | Сторона тела | ||
|---|---|---|---|
| брюшная | боковая | спинная | |
| Ротовая присоска | 11.2–19.6 × 8.4–14 | 2.8–8.4 × 8.4 | 8.4–11.2 × 5.6–11.2 |
| Половой атриум | 11.2–19.6 × 11.2–19.6 | 1–2.8 × 1–2.8 | 11.2–19.6 × 8.4–14 |
| Семенники | 25.2–28 × 14–22.4 | 2.8–5.6 × 2.8–8.4 | 25.2–28 × 14–22.4 |
| Брюшная присоска | 19.6–22.4 × 11.2–14 | 2.8–5.6 × 2.8 | 14–22.4 × 8.4–11.2 |
S. subtriquetrus – крупные трематоды с телом грушевидной формы, значительно расширяющимся от переднего конца к заднему. Задняя половина тела несколько сжата дорсовентрально. Терминальная ротовая присоска крупная, бочонковидная. Спереди к ней примыкает мышечный колпачок. Предротовая губа маленькая. Брюшная присоска сравнительно небольшая, с вентральным устьем.
Толщина наружной пластинки тегумента 14–16.8 мкм. В устьях присосок она достигает 25.2 мкм, а в их полостях сравнительно тонкая (7.4–12.3 и 4.9–6.1 мкм). Базальная пластинка на уровне брюшной присоски не толще 2.8, а на прочих участках покровов – 11.2 мкм. Слой кольцевой мускулатуры неодинаков в разных частях тела. На большей части покровов он представлен мышечными пучками диаметром 2.6–12.3 × 2.6–14 мкм, расположенными на расстоянии 1–2 собственных диаметров один от другого. Пучки состоят из округлых в сечении волокон диаметром 1.3–3.8 мкм (рис. 3). Возле концов тела пучки распадаются на отдельные мышечные волокна. На уровне передней половины ротовой присоски, а также на брюшной стороне тела около устья брюшной присоски волокна имеют указанный выше диаметр и отстоят одно от другого на расстоянии меньше него. На спинной стороне тела на уровне брюшной присоски волокна поперечно-овальные в сечении (1.3–2.5 × 2.5–3.8 мкм) при расстоянии между ними 1–5 собственных диаметров.
Рис. 3.
Покровы Stichorchis subtriquetrus. а, б – срединный сагиттальный срез на спинной стороне тела в его средней части и на уровне брюшной присоски соответственно; в – поперечный срез на боковой стороне тела в его средней части.
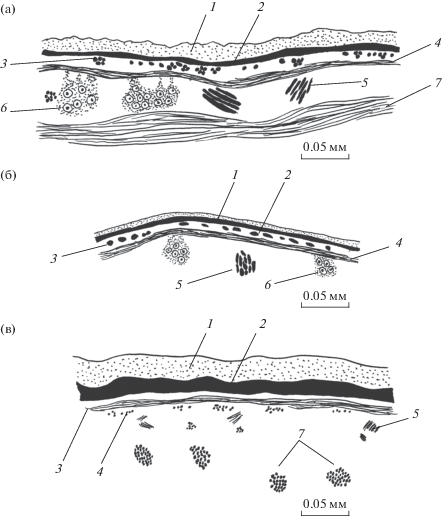
Продольный слой представлен мышечными пучками, удаленными один от другого на 1.5–2 собственных диаметра. Каждый пучок состоит из небольшого числа рыхло расположенных, округлых в сечении волокон диаметром 1.2–2.5 мкм. Сами пучки в основном поперечно-овальные в сечении, и лишь в задней трети тела на латеровентральных участках покровов они более плотные и округлые в сечении. В передней трети тела толщина продольного слоя минимальная и одинаковая на всех сторонах тела (табл. 2). В средней и задней третях тела толщина продольного слоя на брюшной стороне остается прежней, на спинной незначительно увеличивается, а на латеровентральных участках покровов возрастает вдвое. Под продольными мышцами, на уровне цитонов тегумента располагаются диагональные, пересекающиеся под тупым углом, толщиной 7.4–17.2 мкм, на расстоянии 29.5–49.2 мкм одна от другой. Данный слой не доходит ни до одного из концов тела, прослеживаясь примерно до срединной части каждой из присосок.
