

Известия РАН. Серия биологическая, 2019, № 5, стр. 514-523
Таксоцен коллембол лесов Южного Приморья как эталон разнообразия и сложности организации
Н. А. Кузнецова 1, *, А. И. Бокова 1, А. К. Сараева 2, Ю. Б. Швеенкова 3
1 Московский государственный педагогический университет
129164 Москва, ул. Кибальчича, 6, коpп. 3, Россия
2 Институт леса Карельского научного центра РАН
185910 Петрозаводск, Пушкинская, 11, Россия
3 Государственный природный заповедник “Приволжская лесостепь”
440031 Пенза, ул. Окружная, 12а, Россия
* E-mail: mpnk@yandex.ru
Поступила в редакцию 04.03.2019
После доработки 15.04.2019
Принята к публикации 15.04.2019
Аннотация
Проведено сравнение показателей разнообразия сообществ коллембол реликтовых хвойно-широколиственных лесов Южного Приморья и послеледниковых лесов Русской равнины. Отмечено, что в лесах Русской равнины наиболее многочисленны широко распространенные партеногенетические формы, однократные учеты выявляют до 26 видов на 1 м2, в таксоценах нередки случаи сверхдоминирования, в образце площадью несколько квадратных сантиметров совместно встречается обычно не более трех видов одного рода. Значительно более высокие показатели видового разнообразия коллембол обнаружены в лесах Дальнего Востока: на 1 м2 лесной подстилки сосуществует до 60 видов, в одном образце может встречаться до шести видов одного рода (экологические ниши плотно “упакованы”), причем преобладают виды с узкими ареалами и амфимиктическим размножением. Обсуждаются характеристики сообществ, прошедших длительный путь исторического развития, которые можно использовать как эталонные при оценке разнообразия и сложности таксоценов коллембол в лесах умеренного пояса.
Результаты и выводы работ в области экологии сообществ существенно зависят от того, какое место занимает исследуемый объект на шкале между сложноорганизованными многовидовыми сообществами, с одной стороны, и обедненными группировками на нарушенных территориях – с другой. При оценке внешних воздействий в качестве типичных (контрольных) обычно рассматривают сообщества в наименее нарушенных, желательно заповедных, экосистемах. Большинство синэкологических исследований в России проводится в ее европейской части, испытавшей воздействие плейстоценовых оледенений. Таким образом, в качестве референтных, или контрольных на этой территории, обычно выступают исторически молодые сообщества послеледниковых ландшафтов. Правомерен вопрос: какого уровня разнообразия и сложности может достигнуть сообщество в данной природной зоне? Возможно, эти показатели зависят не только и не столько от климатических характеристик. Важен возраст самого ландшафта, поддерживающего продолжительное развитие формирующихся в нем экосистем. Речь в данном случае идет не об экологической, а об эволюционной шкале времени.
В умеренном поясе исключительным видовым богатством отличаются леса Восточной Азии (Latham, Ricklefs, 1993). В таких лесах, сохранившихся на юге Дальнего Востока России, многие отряды насекомых достигают максимальных для умеренного пояса показателей разнообразия (Чернов и др., 2011). Плейстоценовых оледенений на этой территории не было, но в периоды наступления ледников климат становился холоднее и суше, распространялись леса бореального типа (Изменение…, 1999), а при снижении среднегодовой температуры до 1–2°С – даже тундры и лесотундры (Урусов и др., 2014). При потеплении климата лесные экосистемы могли быстро обогащаться видами южной биоты благодаря не прерывавшемуся контакту с лесами более южных зон, вплоть до тропической. В результате широколиственные леса Восточной Азии, к которым относятся и леса Южного Приморья, наиболее насыщены видами растений (Latham, Ricklefs, 1993). Влияние человека на природу Южного Приморья (пожары, вырубки) в последниe 400 лет было сравнительно небольшим в отличие от такового в предыдущем историческом периоде (Урусов, Варченко, 2012).
Именно эти районы привлекали в последние годы внимание известного почвенного зоолога Б.Р. Стригановой и ее коллег (Ганин, Стриганова, 2012) как территория для решения давно обсуждаемой проблемы – “загадки видовой избыточности” почвенной фауны (Anderson, 1975; Ghilarov, 1977). В смешанных и широколиственных лесах юга Дальнего Востока было отмечено наибольшее число видов крупных почвенных беспозвоночных (Курчева, 1977; Ганин, 2011; Михалева, 2017). Фауна одной из групп почвенных микроартропод Дальнего Востока – панцирных клещей – включает в себя половину видов, отмеченных на территории всей России, а Приморский край, как и Хабаровский, отличается наибольшим разнообразием этого таксона (Рябинин, 2011).
Коллемболы, или ногохвостки, – одна из наиболее многочисленных, разнообразных и повсеместно распространенных групп мелких почвенных членистоногих. Многие виды партеногенетические и, следовательно, способны к достаточно быстрому освоению новых территорий (Tomlinson, 1966; Гиляров, 1982). Коллемболы Южного Приморья подробно описаны в цикле эколого-фаунистических исследований, проведенных в 1970–1980 гг., свидетельствующих о достаточно высоком видовом разнообразии группы в регионе (Кутырева, 1979, 1981, 1984; и др.). Современные исследования в регионе сосредоточены на описаниях новых для науки видов (Pomorski, Shveenkova, 2006; Deharveng et al., 2011; Jie et al., 2011; Jordana et al., 2011; Huang, Potapov, 2012; Smolis et al., 2012, Smolis, Deharveng, 2015). Однако в этих работах вопросы видового богатства и структуры сообществ не затрагиваются.
Достижения таксономии позволяют вернуться к рассмотрению видового разнообразия ногохвосток Южного Приморья на новом уровне. Именно эти сообщества могут быть предложены на роль своего рода “эталонных” для оценки уровня сложности таксоценов коллембол различных лесов умеренного пояса. Аналогичный подход используют фитоценологи при анализе растительного покрова, выбирая леса того же дальневосточного региона в качестве референтных (Акатов и др., 2005).
Цель работы – выяснить пределы видового богатства сообществ коллембол для лесных экосистем умеренного пояса Евразии и разработать представление о сложноорганизованном сообществе ногохвосток. Исследование позволит получить ответы на следующие вопросы: сколько близких видов может одновременно существовать в небольшом пространстве, поддерживается ли высокое разнообразие пространственной гетерогенностью среды или плотной “упаковкой” экологических ниш, является ли высокое относительное обилие партеногенетических видов в сообществах свойством коллембол как группы или особенностью послеледниковых экосистем?
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили в заповедниках Уссурийский и Кедровая Падь в кедрово-широколиственных лесах очень сложного строения с трехъярусным древостоем, “который может включать до 10 видов деревьев в первом ярусе на участке 20 × 20 м” (Растительный…, 2010). В каждом из заповедников во второй половине июля 2016 г. были изучены по две пробных площади на бурых почвах: одна – на участках с заметным участием корейской кедровой сосны Pinus koraiensis, другая – пихты чернокорой Abies holophylla.
Пробная площадь “Турова” – Уссурийский заповедник, кедрово-широколиственный лес на пологом склоне (P. koraiensis, Acer mandshuricum, Quercus mongolica и др.); в подлеске – Lonicera maackii, Eleutherococcus senticosus и др.; в напочвенном покрове – Oxalis acetosella, Carex spp., Adiantum pedatum и др. Мертвый покров составлял <40%, толщина подстилки ~10 см.
Пробная площадь “Грабовая” – Уссурийский заповедник, гора Грабовая, пихтово-грабовый лес (A. holophylla, Carpinus cordata, P. koraiensis, Betula costata и др.); в подлеске – C. cordata, Acer tegmentosum, A. barbinerve, E. senticosus и др.; в напочвенном покрове – O. acetosella, Leptorumohra amurensis и др. Мертвый покров >80%, толщина подстилки ~4 см.
Пробная площадь “Кедровая” – заповедник Кедровая Падь, кедрово-пихтово-широколиственный лес на склоне (P. koraiensis, A. holophylla, Tilia mandshurica, C. cordata и др.); в подлеске – клены пяти видов, в том числе A. tegmentosum и A. barbinerve; в напочвенном покрове – L. amurensis, Dryopteris crassirhizoma, Maianthemum dilatatum, O. acetosella и др. Мертвый покров ~50%, толщина подстилки 4 см.
Пробная площадь “Пихтовая” – заповедник Кедровая Падь, долинный пихтово-широколиственный лес (A. holophylla, Juglans mandshurica, P. koraiensis и др.); в подлеске – J. mandshurica, C. cordata, A. tegmentosum, A. mandshuricum, A. mono и др.; в напочвенном покрове – L. amurensis, D. crassirhizoma и др. Мертвый покров ~50%, толщина подстилки 3–4 см.
Структурные особенности сообществ коллембол лесов Южного Приморья сравнивали с данными, собранными в заповедниках Русской равнины – Кивач (Карелия) и Приокско-Террасный (юг Московской обл.). Были использованы материалы по мезофитным зеленомошным и гигрофитным сфагновым соснякам, описания участков приведены ранее (Сараева и др., 2015а, б; Kuznetsova, Saraeva, 2018). Сфагновые леса, несмотря на отличия гидрологического режима от такового в мезофитных лесах, были выбраны как местообитания наиболее разнообразных сообществ коллембол на Русской равнине.
Выявление полного видового разнообразия территории – труднодостижимая задача для любой группы педобионтов, включая коллембол. В связи с этим для адекватного описания разнообразия ногохвосток мы используем метод стандартной интенсивной выборки. Одна учетная серия в каждом местообитании состояла из 81 пробы, которые отбирали на трех участках, каждый площадью 1 м2, расположенных на расстоянии 10 м один от другого, по 27 проб на участке (схема учетов подробно была описана ранее (Сараева и др., 2015а, б)). Все три участка были относительно однородны и сходны по растительному покрову. Пробы отбирали буром площадью сечения 8 см2 на всю глубину лесной подстилки с включением первых сантиметров минеральных слоев почвы. Анализировали данные на следующих пространственных шкалах: 1 проба, 1 м2, местообитание, регион.
Для экстракции коллембол использовали воронки Тульгрена. Весь материал монтировали в постоянных микропрепаратах и определяли до вида. Всего было изучено 13 940 экз. коллембол 120 видов из лесов Южного Приморья и 6498 экз. 60 видов из лесов Русской равнины.
Видовое богатство (число видов на участках определенной площади) рассчитывали для нескольких масштабов: отдельных проб (8 см2), 1 м2 и отдельных местообитаний (3 м2). Мерой варьирования средних показателей было стандартное отклонение (SD). Полноту выявления видового состава оценивали с помощью индекса Jacknife 1 в программе PAST (Hammer et al., 2001). Для анализа видовой структуры использовали индексы Бергера–Паркера, Шеннона и выравненности (расчет на основе индекса Шеннона), а также ранговые распределения и отношение числа видов к числу родов. В качестве адаптивных свойств видов (traits) рассматривали два признака: способ размножения (партеногенетическое или обоеполое) и жизненную форму (эпи-, геми- и эуэдафическая). К доминантам относили виды с относительным обилием >12.4% в серии, к субдоминантам – 3.9–12.4%, малочисленным – 1.3–3.9%, редким – <1.3% (шкала Энгельмана).
Видовое разнообразие, как известно, складывается из локального альфа-разнообразия и бета-разнообразия, отражающего различия видового состава разных участков. В нашей работе оценивали вклад видового разнообразия участков площадью 1 м2 в разнообразие местообитания с помощью аддитивного парционирования. Для расчетов использовали RStudio, функцию “adipart” в пакете “vegan” (R Development…, 2011).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Общая численность ногохвосток. В кедрово-широколиственных лесах Южного Приморья этот показатель обычно существенно выше (38–75 тыс. экз./м2), чем в зеленомошных сосняках Русской равнины (9–14 тыс. экз./м2), но близок к таковому в сфагновых сосняках (31–45 тыс. экз./м2).
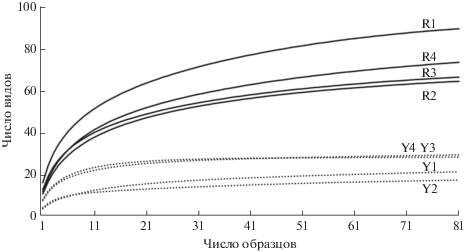
Видовое богатство сообществ коллембол. В лесах Южного Приморья оно существенно выше, чем в лесах Русской равнины, на всех рассмотренных пространственных шкалах: в одном образце (8 см2) – в среднем 10–16 vs. 3–8 видов, на 1 м2 – 46–61 vs. 13–26 видов, в одной серии в местообитании – 64–89 vs. 17–30 видов (табл. 1, 2). В выборке из 500 особей в древних лесах обнаруживается 42–51 вид, в лесах Русской равнины лишь 15–26 видов, в выборке из 100 особей – 23–27 и 11–19 видов. Кумулятивные кривые показывают более или менее быстрое выявление видового богатства в лесах Русской равнины, которое в наших сериях достигает 80–100% ожидаемого числа. Напротив, в Южном Приморье кривые так и не выходят на плато и доля обнаруженных видов в среднем ниже (рис. 1, табл. 1).
Таблица 1.
Относительное обилие доминирующих видов (% общей численности) и характеристики таксоценов коллембол в лесах Южного Приморья
| Вид, показатель | Уссурийский зап. | Зап. Кедровая Падь | ||
|---|---|---|---|---|
| “Турова” | “Грабовая” | “Кедровая” | “Пихтовая” | |
| Folsomia palearctica Potapov et Babenko | 4.4 | + | + | 13.7 |
| F. ozeana Yosii | 6.6 | 5.0 | + | 14.0 |
| F. octoculata Handschin | 18.5 | 24.6 | 24.3 | 23.3 |
| Tomocerina minuta (Tullberg) | 14.8 | 20.8 | 9.5 | 6.8 |
| Число экземпляров в серии проб | 4884 | 2491 | 2666 | 3899 |
| Общая численность, тыс. экз./м2 | 75.4 | 38.4 | 41.1 | 60.2 |
| Число видов в серии проб | 89 | 64 | 74 | 66 |
| Ожидаемое число видов по Jacknife 1 | 103 | 74 | 87 | 77 |
| Число видов на 1 м2 | 60.7 ± 5.1 | 45.7 ± 2.3 | 50.4 ± 3.2 | 45.7 ± 8.4 |
| Число видов на пробу ±SD | 15.8 ± 4.2 | 10.4 ± 3.4 | 11.4 ± 4.4 | 12.4 ± 3.3 |
| Индекс Бергера–паркера | 0.19 | 0.25 | 0.24 | 0.24 |
| Индекс Шеннона, бит | 4.28 | 3.99 | 4.24 | 3.93 |
| Выравненность | 0.66 | 0.66 | 0.68 | 0.66 |
| Среднее число видов в роде | 1.8 | 1.7 | 1.7 | 1.6 |
Таблица 2.
Относительное обилие доминирующих видов (% общей численности) и характеристики таксоценов коллембол в лесах Русской равнины
| Вид, показатель | Карелия, зап. Кивач | Приокско-Террасный зап. | ||
|---|---|---|---|---|
| зеленомошный | сфагновый | зеленомошный | сфагновый | |
| Desoria hiemalis (Schoett) | 15. 9 | 6.4 | – | – |
| Isotomiella minor (Schaeffer) | 32.1 | 20.9 | 26.5 | 13.2 |
| Mesaphorura macrochaeta Rusek | – | – | 17.1 | + |
| Parisotoma notabilis (Schaeffer) | 13 | 7.5 | 20.9 | + |
| Xenyllodes armatus Axelson | – | 9.7 | – | 39.0 |
| Число экземпляров в серии проб | 923 | 1993 | 579 | 2915 |
| Общая численность, тыс. экз./м2 | 14.2 | 30.7 | 8.9 | 45.0 |
| Число видов в серии проб | 17 | 29 | 20 | 30 |
| Ожидаемое число видов по Jacknife 1 | 20 | 32 | 25 | 30 |
| Число видов на 1 м2 | 13.0 ± 0.0 | 24.6 ± 2.5 | 15.3 ± 0.8 | 26.4 ± 1.5 |
| Число видов на пробу ±SD | 3.8 ± 1.8 | 7.3 ± 2.7 | 3.4 ± 0.2 | 8.1 ± 2.1 |
| Индекс Бергера–Паркера | 0.32 | 0. 21 | 0.27 | 0.39 |
| Индекс Шеннона, бит | 2.96 | 3.89 | 3.092 | 3.25 |
| Выравненность | 0.73 | 0.80 | 0.70 | 0.66 |
| Среднее число видов в роде | 1.1 | 1.3 | 1.1 | 1.3 |
Примечание. “+” – редкие виды (обилие <1.3% общего); “–” – вид не обнаружен; для табл. 1 и 2.
Рис. 1.
Число выявленных видов в сериях из Южного Приморья (сплошные кривые) и Русской равнины (точечные кривые) в зависимости от числа образцов. R1 и R2 – Уссурийский заповедник (“Турова” и “Грабовая” площадки соответственно), R3 и R4 – зап. Кедровая падь (“Кедровая” и “Пихтовая” площадки соответственно), Y1 и Y2 – зеленомошные, Y4 и Y3 – сфагновые сосняки заповедников Приокско-Террасного и Кивач соответственно; для рис. 1 и 2.
Таксономическая структура. Облик населения коллембол в лесах сравниваемых регионов определяется семейством Isotomidae, но в лесах Южного Приморья по сравнению с Русской равниной выше разнообразие и численность видов семейств Onychiuridae и Tomoceridae. На Русской равнине по данным однократных учетов большинство родов представлено лишь одним видом: отношение числа видов к числу родов – 1.1–1.3. В Южном Приморье эта величина составляет 1.6–1.8.
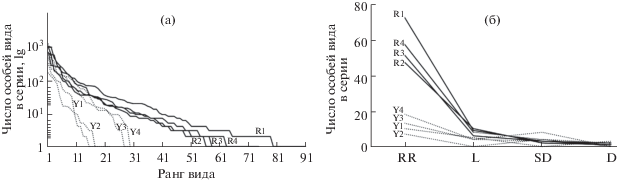
Структура доминирования. Индекс Бергера–Паркера, т.е. доля первого по численности вида, в сообществах ногохвосток Южного Приморья не превышала 0.25, наибольшее значение на Русской равнине – 0.39 (табл. 1, 2). В европейской части России отдельные учетные серии “улавливают” небольшое число видов из категории малочисленных. Ранговые распределения видов приближаются к геометрическому типу, особенно в зеленомошных сосняках. В сериях проб из Южного Приморья эти кривые более пологие, что отражает присутствие в сообществах многих видов с невысокой плотностью и близкие к логнормальному распределению численностей (рис. 2а). Интересно, что индекс выравненности обилия видов в древних лесах оказался ниже, чем в послеледниковых лесных ландшафтах. Это означает, что высокие значения индекса Шеннона (>3.9 бит в сериях из Южного Приморья, но <3.9 на Русской равнине) обусловлены в первом случае именно большим видовым богатством. Кривые Раункиера наглядно свидетельствуют о дефиците редких видов на Русской равнине (рис. 2б).
Рис. 2.
Графики ранговых распределений видов (а) и графики распределения видов по классам обилия Раункиера (б). RR – редкие виды, L – малочисленные, SD – субдоминанты, D – доминанты.
Доминирующие виды. В лесах Южного Приморья в основном преобладают виды восточно-палеарктические или с более узкими ареалами (Folsomia octoculata, F. ozeana и др.). Некоторые виды с узкими ареалами встречаются и в европейской части России, но не повсеместно, а на севере и востоке региона или в горах (например, Tomocerina minuta, Protaphorura taimyrica и Folsomia palearctica). В молодых ландшафтах Русской равнины наиболее многочисленны именно широко распространенные голарктические виды, прежде всего Isotomiella minor и Parisotoma notabilis (табл. 2).
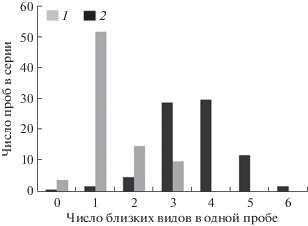
Сосуществование близких видов. Число близких видов, непосредственно контактирующих между собой в одном местообитании, дает представление о плотности “упаковки” ниш в сообществе. В обоих регионах наибольшее число видов отмечено в роде Folsomia. Для сравнения в каждом регионе была выбрана пробная площадь, где разнообразие рода было максимальным: сфагновый сосняк в Приокско-Террасном заповеднике (3 вида) и пробная площадь “Турова” в Уссурийском заповеднике (6 видов). В Приокско-Террасном заповеднике совместная встречаемость видов Folsomia была отмечена менее чем в трети проб, в Уссурийском заповеднике большинство проб включали в себя 3–4 вида, а некоторые пробы – до 6 (рис. 3).
Рис. 3.
Совместная встречаемость видов рода Folsomia в одной пробе в сериях из 81 пробы в Приокско-Террасном (1) и Уссурийском (2) заповедниках.
Cоотношение партеногенетических и амфимиктических видов. По нашей предварительной оценке партеногенетических видов коллембол в изученном материале меньше, чем обоеполых (22% списка на Русской равнине и ~9% в Южном Приморье). Однако различия сообществ коллембол этих регионов по данному признаку очевидны: на долю партеногенетических видов приходится >50% общей численности ногохвосток на послеледниковой территории и <10% в древних лесах.
Жизненные формы. В Южном Приморье по численности явно преобладают гемиэдафические формы, в то время как на Русской равнине обычно более многочисленны эуэдафические (табл. 3). В древних лесах среднее на серию число геми- и эуэдафических видов в 4 раза больше, чем на постгляциальной территории. Напротив, различия в числе эпиэдафических форм между регионами не так велики, что создает впечатление относительного дефицита видов этой группы в лесах Южного Приморья (табл. 3).
Таблица 3.
Соотношение жизненных форм коллембол в лесах Южного Приморья и Русской равнины, средние показатели по четырем пробным площадям каждого региона ±SD
| Регион | Среднее число видов | Относительное обилие, % | ||||
|---|---|---|---|---|---|---|
| эпиэдафон | гемиэдафон | эуэдафон | эпиэдафон | гемиэдафон | эуэдафон | |
| Южное Приморье | 17 ± 4 | 30 ± 5 | 27 ± 4 | 9 ± 2 | 69 ± 3 | 21 ± 3 |
| Русская равнина | 10 ± 6 | 8 ± 10 | 7 ± 10 | 20 ± 11 | 35 ± 21 | 45 ± 15 |
Пространственная структура разнообразия. Участок площадью 1 м2 включает в себя большую часть видов местообитания. По данным аддитивного парционирования эта часть несколько выше в местообитаниях Южного Приморья по сравнению с лесами Русской равнины (0.8 ± 0.07 и 0.7 ± 0.02 соответственно). Различия в видовом составе разных участков в пределах одного местообитания несколько ниже в древних лесах, чем на равнине (0.2 ± 0.07 vs. 0.3 ± 0.02). Полученные оценки вклада альфа- и бета-компонентов разнообразия во всех случаях отличались от случайных.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Влияние возраста ландшафта на разнообразие почвенных животных начали обсуждать относительно недавно. Например, в сообществах почвенных клещей доля групп обоеполых, эндемичных и редких видов, а также общее видовое богатство были выше в лесах древних ландшафтов Нидерландов по сравнению с более молодыми послеледниковыми территориями (Zaitsev et al., 2013). Об этих же закономерностях свидетельствуют и наши данные.
Фауна почвенных коллембол обнаруживает в Южном Приморье те же зоогеографические особенности, что и фауна насекомых: преобладание восточно-палеарктических и лишь небольшое число широкоареальных (голарктических, транспалеарктических, евро-сибирских) видов (Чернов и др., 2011). Интересно, что экологически близкая к коллемболам группа панцирных клещей в том же регионе почти на 70% состоит из широкоареальных видов (Рябинин, 2009).
До сих пор наиболее высокие значения видового богатства коллембол в умеренном поясе были отмечены в Центральной Японии (до 60 и 65 видов в лиственных и хвойных лесах соответственно) (Hasegava et al., 2009). В лесах Южного Приморья нами было обнаружено до 89 видов в серии. Объем изученного материала в обоих регионах был сопоставим: суммарная площадь проб при однократных учетах в одном местообитании 600 см2 в сериях из Японии и 648 см2 в наших сериях из Южного Приморья.
Высокое видовое богатство сообществ ногохвосток в древних лесах Южного Приморья связано с большим числом редких видов. Кажется, что это противоречит нашим сведениям о Русской равнине, где многолетний мониторинг населения коллембол одного и того же лесного участка ежегодно выявлял для него все новые и новые виды (Кузнецова, 2007). Обобщенная за 8 лет выборка из ~50 тыс. экземпляров показала, что редких видов в сообществе ~35% и их различие по обилию с доминантами достигает 4 порядков. Однако в данной работе обсуждаются редкие виды, учитываемые однократной выборкой, и таких видов в сериях Русской равнины существенно меньше, чем в Южном Приморье. По-видимому, это означает, что редкие виды Южного Приморья все-таки чаще встречаются и имеют более высокую плотность популяций, чем редкие виды коллембол Русской равнины.
Явное доминирование партеногенетических видов, особенно P. notabilis и I. minor, на Русской равнине становится более понятным с точки зрения геологически недавнего освобождения этой территории от плейстоценовых оледенений, последствия которых все более полно вырисовываются (Изменение…, 1999; Эволюция…, 2008; и др.). Даже одиночные особи партеногенетиков, попавшие в благоприятные условия, потенциально могут дать начало новым популяциям. Особенно много партеногенетических видов среди эуэдафических ногохвосток, обитающих глубже видов других жизненных форм (Чернова и др., 2009). Именно эти виды особенно многочисленны в лесах Русской равнины. Медленно распространяющиеся обоеполые виды, напротив, имеют конкурентное преимущество в стабильных условиях, что соотносится с преобладанием таких видов в древних лесах Южного Приморья. Отмечено, что адаптивные черты (с одной стороны, способность к эффективному расселению, а с другой – к успешному конкурированию) находятся между собой в отношениях компромисса (trade-off) (Kneitel, Chase, 2004). Можно предположить, что партеногенез положительно коррелирует со скоростью распространения, если иметь в виду скорость не передвижения, а освоения нового пригодного для обитания пространства.
Филогеографические исследования трех видов коллембол из разных районов Европы показали, что генетические дистанции между популяциями и внутри популяций каждого вида были выше на юге региона по сравнению с таковыми в центральных и северных районах. Это соответствует представлениям о заселении в голоцене севера Европы видами с южных территорий (von Saltzwedel et al., 2016). Ранняя дивергенция выявленных генетических линий (20–5 млн лет назад) свидетельствует о том, что почвенные микроартроподы – “живые ископаемые” миоцена – успешно пережили плейстоценовые оледенения в рефугиумах, откуда распространились затем на послеледниковые территории.
Многолетние исследования коллембол Русской равнины показали, что геометрический характер кривых рангового распределения – типичное явление даже для лесов заповедных территорий (Кузнецова, 2003). Ногохвостки выглядели исключением из правила, согласно которому такие кривые распределения свойственны обедненным группировкам в суровых условиях, где малое число видов захватывает основную часть ресурсов (Мэгарран, 1992). Однако в лесах Южного Приморья у ногохвосток распределение иного типа, близкое к логнормальному, что отражает более равномерное разделение ресурсов между видами и указывает на “зрелое и разнообразное естественное сообщество” (Мэгарран, 1992, с. 25). Таким образом, особенности этого аспекта видовой структуры сообществ коллембол Русской равнины свидетельствуют скорее о незавершенности процесса их формирования и соответствуют стадии “захвата ресурсов” немногими быстро осваивающими новые территории видами.
Конкурентное высвобождение, т.е. рост плотности популяции одного из видов, наблюдаемый в отсутствие сдерживающих конкурентов, особенно ярко проявляется в островных фаунах (MacArthur et al., 1972). На материках такой феномен отмечен в обедненных видами сообществах, где компенсационные процессы приводят к сверхдоминированию отдельных видов (Чернов, 2005). Это явление, при котором доля какого-либо вида превышала 40% общей численности населения, мы не раз отмечали в лесах Русской равнины (Kuznetsova, 2002). Сверхдоминирование, по-видимому, также может рассматриваться как показатель недостаточной видовой насыщенности и неполночленности сообществ. Доля участия одного вида в общей численности коллембол древних лесов Южного Приморья по нашим (индекс Бергера–Паркера, табл. 1) и опубликованным ранее (Кутырева, 1979) данным ни разу не превышала 0.25.
Высокое разнообразие может достигаться как более плотной “упаковкой” экологических ниш (т.е. локальным сосуществованием), так и мозаичностью распределения видов. В древних лесах обнаружено именно тесное сосуществование популяций разных видов, при котором на площади нескольких квадратных сантиметров лесной подстилки уживается в среднем >10 (максимально 26) видов. Интересно, что в таком ограниченном пространстве может встречаться даже по нескольку крупных видов Tomoceridae. Мозаичность распределения, несомненно, важна, однако вклад этого компонента в общее разнообразие ногохвосток древних лесов по данным аддитивного парционирования даже несколько ниже, чем на постгляциальных территориях. Другими словами, участки площадью 1 м2 внутри одного местообитания более сходны между собой по видовому составу ногохвосток в древних лесах, чем в послеледниковых. Сообщества панцирных клещей в лесах на территории Нидерландов также были более сходными между собой в древних ландшафтах и сильнее различались на более молодых территориях (Zaitsev et al., 2013). Авторы связывают это явление с ростом участия в населении более конкурентоспособных обоеполых видов.
Одна из черт сообществ ногохвосток Южного Приморья пока не находит убедительного объяснения – это чрезвычайное развитие гемиэдафической группы жизненных форм, на долю которых приходится ~70% общей численности. Это явление было отмечено ранее именно для зональных бурых почв под древними хвойно-широколиственными лесами (Кутырева, 1981). Преобладание гемиэдафических форм требует дальнейшего изучения и пока не может рассматриваться в качестве характерной черты сложноорганизованных сообществ ногохвосток.
ЗАКЛЮЧЕНИЕ
Полученные материалы, дополненные опубликованными ранее данными, позволяют описать признаки сложноорганизованного сообщества ногохвосток, сложившегося в результате длительного исторического развития. К ним относятся следующие критерии: суммарное обилие амфимиктических видов существенно больше, чем партеногенетических; большинство видов не относится к широко распространенным, видовая насыщенность >50 видов на 1 м2, много редких видов; ранговое распределение по численности близко к логнормальному; отсутствие феномена сверхдоминирования; плотная “упаковка” экологических ниш, при которой несколько (5–6) видов одного рода сосуществуют в пространстве лесной подстилки объемом ~100 мл.
Сведения о сообществах коллембол древних лесов Южного Приморья показывают, что таксоцены ногохвосток Русской равнины относительно молодые, недостаточно насыщенные видами, с небольшим числом реально контактирующих близких видов, нередкими случаями сверхдоминирования, геометрическим характером кривых рангового распределения видов и преобладанием широко распространенных партеногенетических форм.
Наши материалы подтверждают, что возраст территории играет большую роль в формировании сообществ почвенных животных. Из этого следует, что при характеристике их структуры нужно различать два варианта референтных (контрольных) сообществ в лесных экосистемах умеренного пояса, свойственных исторически молодым и древним ландшафтам.
Авторы благодарны М.Б. Потапову и А.Б. Бабенко за уточнение видовой принадлежности ряда видов, А.В. Куприну, А.Ю. Короткевич и П.А. Короткевич за помощь в сборе материала, администрации Уссурийского заповедника и национального парка “Земля леопарда” (в состав которого входит заповедник Кедровая Падь) за содействие в проведении полевых исследований.
Работа выполнена при финансовой поддержке РФФИ (гранты 16-04-01228 (Н.А. Кузнецова и А.К. Сараева) и 18-54-53032 ГФЕН-а (А.И. Бокова и Ю.Б. Швеенкова)).
Список литературы
Акатов В.В., Чефранов С.Г., Акатова Т.В. Роль исторических процессов в определении современного видового богатства древесного яруса лесов Западного Кавказа // Журн. общ. биологии. 2005. Т. 66. № 6. С. 459–470.
Ганин Г.Н. Структурно-функциональная организация сообществ мезопедобионтов юга Дальнего Востока России. Владивосток: Дальнаука, 2011. 380 с.
Ганин Г.Н., Стриганова Б.Р. Факторы формирования и поддержания избыточного видового разнообразия животного населения почв (на примере Дальнего Востока) // Изв. РАН. Сер. биол. 2012. № 3. С. 346–360.
Гиляров М.С. Экологическое значение партеногенеза // Успехи соврем. биологии. 1982. Т. 93. № 1. С. 10–22.
Изменение климата и ландшафтов за последние 65 миллионов лет (кайнозой: от палеоцена до голоцена) / Ред. Величко А.А. М.: ГЕОС, 1999. 260 с.
Кузнецова Н.А. Новые подходы к оценке структурной организации сообществ коллембол (Hexapoda: Collembola) // Экология. 2003. № 4. С. 281–288.
Кузнецова Н.А. Многолетняя динамика популяций коллембол в лесной и производной экосистемах // Зоол. журн. 2007. Т. 86. № 1. С. 30–43.
Курчева Г.Ф. Почвенные беспозвоночные советского Дальнего Востока. М.: Наука, 1977. 130 с.
Кутырева Л.Т. Фауна и население коллембол в почвах елово-широколиственных лесов Южного Приморья // Фауна и экология беспозвоночных. М.: МГПИ, 1979. С. 53–60.
Кутырева Л.Т. Соотношение морфоэкологических групп ногохвосток в различных почвах Южного Приморья // Проблемы почвенной зоологии. Киев: Наук. думка, 1981. С. 115–116.
Кутырева Л.Т. Ногохвостки во вторичных дубняках Приморского края // Фауна и экология ногохвосток. М.: Наука, 1984. С. 118–123.
Михалева Е.В. Фауна двупарноногих многоножек (Diplopoda) азиатской части России. Владивосток: Дальнаука, 2017. 336 с.
Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 161 с.
Растительный мир Уссурийской тайги: полевой атлас-определитель. Владивосток: Изд-во Дальневосточного федерального ун-та, 2010. 476 с.
Рябинин Н.А. Особенности распространения панцирных клещей (Acariformes: Oribatida) в почвах Дальнего Востока России // Вестн. ДВО РАН. 2009. № 3. С. 54–60.
Рябинин Н.А. Биологическое разнообразие панцирных клещей (Oribatida) Дальнего Востока России // Амурск. зоол. журн. 2011. Т. 3. № 1. С. 11–15.
Сараева А.К., Потапов М.Б., Кузнецова H.А. Разномасштабное распределение коллембол (Collembola) в однородном напочвенном покрове: сфагновый мох // Зоол. журн. 2015а. Т. 94. № 5. С. 517–537.
Сараева А.К., Потапов М.Б., Кузнецова H.А. Разномасштабное распределение коллембол (Collembola) в однородном напочвенном покрове: устойчивость параметров в пространстве и времени // Зоол. журн. 2015б. Т. 94. № 9. С. 1029–1045.
Урусов В.М., Варченко Л.И. Южноприморские леса с северокорейскими флористическими элементами: география, происхождение, судьба // Вестн. КрасГАУ. 2012. № 12. С. 66–73.
Урусов В.М., Врищ Д.Л., Варченко Л.И. Узловые моменты эволюции флор и ландшафтов Дальнего Востока в мезозое-кайнозое // Геогр. вестн. 2014. № 3(30). С. 26–37.
Чернов Ю.И. Видовое разнообразие и компенсационные явления в сообществах и биотических системах // Зоол. журн. 2005. Т. 84. № 10. С. 1221–1238.
Чернов Ю.И., Лелей А.С., Стороженко С.Ю. Таксономическое разнообразие насекомых Дальнего Востока России // Определитель насекомых Дальнего Востока России. Дополнительный том. Анализ фауны и общий указатель названий. Владивосток: Дальнаука, 2011. С. 7–45.
Чернова Н.М., Потапов М.Б., Савенкова Ю.Ю., Бокова А.И. Экологическая роль партеногенеза у коллембол // Зоол. журн. 2009. Т. 88. № 12. С. 1455–1470.
Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н.) / Ред. Маркова А.К., ван Кольфсхотен Т., Бохнкке Ш., Косинцев П.А., Мол И., Пузаченко А.Ю., Симакова А.Н., Смирнов Н.Г., Верпоорте А., Головачев И.Б. М.: КМК, 2008. 556 с.
Anderson J.M. The enigma of soil animal species diversity // Progress in soil zoology. Prague: Academia, 1975. P. 51–58.
Deharveng L., Bedos A., Weiner W. Two new species of the genus Leenurina Najt & Weiner, 1992 (Collembola, Neanuridae, Caputanurininae) from Primorskij Kraj (Russia) // ZooKeys. 2011. V. 115. P. 39–52.
Ghilarov M.S. Why so many species and so many individuals can coexist in the soil? // Ecol. Bull. (Stockholm). 1977. V. 25. P. 593–597.
Hammer Ř., Harper D.A.T., Ryan P.D. PAST: Paleontological statistics software package for education and data analysis. 2001. Palaeontologia Electronica 4,1. http://nhm2.uio.no/norlex/past/download.html
Hasegawa M., Fukuama K., Makino S., Okochi I., Tanaka H., Okabe K., Goto H., Mizoguchi T., Sakata T. Collembolan community in boreal-leaved forests and in conifer stands of Cryptomeria japonica in Central Japan // Pesg. Agropec. Bras. Brasilia. 2009. V. 44. № 8. P. 881–890.
Huang C.-W., Potapov M. Taxonomy of the Proisotoma complex. IV. Notes on chaetotaxy of femur and description of new species of Scutisotoma and Weberacantha from Asia // Zootaxa. 2012. № 3333. P. 38–49.
Jie D., Potapov M., Sokolova E. Further study on the labial palp in the Isotomidae (Collembola) with reference to the genus Heteroisotoma Stach // Zootaxa. 2011. № 2991. P 49–61.
Jordana R., Potapov M., Baquero E. New species of Entomobryini from Russia and Armenia (Collembola, Entomobryomorpha) // Soil Organisms. 2011. V. 83. № 2. P. 221–248.
Kneitel J.M., Chase J.M. Trade-offs in community ecology: linking spatial scales and species coexistence // Ecol. Lett. 2004. № 7. P. 69–80.
Kuznetsova N.A. Classification of collembolan communities in the East-European taiga // Pedobiologia. 2002. V. 46. № 3–4. P. 373–384.
Kuznetsova N.A., Saraeva A.K. Beta-diversity partitioning approach in soil zoology: A case of Collembola in pine forests // Geoderma. 2018. V. 332. P. 142–152.
Latham R.E., Ricklefs R.E. Continental comparisons of temperate-zone tree species diversity // Species diversity in ecological communities: historical and geographical perspectives. Chicago: Univ. Chicago Press, 1993. P. 294–315.
MacArthur R.H., Diamond J.R., Karr J.R. Density compensation in island faunas // Ecology. 1972. V. 53. № 2. P. 330–342.
Pomorski R.J., Sveenkova Y.B. New genus with three new species of Thalassaphorurini (Collembola: Onychiuridae) from Russian Far East // Insect Syst. Evol. 2006. V. 37. P. 191–196.
R Development Core Team. R: A Language and Environment for Statistical Computing. 2011. Vers. 2.13.1. http://www.r-project.org.
Smolis A., Deharveng L. Diversity of Paranura Axelson, 1902 (Collembola: Neanuridae: Neanurinae) in Pacific Region of Russia and United States // Zootaxa. 2015. № 4033. P. 2.
Smolis A., Deharveng L., Bedos A. Description of two new species of the genus Micranurida Borner, 1901 (Collembola: Neanuridae, Pseudachorutinae) from Russia, with notes on the genus Lanzhotia Rusek, 1985 // Entomol. Fenn. 2012. V. 23. P. 149–154.
Tomlinsson J. The Advantages of Hermaphroditism and Parthenogenesis // J. Theoret. Biol. 1966. V. 11. P. 54–58.
Von Saltzwede H., Scheu S., Schaefer I. Founder events and pre-glacial divergences shape the genetic structure of European Collembola species // BMC Evol. Biol. 2016. V. 23. P. 81–87.
Zaitsev A.S., van Straalen N.M., Berg M.P. Landscape geological age explains large scale spatial trends in oribatid mite diversity // Landscape Ecol. 2013. V. 28. P. 285–296.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая