

Известия РАН. Серия биологическая, 2020, № 3, стр. 274-282
Пространственная структура популяции, возрастной состав и семенная продуктивность Delphinium macropogon Prokh. в Талгинском ущелье
Д. М. Анатов *
Горный ботанический сад ДФИЦ РАН
367000 Махачкала, ул. М. Гаджиева, 45, Россия
* E-mail: djalal@list.ru
Поступила в редакцию 01.02.2018
После доработки 26.11.2018
Принята к публикации 24.12.2018
Аннотация
Рассмотрены пространственная структура, демографические показатели и семенная продуктивность Delphinium macropogon Prokh. в Талгинском ущелье. Отмечено, что популяция D. macropogon характеризуется сильной разреженностью и низкой семенной продуктивностью; возрастной состав популяции неполночленный, с преобладанием молодых генеративных особей. Установлено, что демографические показатели, проанализированные с учетом индексов восстановления (Iв = 0.24) и виталитета (IVC = 0.88), депрессивные. По критерию Δ–ω популяция отнесена к зреющему типу. С помощью комплексного анализа популяционных характеристик D. macropogon в Талгинском ущелье обнаружено, что процессы заселения территории происходят на низком уровне плотности, чему препятствуют слабый прирост за счет молодой фракции, низкой семенной продуктивности и всхожести семян.
В настоящее время одна из приоритетных задач ботанической науки – изучение эндемичных видов в целях обоснования мер их охраны. Ранее была отмечена (Тахтаджян, 1981) необходимость при прочих равных условиях составления списка краснокнижных видов отдавать предпочтение эндемичным и особо редким видам, но, как правило, только в тех случаях, когда имелись определенные указания на грозящую им опасность. Во всех случаях предпочтение должно отдаваться тем растениям, реальная или потенциальная хозяйственная ценность которых установлена. Все это предусматривает расширенные исследования по биологии и экологии этих видов.
Анализ внутривидовой изменчивости эндемичных растений – важная составляющая ботанического, генетического ресурсоведения и необходимая предпосылка решения ряда проблем теории микроэволюции, биосистематики и популяционной биологии, а также исходный этап в селекции и интродукции новых видов.
Один из родов Кавказа, имеющих декоративное и лекарственное значение – род Delphinium L. (живокость). Этот род многолетних травянистых растений включает в себя ~370 видов, произрастающих в Северном полушарии и в горах тропической Африки. Многие виды происходят из Юго-Восточной Азии, особенно из Китая, где произрастает >150 видов. На территории России и сопредельных стран произрастает ~100 видов (Малютин, 1992). На Кавказе насчитывается >23 видов, многие из которых являются кавказскими эндемиками (Гроссгейм, 1950; Литвинская, Муртазалиев, 2009).
В Дагестане род Delphinium L. представлен 13 видами (Димитрова, 1966; Галушко, 1978; Муртазалиев, 2009), которые играют существенную роль в растительных сообществах, нередко могут быть доминантами и важными эдификаторами. На территории Дагестана вероятно существование доли полиморфных, таксономически слабо исследованных видов, характеризующихся высокой морфологической изменчивостью и содержанием алкалоидного состава.
На территории Дагестана в роде Delphinium L. преобладают виды, распространенные только на Кавказе (исключение – вид D. puniceum Pall.), среди которых высока доля эндемиков Дагестана (5 или 41.7%). Это свидетельствует о том, что Дагестан – один из центров вторичного видообразования рода Delphínium L. на Кавказе (Анатов, 2010). Первичный центр происхождения предположительно находится на территории современного Китая (Димитрова, 1969; Wang Wencai, Warnock, 2001).
Один из малоизученных эндемиков Дагестана из рода Delphinium L. – D. macropogon Prokh. (живокость крупнобородая) – многолетние растения из подрода Oligophyllon Dimitrova (Димитрова, 1969, 2013) высотой 50–100 (140) см, с цельным яйцевидным клубнем, одиночным голым стеблем, с длинночерешчатыми листьями. Кисти довольно плотные, 9–12 см длиной, многоцветковые, короткопушистые или почти голые, обыкновенно единичные, реже с немногочисленными боковыми укороченными кистями. Цветет он во второй половине июня, плодоношение и созревание происхдят в середине–конце июля (Проханов, 1961). Произрастает этот эндемик в лиственных лесах возвышенностей и ущелий Кукурт-Тау, горы Мискин-Баш, а также столовой горы Тарки-Тау на высотах 350–900 м над у. м.
Пожалуй, со времен открытия этого вида и описания морфологии Прохановым (Проханов, 1961), а также анатомических исследований черешка и стебля, проведенных Димитровой в 1960-х гг. (Димитрова, 1966, 1969), до настоящего времени D. macropogon практически не исследован, популяционно-экологические аспекты жизни вида не изучены, его статус не определен.
Цель работы – анализ пространственной структуры, возрастного состава популяции и внутрипопуляционной изменчивости особей D. macropogon по морфологическим признакам генеративного побега в Талгинском ущелье.
МАТЕРИАЛЫ И МЕТОДЫ
Полевые исследования популяции D. macropogon были проведены в Талгинском ущелье в 2016 г. с использованием общепринятых методик в популяционных анализах (Работнов, 1950; Уранов, 1975; Андреева и др., 2002; Злобин, 2009). Для выделения возрастных состояний использовали известные методики популяционного анализа (Работнов, 1950; Ценопопуляции…, 1976, 1977, 1988; Онтогенетический…, 2000). При выделении возрастных состояний учитывались следующие признаки: высота растений, число и размеры листьев, наличие боковых генеративных кистей.
Талгинское ущелье – это уникальный памятник природы, резерват многих реликтовых, эндемичных и исчезающих видов растений (рис. 1). Ущелье относится к предгорному физико-географическому и флористическому району (Муртазалиев, 2004). Оно расположено недалеко от г. Махачкалы и представляет собой изолированную аридную территорию, входящую в систему хребтов горы Кукурт-Тау. От Каспия ущелье отделено хребтами Тарки-Тау и Нарат-Тюбе. Рельеф скалистый и каменисто-щебнистый, почвенный покров развит лишь на пологих гребнях ущелья и его дне. В местах произрастания D. macropogon в почвенном покрове преобладают коричневые и бурые лесные почвы, которые по реакции pH нейтральные и слабощелочные. Среднее количество осадков 300–400 мм. Лето сухое и жаркое со среднесуточной температурой воздуха +24°С. Зима мягкая, среднесуточная температура воздуха в январе –1°С. Среднегодовая относительная влажность 65% (Акаев и др., 1996; Магомедова, Гасанова, 2014).
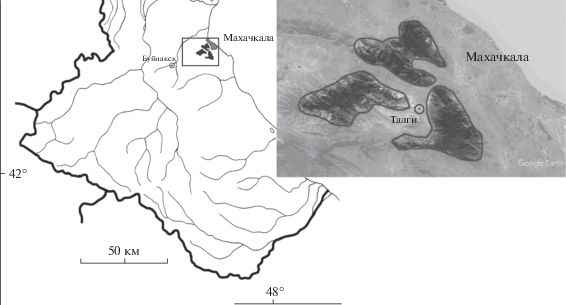
Эколого-ценотическую стратегию (ЭЦС) вида определяли по ранее опубликованным классификациям (Grime, 1974; Работнов, 1975, 1985), признаки для определения ЭЦС – по методике Блиновой (Блинова, 2014). Закладка участков и учеты проводились в середине–конце июня во время массового цветения и в середине июля во время плодоношения – начало созревания плодов.
Сообщество, где произрастает D. macropogon, относится к ясеннику дубово-кустарниково-разнотравному – Fraxetum quercoso-fruticoso-varioherbosum. Исследуемые учетные площади сосредоточены у основания известняковых склонов Талгинского ущелья, где создаются условия (в данном случае достаточное количество воды, стекаемой с вершин скал к основанию) для произрастания лесного типа растительности. Почвенный слой слабо развит (до 10 см) и формируется на поверхности образованного в процессе выветривания мелкощебнистого слоя известняков. В местах произрастания D. macropogon рельеф представлен пологими щебнистыми склонами (крутизна склонов 5–35°).
На учетных площадках с участием эндемика выявлено 26 видов цветковых растений, из которых 8 относятся к древесно-кустарниковым растениям (табл. 1). Основные доминанты древесных видов – Fraxinus excelsior LLiebl. и Cornus mas L.; в совокупности на эти виды приходится 55–80% общей сомкнутости крон. Типичные травянистые представители – Geum urbanum L., Stellaria graminea L. Campanula rapunculus L. и некоторые другие виды. По отношению к экологическим факторам большинство травянистых видов относятся к мезофитам.
Таблица 1.
Некоторые характеристики сообществ с участием Delphinium macropogon в Талгинском ущелье
| Виды | Площадки | ∑ | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 (457 м СЗ скл.) |
2 (480 м СВ скл.) |
3 (493 м С скл.) |
|||||||
| Ч | ПП | Ч | ПП | Ч | ПП | Ч | ПП | Вс | |
| Древесный ярус, сомкнутость, % | |||||||||
| Fraxinus excelsior L. | 1 | 40 | 2 | 30 | 1 | 25 | 4 | 32 | 100 |
| Quercus petraea (Matt.) Liebl. | 1 | 25 | 1 | 25 | 1 | 20 | 3 | 23 | 100 |
| ∑ | 2 | 65 | 3 | 55 | 2 | 45 | 7 | 55 | 100 |
| Кустарниковый ярус, сомкнутость, % | |||||||||
| Cornus mas L. | 3 | 15 | 1 | 10 | 1 | 10 | 5 | 12 | 100 |
| Lonicera caprifolium L. | 1 | 3 | 1 | 3 | – | – | 2 | 2 | 67 |
| Viburnum lantana L. | 1 | 5 | – | – | – | – | 1 | 2 | 33 |
| Rhus coriaria L. | 1 | 3 | – | – | – | – | 1 | 1 | 33 |
| Rhamnus cathartica L. | – | – | 1 | 2 | – | – | 1 | 1 | 33 |
| Euonymus verrucosa Scop. | – | – | – | – | 1 | 3 | 1 | 1 | 33 |
| ∑ | 6 | 26 | 3 | 15 | 2 | 13 | 11 | 19 | 50 |
| Травяной ярус, покрытие, % | |||||||||
| D. macropogon Prokh. | 21 | 7 | 19 | 2.5 | 18 | 3 | 58 | 4.2 | 100 |
| Geum urbanum L. | 25 | 8 | 17 | 5 | 15 | 4 | 60 | 5.7 | 100 |
| Stellaria graminea L. | 30 | 7 | 15 | 3.5 | 14 | 3 | 59 | 5.5 | 100 |
| Physocaulis nodosus (L.) W.D.J. Koch | 3 | 0.5 | 10 | 4 | 17 | 7 | 30 | 3.8 | 100 |
| Campanula rapunculus L. | 3 | 0.5 | 9 | 3 | 20 | 3 | 32 | 2.2 | 100 |
| Vicia balansae Boiss. | 2 | 0.1 | 4 | 0.2 | 3 | 0.1 | 9 | 0.1 | 100 |
| Viola odorata L. | 3 | 0.2 | 10 | 2 | – | – | 13 | 0.3 | 67 |
| Asparagus officinalis L. | – | – | 3 | 0.3 | – | – | 3 | 0.1 | 33 |
| Melandrium album (Mill.) Garcke | – | – | – | – | 3 | 0.2 | 3 | 0.1 | 33 |
| Dactylis glomerata L. | – | – | – | – | 2 | 0.2 | 2 | 0.1 | 33 |
| Hylotelephium caucasicum (Grossh.) H. Ohba | – | – | – | – | 2 | 0.1 | 2 | 0.1 | 33 |
| Orobanche sp. | – | – | – | – | 2 | 0.1 | 2 | 0.1 | 33 |
| Chelidonium majus L. | 1 | 0.1 | – | – | – | – | 1 | 0.1 | 33 |
| Chaerophyllum angelicifolium M. Bieb. | 1 | 0.1 | – | – | – | – | 1 | 0.1 | 33 |
| Lactuca chaixii Vill. | 1 | 0.1 | – | – | – | – | 1 | 0.1 | 33 |
| Bromopsis variegata (M. Bieb.) Holub | 1 | 0.1 | – | – | – | – | 1 | 0.1 | 33 |
| Lapsana intermedia M. Bieb. | – | – | 1 | 0.1 | – | – | 1 | 0.1 | 33 |
| Silene italica (L.) Pers. | – | – | – | – | 1 | 0.1 | 1 | 0.1 | 33 |
| Brachypodium sylvaticum (Huds.) P. Beauv. | – | – | – | – | 1 | 0.1 | 1 | 0.1 | 33 |
| ∑ | 91 | 23.7 | 88 | 20.6 | 98 | 20.9 | 280 | 21.7 | 56 |
У травянистых растений роль ведущих элементов характеристики горизонтальной структуры популяций играют плотность (среднее для данной популяции число особей на единицу площади) и тип размещения особей на площади (Greig-Smith, 1961; Harper 1977; Плотников, 1979; Злобин, 2009; Марков, 2012). Для изучения пространственной структуры были заложены три пробные площадки по 25 м2 вдоль северного склона в типичных местах их произрастания. Каждая площадка была разбита на 25 площадок размером 1 × 1 м2, где были подсчитаны все особи изучаемого вида. Были учтены 6 показателей пространственной структуры (проективное покрытие (ПП) вида, плотность в пересчете на 25 м2, число древесных видов, сомкнутость крон, ПП трав, ПП площадки (покрытие трав + фактическое покрытие древесными видами)) и 6 для изучения возрастного состава (число проростков, ювенильных, имматурных, виргинильных, молодых генеративных, зрелых генеративных особей).
Тип размещения особей оценивался по характеру распределения частот плотностей с построением графиков и последующего сопоставления с частотами теоретического распределения. В качестве оценки типа размещения особей использовалось отношение дисперсии (S 2) к средней арифметической (Х). При S 2= Х констатируют регулярное (равномерное), при S2 < Х – случайное, а при S 2 > Х – контагиозное (скученное) размещения (Плотников, 1979; Марков, 2012).
Оценка жизненности популяции проводилась по виталитету, предложенному Злобиным (Злобин, 1989). Мы отобрали десять признаков, являющихся показателями жизненной стратегии этого вида. Это признаки, влияющие на фитомассу растения и его продукционные процессы (высота генеративного побега, длина растения до главной кисти, число междоузлий, длина главного соцветия, число цветков главного соцветия, общее число цветков, плодов, семян, масса плодов и семян на растении). Степень процветания или депрессивности популяции вычисляли по индексу виталитета популяции (IQ): IQ = (a + b)/2c, где а, b, с – числа крупных, средних и мелких особей соответственно.
Изучение семенной продуктивности проводили по общепринятой методике (Ходачек, 1970, 2008; Вайнагий, 1974). Так как объект исследований – узколокальный эндемик, особи не выкапывались. Частично измерения осуществлялись в полевых условиях (растения маркировались в период массового цветения), а в период плодоношения были собраны плоды, дальнейшие подсчеты и взвешивания проводились в лаборатории. При этом учитывали числа цветков, плодов, листовок, семян, коэффициент плодоцветения (Кпц), массы плодов и семян на растении, коэффициент эффективности плода (Кэ) – отношение массы семян плода к массе плода.
Для выявления типа ценопопуляции по критерию Δ–ω были вычислены индекс возрастности популяции (Уранов, 1975), индекс эффективности популяции и эффективная плотность популяции (Животовский, 2001).
Математическую обработку результатов проводили с использованием методов описательной статистики, факторного, корреляционного анализов (Лакин, 1980). Коэффициент вариации (Cv) оценивали по шкале Мамаева (Мамаев, 1975). Для оценки среднего уровня связей применялся усредненный по всем признакам коэффициент детерминации $R_{{{\text{ch}}}}^{2}$ (Ростова, 2002). Статистическую обработку полученных результатов и построение графиков выполняли с использованием программы Statistica v.5.5.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
На рис. 2 отображено размещение особей в пространстве, где видно много пустых площадок и скученное размещение особей. Так как S 2/Х = 2.1, размещение относится к контагиозному. Встречаемость вида на 75 учетных площадках (1 × 1 м2) составляет 48%, из которых 32% – единичные особи, значительно реже встречаются площадки с двумя и тремя особями и лишь в одном случае было отмечено S 2/Х > 3 (9 особей).
Рис. 2.
Схематическое отображение пространственного размещения особей и соответствующие частоты встречаемости Delphinium macropogon на учетных площадках.
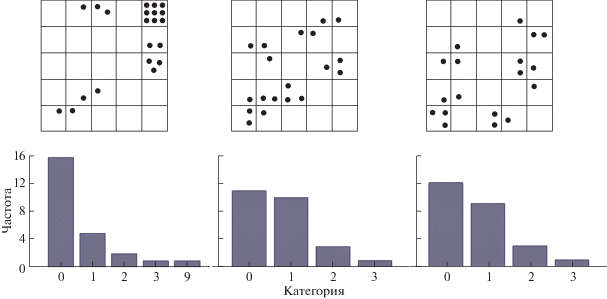
Плотность этого вида в типичных местах произрастания характеризуется как очень разреженная и в среднем составляет 19.3 особи на 25 м2 (табл. 2). ПП трав в целом невысокое (21.7%), как и общее ПП самих площадок (29.9%). Амплитуда изменчивости пространственных показателей характеризовалась как очень высокая. Средними значениями по CV, характеризовался признак “сомкнутость крон”.
Таблица 2.
Сравнительная характеристика популяции Delphinium macropogon по плотности на пробных площадках
| Признак | X ± Sx | min–max | Cv, % |
|---|---|---|---|
| ПП вида, % | 0.3 ± 0.14 | 0–10 | 396.1 |
| Плотность на 25 м2 | 19.3 ± 3.66 | 0–225 | 164.1 |
| Сомкнутость крон, % | 91.9 ± 2.30 | 10–100 | 21.6 |
| ПП трав, % | 21.7 ± 2.09 | 0–75 | 83.6 |
| ПП площадки, % | 29.9 ± 2.11 | 0–80 | 61.1 |
Исходя из полученных данных можно заключить, что по эколого-ценотической стратегии D. macropogon относится к стресс-толерантам, к группе ценотических патиентов Sn (делят ресурсы с конкурентами (Cn) в сомкнутых сообществах). С учетом полученных данных можно констатировать, что основное эколого-ценотическое условие, необходимое для существования этого вида, – высокая сомкнутость крон древесных видов (75–100%), способствующая сильному затенению, сохранению влаги и формированию низкого проективного покрытия травяного яруса, что в конечном счете благоприятствует прорастанию семян и уменьшению межвидовой конкуренции.
Высота растения D. macropogon в генеративном состоянии (g2) в среднем составляет 68.1 см, варьируя от 48 до 94 см (табл. 3). Большая часть длины растения – вегетативно-генеративная часть до главного соцветия. Размах числа междоузлий небольшой (5–9), а длина соцветия варьирует значительно (4–20 см, в среднем 7.9 см), что во многом связано с повреждающим действием насекомых фитофагов. У многих растений D. macropogon часто наблюдаются повреждения верхушек соцветий, чем обусловливаются небольшие длины соцветия и числа цветков. Из всех учтенных особей только 36.4% перешли к стадии плодоношения. Показатели семенной продуктивности также значительно варьируют. Размах варьирования числа семян на растении составляет 4–408, и в среднем одно растение продуцирует 167.4 семян, т.е. от почти полного отсутствия полноценных семян до нескольких сотен, что также не является показателем высокой семенной продуктивности. Масса 100 семян в среднем составляет 86.8 мг, коэффициент плодоцветения – 42.2%. Полевая всхожесть в среднем в условиях экспериментальных баз Горного ботанического сада ДНЦ РАН – 15.5%.
Таблица 3.
Морфометрические признаки и семенная продуктивность Delphinium macropogon в условиях Талгинского ущелья
| Признак | X ± Sx | min–max | Cv,% | $R_{{{\text{ch}}}}^{2}$,% | A | E | |
|---|---|---|---|---|---|---|---|
| Высота растения, см | 68.1 ± 2.01 | 48–94 | 17.0 | 21.9 | 0.29 | –0.13 | |
| Длина, см | |||||||
| растения до соцветия | 60.2 ± 1.85 | 42–87 | 17.6 | 16.3 | 0.42 | 0.06 | |
| главного соцветия | 7.9 ± 0.58 | 4–20 | 42.2 | 15.6 | 1.46* | 3.78* | |
| Число | |||||||
| междоузлий | 6.7 ± 0.16 | 5–9 | 13.5 | 29.8 | 0.85 | 0.76 | |
| цветков глав. соцветия | 14.4 ± 2.43 | 4–41 | 66.8 | 21.9 | 0.93 | 0.37 | |
| цветков | 21.9 ± 2.46 | 5–85 | 64.7 | 35.7 | 2.85* | 12.15* | |
| плодов | 13.6 ± 4.19 | 1–47 | 106.8 | 52.0 | 1.35 | 1.39 | |
| листовок | 37.6 ± 11.60 | 2–128 | 106.9 | 51.2 | 1.26 | 1.00 | |
| семян | 167.4 ± 45.25 | 4–408 | 93.6 | 36.9 | 0.22 | –1.93 | |
| Масса, мг | |||||||
| плодов | 267.5 ± 82.84 | 11–888 | 107.3 | 51.3 | 1.25 | 0.81 | |
| семян | 155