Таблица 2.
Диаметры продольных мышечных пучков в покровах Stichorchis subtriquetrus, мкм
| Треть тела | Сторона тела | ||
|---|---|---|---|
| брюшная | латеровентральная | спинная | |
| Передняя | 5–12.5 × 7.5–10 | 5–12.5 × 7.5–10 | 5–12.5 × 7.5–10 |
| Средняя | 7.5–12.5 × 5–12.5 | 10–25 × 15–40 | 10–15 × 20–25 |
| Задняя | 10–12.5 × 12.5–30 | 12.5–25 × 12.5–25 | 10–15 × 10–32.5 |
D. mehrai – мелкие трематоды конической формы, слегка согнутые вентрально. Ротовая присоска относительно крупная, субтерминальная, с двумя дивертикулами. Брюшная присоска с субвентральным устьем. Ее диаметр практически совпадает с максимальным диаметром тела.
Толщина наружной пластинки тегумента 4.4–5 на поверхности тела и 1–2 мкм в полостях присосок. Толщина базальной пластинки 0.5–1.2 мкм, а в устьях присосок эта пластинка более тонкая. Мышечные слои покровов вплотную прилегают один к другому, особенно на спинной стороне тела (рис. 4). Кольцевые мышцы регулярно расположенные, в основном округлые, реже поперечно-овальные в сечении, диаметром на уровне присосок 1, между присосками – 1.7–2.5 × 2–2.5 мкм. На спинной стороне тела расстояние между соседними мышечными волокнами не превышает 1 диаметра, на брюшной стороне они расположены вплотную одно к другому. Продольные мышцы в подавляющем большинстве поперечно-овальные в сечении. Их диаметр на брюшной и боковых сторонах тела наибольший (2–2.5 × 3.5–5 мкм) при расстоянии одной от другой не более 1 диаметра. На спинной стороне они тоньше (0.5–1 × 1 мкм) и разнесены на расстояние до 3 диаметров. Диагональные мышцы толщиной 2.8–5.6 мкм пересекаются под тупым углом. Расстояние между соседними мышечными элементами превышает их толщину вдвое.
Рис. 4.
Покровы Diplodiscus mehrai. а, б – поперечный срез на брюшной и спинной сторонах тела соответственно; в – срединный сагиттальный срез на брюшной стороне тела.
G. hominis – крупные трематоды с сильно расширяющимся в задней половине телом. Расширенная часть тела уплощенная, с обширной вентральной впадиной и подогнутыми на брюшную сторону тонкими краями. Ротовая присоска небольшая, с субтерминальным устьем и двумя дивертикулами. Брюшная присоска шаровидная, с вентральным устьем. Ее диаметр в 2.5 раза меньше максимальной ширины тела.
Толщина наружной пластинки тегумента на всей поверхности тела составляет 8.4 и уменьшается до 5.6 мкм лишь возле устья ротовой присоски. В полостях ротовой и брюшной присосок она равна соответственно 2.5 и 2.5–4.9 мкм. Толщина базальной пластинки 2.8–3.2 мкм. Поперечно-овальные в сечении кольцевые мышцы диаметром 8.2–11.2 × 11.2–14 мкм расположены на расстоянии 1.5–2 собственных диаметров одна от другой (рис. 5). Возле устья ротовой присоски кольцевой слой тоньше – 5.6–8.4 × 8.4–11.2 мкм.
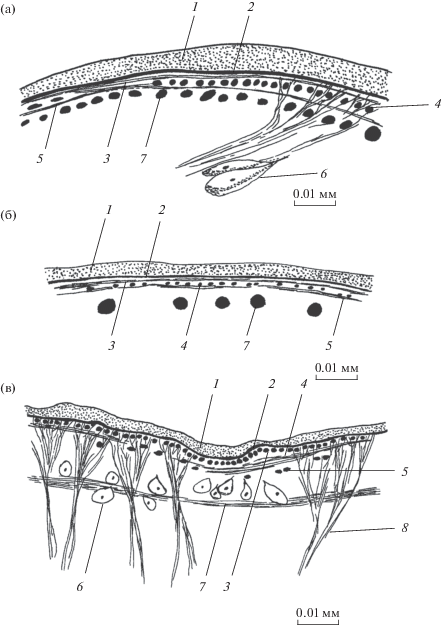
Рис. 5.
Покровы Gastrodiscoides hominis на спинной стороне тела. а – поперечный срез в средней части тела, б – срединный сагиттальный срез.
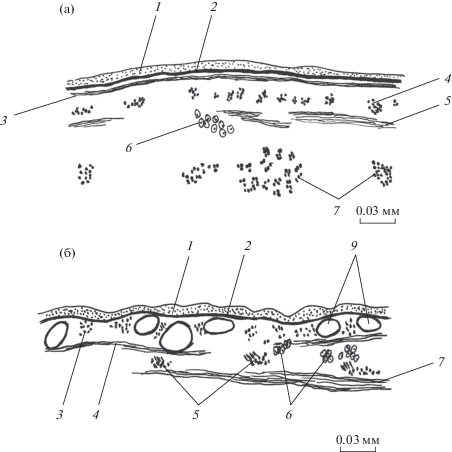
Диаметры продольных мышц возрастают в направлении спереди назад. На уровне ротовой присоски они различаются на спинной и брюшной сторонах тела и равны соответственно 4.9–7.4 × 4.9–12.3 и 4.9–14.8 × 4.9–14.8 мкм. Разница состоит также в том, что на брюшной стороне тела больше овальных в сечении мышц, и они отстоят одна от другой на 1–2 собственных диаметра, тогда как на спинной стороне большинство мышц поперечно-овальные в сечении и расстояние между ними 1–4 диаметра. Между присосками продольный слой одинаковой толщины на обеих обсуждаемых сторонах – 4.9–12.3 × 12.3–29.5 мкм при расстоянии между мышцами 1–2 диаметра. В районе брюшной присоски максимальная толщина слоя остается прежней, но отдельные мышечные элементы сильнее варьируют в размерах сечения (2.5–12.3 × 2.5–29.5 мкм) и разнесены на расстояние 1–3 собственных диаметров. На боковых сторонах тела мышечные элементы самые тонкие: в передней трети тела – 2.5–3.7 × 2.5–7.4 (округлые или поперечно-овальные в сечении), в средней трети – 2.5–6.2 × 2.5–4.9 (округлые или овальные в сечении), в задней трети – 2.5–8.6 × 2.5–12.3 мкм (все три формы сечения). Их расположение на боковых сторонах тела наименее равномерное и самое редкое – на расстоянии 1–5 собственных диаметров один от другого.
Диагональные мышцы, расположенные на уровне цитонов тегумента или немного глубже, пересекаются под тупым углом. Их толщина на спинной и брюшной сторонах уменьшается в направлении от плоскости симметрии тела к его боковым краям от 14.8–24.6 до 4.9–12.3 мкм. Расстояние между соседними мышечными элементами в 1.5–2 раза больше их толщины.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
У всех исследованных объектов толщина наружной пластинки тегумента минимальная в полостях присосок. Аналогичный факт отмечен у Lecithochirium sp. (Lecithochiriidae) и Ophiosoma patagiatum (Strigeidae) (Ястребова и др., 2004; Хахалкина, Ястребов, 2005). То же самое наблюдается в полостях некоторых других присоскоподобных структур – органа Брандеса Hysteromorpha triloba (Diplostomidae), переднего сегмента тела и полового атриума Ichthyocotylurus platycephalus (Strigeidae) (Ястребова, Ястребов, 1999; Хахалкина и др., 2002). Малая толщина наружной пластинки тегумента предположительно связана с относительно слабым механическим воздействием среды на обсуждаемые участки покровов. Напротив, в устьях обеих присосок P. cervi и S. subtriquetrus наружная пластинка тегумента имеет максимальную толщину, как у Patagifer bilobus (Echinostomatidae) и Leucochloridium macrostomum (Leucochloridiidae) (Ястребов, 1998б; Ястребов, Ястребова, 2014). Возможно, подобное кольцеобразное утолщение образуется конвергентно в разных группах трематод и служит гидравлическим амортизатором в ходе фиксации, поскольку именно устьевые зоны присосок интенсивно контактируют с субстратом. Очевидно, что минимальная толщина наружной пластинки тегумента в полостях присосок и ее максимальная толщина в устьях не являются общими правилами. Мы полагаем, однако, что дальнейшие исследования существенно увеличат число примеров такого рода.
На прочих участках покровов каждого объекта толщины наружных пластинок тегумента одинаковые, как и у другого представителя отряда – Diplodiscus subclavatus (Ястребова и др., 2000). Данное явление характерно для трематод, обитающих в механически статичных условиях, например Brandesia turgida (Pleurogenidae), Eucotyle cohni (Eucotylidae) и ряда представителей циклоцелид (Hyptiasmus arcuatus, Haematotrephus lanceolatus, Uvitellina adelphus). У реофильных трематод, напротив, нередко обнаруживаются отчетливые различия в толщинах наружных пластинок тегумента на разных участках тела. Так, у P. bilobus, Azygia robusta (Azygiidae), Halipegus ovocaudatus (Halipegidae), Tetraserialis tscherbakovi (Notocotylidae), Typhlocoelium sisowi (Cyclocoelidae) она толще на спинной стороне тела, чем на брюшной. Упоминавшийся выше L. macrostomum, а также Clinostomum complanatum (Clinostomidae) имеют иной характер различий: наружная пластинка тегумента толще в передней половине тела по сравнению с задней (Ястребов, 1997, 1998а; Ястребова и др., 2008; Ястребов, Ястребова, 2014). В обоих рассматриваемых вариантах она, по-видимому, утолщена на участках, подвергающихся максимальным механическим воздействиям. У парамфистоматид при обитании в механически агрессивной среде одинаковая толщина наружной пластинки тегумента практически на всей площади покровов, скорее всего, связана с их образом жизни, а именно с питанием одноклеточными организмами, живущими в пищеварительных органах хозяев (Готовцева, 1967; Halton, 1967; Никитин, 1985). Парамфистоматид относят к жизненной форме заднеприкрепляющихся трематод (Ошмарин, Егорова, 1978), тела которых при хищничестве обращены в просвет органа хозяина и подвергаются механическим воздействиям равномерно по всему периметру. Считать это закономерностью, общей для всего отряда, не позволяет строение Gastrothylax crumenifer (Gastrothylacidae). Наружная пластинка тегумента данного вида имеет максимальную толщину на спинной стороне тела (Ястребов, Ястребова, 2014). Рациональных объяснений этого у нас пока нет.
В покровах всех объектов числа мышечных слоев и их последовательности одинаковы. По-видимому, эти признаки довольно консервативны в пределах класса Trematoda (Smyth, 1966; Гинецинская, 1968; Smyth, Halton, 1983; Mair et al., 1998; Halton, Maule, 2004). Кольцевые и продольные мышцы образуют однорядный слой, состоящий у S. subtriquetrus из мышечных пучков, а у прочих видов – из отдельных мышечных волокон. Наличие пучков в покровах G. crumenifer (Ястребова, 2004) показывает, что оба варианта “оформления” мышечных элементов присутствуют у разных видов парамфистоматид, хотя пока не ясно в каких пропорциях. Известно, что в ряде случаев какой-либо мышечный слой (чаще всего диагональный) может отсутствовать на том или ином участке покровов сосальщиков (Ястребов и др., 2008). Подобная редукция встречается и у парамфистоматид. Мускулатура покровов двуслойна возле концов тела S. subtriquetrus, где диагональные мышцы не обнаружены. Показано также отсутствие продольной мускулатуры на уровне ротовой присоски и диагональной мускулатуры – на спинной стороне тела позади экскреторной поры D. subclavatus (Krupenko, 2013). В названных случаях редукция мышечных элементов происходит в районе массивных присасывательных органов, где подвижность покровов, возможно, ограничена.
Крупенко (Krupenko, 2013) считает мускулатуру покровов D. subclavatus четырехслойной, указывая на наличие внутренней продольной мускулатуры, расположенной под диагональной. Наши данные (Ястребова и др., 2000) заставляют усомниться в приведенном суждении. Дело в том, что внутренние продольные мышцы покровов в реальности скорее продольные паренхимные, поскольку соединяют одну с другой присоски D. subclavatus, далеко уходя от покровов в своих терминальных частях. А топографическая близость слоя к покровам еще не делает его принадлежностью к таковым.
Хотя описание паренхимной мускулатуры не входит в наши задачи, отметим вкратце, что сходная картина наблюдается у всех объектов данного исследования. У каждого из них продольные паренхимные мышцы образуют две четко обособленные группы. Первая группа (периферическая) расположена непосредственно под диагональной мускулатурой. У P. cervi она представлена не слоем, а всего несколькими мышечными пучками на спинной и брюшной сторонах тела. И, видимо, не случайно другие авторы также указывают на трехслойность мускулатуры в покровах этого вида (Толстенков и др., 2007). У прочих видов наблюдается слой мускулатуры разной степени разреженности, состоящий из мышечных волокон либо из мышечных пучков, концевые участки которых заметно удалены от покровов. Вторая группа имеет разное пространственное распределение, но расположена интерцекально. Мышцы обеих групп крепятся к присоскам в строго определенных, не совпадающих участках. Таким образом, отнесение обсуждаемого мышечного слоя к покровам или к паренхимной мускулатуре пока определяется авторскими интерпретациями и остается дискуссионным.
У трематод слой кольцевой мускулатуры обычно тоньше продольного, так как последний несет основную нагрузку по перемещению тела в ходе локомоции (Ястребов, Ястребова, 2014). Однако D. mehrai и G. hominis демонстрируют иную картину: продольный слой на большей части покровов практически не отличается по толщине от кольцевого, а на отдельных участках тоньше него. Можно ожидать подобного и у других парамфистоматид в силу перераспределения функциональной нагрузки между продольными мышцами покровов и расположенными рядом продольными паренхимными мышцами.
Сопоставление толщины мышечных слоев подразумевает сравнение их мощности. Однако для корректного сравнения требуются не анатомические, а морфометрические исследования, выполнить которые непросто из-за наличия региональных дифференцировок в архитектонике мускулатуры покровов. Например, минимальная толщина слоя кольцевых мышц S. subtriquetrus и D. mehrai наблюдается на уровне присосок. Она может быть объяснена тем же, чем упомянутое выше отсутствие продольной или диагональной мускулатуры в районе этих крупных органов. Брюшная присоска G. hominis невелика на фоне широкой задней части тела, и кольцевой слой наиболее тонок лишь на уровне ротовой присоски. У C. сalicophorum, напротив, он толще всего на участке от переднего конца тела до полового атриума включительно, а у P. cervi его толщина практически неизменна.
Последний вид демонстрирует также высокое постоянство толщины продольного слоя мышц, который несколько утолщен лишь на уровне присосок. У остальных видов по данному параметру наблюдаются иные варианты. D. mehrai имеет максимальную толщину продольного слоя на брюшной и боковых сторонах тела. Возможно, это связано с необходимостью прижиматься к субстрату, а также сгибаться на брюшную сторону при локомоции, поскольку парамфистоматиды передвигаются “шагами” подобно пиявкам. Не идентичная, но сходная картина отмечена в покровах S. subtriquetrus: максимальная толщина слоя наблюдается на латеровентральных участках покровов в двух задних (самых широких) третях тела. Отметим, что аналогичные латеровентральные утолщения продольных мышц обнаружены в локомоторном отделе тела Lecithochirium sp. (Ястребова и др., 2004). Для прижимания к субстрату и создания тяги при перемещении тела могут быть использованы продольные мышцы и на других участках покровов. В этой связи противоположная тенденция – заметное истончение продольного слоя на боковых сторонах тела C. сalicophorum и G. hominis. Среди других трематод такая же закономерность отмечена у P. bilobus (Ястребов, Ястребова, 2014).
Региональные дифференцировки кольцевой и продольной мускулатуры определяются изменениями не только толщины, но и разреженности мышечных слоев. В частности, расстояние между соседними кольцевыми мышцами C. сalicophorum и G. hominis, измеряемое в относительных величинах, остается практически неизменным на всей площади покровов. S. subtriquetrus отличается от этих видов лишь более тесным расположением мышечных элементов на брюшной стороне тела в районе брюшной присоски, а P. сervi и D. mehrai – на всей брюшной стороне. С мышцами продольного слоя ситуация иная. Постоянное расстояние между ними имеют P. сervi и S. subtriquetrus. У D. mehrai, C. сalicophorum и G. hominis продольные мышцы более всего разрежены там, где слой тоньше всего – на спинной стороне тела у первого вида и на боковых сторонах у двух других.
В архитектонике диагональной мускулатуры в отличие от кольцевой и продольной обращают на себя внимание высокая регулярность расположения мышечных элементов и незначительные изменения их толщины. У четырех видов из пяти расстояние между соседними волокнами в ряду постоянно. Лишь у P. cervi они расположены более часто в средней трети тела, не занятой присосками, и, кроме того, имеют на этом участке максимальную толщину. Толщина диагональных мышц G. hominis меняется иначе, уменьшаясь от плоскости симметрии тела в латеральных направлениях.Точно такой же характер изменений наблюдается у представителя циклоцелид T. sisowi (Ястребова и др., 2008). Оба вида, несмотря на весьма отдаленное родство, объединяет сильно уплощенное тело с тонкими краями, на которых мощная мускулатура не требуется. У остальных трех объектов толщина диагональных мышц постоянна на всех участках покровов, где они имеются.
Все обсуждаемые виды, а также D. subclavatus и G. crumenifer (Ястребова и др., 2000; Ястребов, Ястребова, 2014) имеют общую черту – тупой угол пересечения диагональных мышц. Это обусловливает высокую степень синергизма диагонального мышечного слоя относительно слабому кольцевому слою при удлинении тела, которое у представителей отряда, как правило, массивно. Данная черта строения, по всей вероятности, распространена в классе Trematoda довольно широко. Так, Судариков (Судариков, 1959) в морфологической характеристике отряда Strigeidida декларирует тупой угол пересечения диагональных мышц как общий признак, хотя не ясно, на каком именно материале сделано это обобщение. Однако упомянутые выше виды стригеидид, а также ряд других (Alaria alata, Cardiocephalus longicollis, Apharyngostrigea cornu, Pseudoapatemon tiaratus) действительно имеют названный признак (Хахалкина и др., 2004; Белова и др., 2008а, б; Белова, Ястребов, 2010). Он обнаружен также у представителей иных групп трематод – Clinostomidae, Lecithochiriidae, Leucochloridiidae, Eucotylidae, Cyclocoelidae, Azygiidae (Ястребов, 1997, 1998б; Ястребова и др., 2000, 2004, 2008; Смирнова и др., 2010). При этом у A. robusta тупой угол пересечения диагональных мышц в локомоторном отделе тела сменяется острым углом в половом отделе. В случаях когда тело гельминта имеет первичную дифференцировку на локомоторный и половой отделы (Oshmarin, 1958), а также у L. macrostomum наблюдается ослабление диагональной мускулатуры позади брюшной присоски. Оно выражается либо в уменьшении толщины мышц, либо в более редком их расположении. У парамфистоматид подобное ослабление не наблюдается, поскольку тело их не дифференцировано. Отметим, что среди представителей перечисленных групп, за исключением леукохлоридиид, нет мелких гельминтов. Это лишь подчеркивает адаптивную роль синергизма кольцевых и “тупоугольных” диагональных мышц в удлинении сравнительно крупного тела трематод, особенно на фоне изгоняющего потока веществ в полых органах хозяев и перистальтики самих органов.
ЗАКЛЮЧЕНИЕ
При всем сходстве образа жизни и габитуса различных видов парамфистоматид в строении их покровов нет полного единообразия. Параметры, связанные с наличием или отсутствием мышечных пучков, с толщиной и степенью разреженности мышечных слоев на разных участках тела, весьма вариабельны. Вариации отражают, по-видимому, не только особенности условий существования отдельных представителей отряда, но также разные способы освоения среды гельминтами. Представления о многообразии способов несомненно будут расширены в ходе дальнейших морфологических исследований таксона. Отмеченное относится не только к покровам, но также к строению органов фиксации и копулятивных аппаратов. Разнообразие архитектоники мускулатуры в сочетании с освоением широкого круга позвоночных-хозяев характеризует парамфистоматид как интенсивно эволюционирующую группу, привлекательную с точки зрения эволюционной морфологии животных.
Список литературы
Белова Н.А., Ястребов М.В. Мышечная система Cardiocephalus longicollis (Trematoda, Strigeidae) // Биоразнообразие и экология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2010. Т. 46. С. 33–47.
Белова Н.А., Ястребов М.В., Филимонова Л.В. Мышечная система Apharyngostrigea cornu (Zeder, 1800) Ciurea, 1927 (Trematoda, Strigeidae) // Систематика и биология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2008а. Т. 45. С. 5–20.
Белова Н.А., Ястребов М.В., Филимонова Л.В. Мышечная система двух представителей подотряда Strigeata (Plathelminthes, Trematoda) // Систематика и биология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2008б. Т. 45. С. 21–39.
Гинецинская Т.А. Трематоды, их жизненные циклы, биология и эволюция. Л.: Наука, 1968. 411 с.
Готовцева М.З. Особенности питания и патогенного влияния парамфистомид рубца жвачных // Докл. ВАСХНИЛ. 1967. № 3. С. 37–38.
Никитин В.Ф. Желудочно-кишечные трематодозы жвачных. М.: Агропромиздат, 1985. 240 с.
Никишин В.П. Цитоморфология скребней. М.: Геос, 2004. 234 с.
Ошмарин П.Г., Егорова М.Н. Эколого-морфологические типы трематод // Экология гельминтов. Ярославль: Изд-во ЯрГУ, 1978. С. 52–71.
Скрябин К.И. Подотряд Paramphistomatata (Szidat, 1936) Skrjabin et Schulz, 1937 // Трематоды животных и человека / Под ред. Скрябина К.И. М.; Л.: Изд-во АН СССР, 1949. Т. 3. 623 с.
Смирнова Д.А., Ястребов М.В., Ястребова И.В. Мышечная система Clinostomum foliiforme (Trematoda, Clinostomida) // Биоразнообразие и экология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2010. Т. 46. С. 257–269.
Судариков В.Е. Отряд Strigeidida (La Rue,1926) Sudarikov, 1959. Ч. 1. Морфологическая характеристика стригеидид и надсемейство Strigeoidea Railliet, 1919 // Трематоды животных и человека / Под ред. Скрябина К.И. М.: Изд-во АН СССР, 1959. Т. 16. С. 219–631.
Толстенков О.О., Теренина Н.Б., Шалаева Н.М., Гайворонская Т.В. Организация мышечной системы и распределение NO-ергических и серотонинергических элементов у трематод Allocreadium isoporum Looss, 1894 (Allocreadiidae) и Paramphistomum cervi Zeder, 1790 (Paramphistomatidae) // Зоология беспозвоночных. 2007. Т. 4. № 2. С. 139–149.
Хахалкина Н.А., Ястребов М.В. Мышечная система Ophiosoma patagiatum (Trematoda, Strigeidae) // Зоол. журн. 2005. Т. 84. № 3. С. 301–312.
Хахалкина Н.А., Ястребов М.В., Фрезе В.И. Мышечная система Hysteromorpha triloba (Trematoda, Diplostomidae) // Теоретические и прикладные проблемы паразитологии: Тр. ИНПА РАН. М.: Наука, 2002. Т. 43. С. 277–287.
Хахалкина Н.А., Ястребов М.В., Фрезе В.И. Прикрепительные и локомоторные органы Alaria alata (Goese, 1782) Krause, 1914 (Trematoda, Alariidae) // Успехи общей паразитологии: Тр. ИНПА РАН. М.: Наука, 2004. Т. 44. С. 415–427.
Ястребов М.В. Мускулатура тела некоторых трематод и фиксация фаз в эволюции присасывательной функции // Зоол. журн. 1997. Т. 76. № 6. С. 645–656.
Ястребов М.В. Строение двигательных аппаратов трематод с пассивной фиксацией на примере Haplometra cylindracea (Plagiorchiidae) и Brandesia turgida (Pleurogenidae) // Зоол. журн. 1998а. Т. 77. № 2. С. 166–176.
Ястребов М.В. Локомоторные аппараты некоторых Trematoda (Plathelminthes) с недифференцированным телом // Зоол. журн. 1998б. Т. 77. № 6. С. 627–638.
Ястребов М.В., Ястребова И.В. Мышечная система трематод (строение и возможные пути эволюции). М.: КМК, 2014. 343 с.
Ястребов М.В., Ястребова И.В., Филимонова Л.В. Об эволюционной пластичности мышечной системы гермафродитного поколения трематод (Plathelminthes, Trematoda) // Систематика и биология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2008. Т. 45. С. 184–213.
Ястребова И.В. Архитектоника мускулатуры Gastrothylax crumenifer (Creplin, 1847) (Trematoda, Gastrothylacidae) // Основные достижения и перспективы развития паразитологии: Матер. междунар. науч. конф. М.: Изд-во ИНПА РАН, 2004. С. 378–379.
Ястребова И.В., Ястребов М.В. Мускулатура стенки тела Ichthyocotylurus platycephalus (Trematoda, Strigeidae) // Зоол. журн. 1999. Т. 78. № 11. С. 1267–1274.
Ястребова И.В., Ястребов М.В., Филимонова Л.В. Мышечная система двух представителей отряда Cyclocoelida (Plathelminthes, Trematoda) // Систематика и биология паразитов: Тр. Центра паразитологии ИПЭЭ РАН. М.: Наука, 2008. Т. 45. С. 214–231.
Ястребова И.В., Ястребов М.В., Фрезе В.И. Мускулатура стенки тела некоторых реофильных представителей класса Trematoda // Актуальные проблемы общей паразитологии: Тр. ИНПА РАН. М.: Наука, 2000. Т. 42. С. 348–361.
Ястребова И.В., Ястребов М.В., Фрезе В.И. Мышечная система Lecithochirium sp. (Trematoda, Lecithochiriidae) // Успехи общей паразитологии: Тр. ИНПА РАН. М.: Наука, 2004. Т. 44. С. 433–450.
Halton D. Observations on the nutrition of digenetic trematodes // Parasitology. 1967. V. 57. № 4. P. 639–660.
Halton D., Maule A. Flatworm nerve-muscle: structural and functional analysis // Can. J. Zool. 2004. V. 2. P. 316–333.
Jones A., Bray R., Gibson D. Keys to the Trematoda. V. 2. London: CABI Publ., Natural History Museum, 2005. 745 p.
Krupenko D. Muscle system of Diplodiscus subclavatus (Trematoda: Paramphistomida) cercariae, pre-ovigerous, and ovigerous adults // Parasitol. Res. 2013. V. 113. P. 941–952.
Mair G., Maule A., Shaw C., Halton D. Muscling in on parasitic flatworms // Parasitol. Today. 1998. V. 14. № 2. P. 73–76.
Oshmarin P. On the differentiation of the body of trematodes into motor and genital parts and on the phylogenetical age of this property // Acta Veterinaria. 1958. V. 8. P. 257–263.
Smyth J. The physiology of trematodes. Edinburgh; London: Oliver & Boyd, 1966. 256 p.
Smyth J., Halton D. The physiology of trematodes. Cambridge: Cambr. Univ. Press, 1983. 446 p.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая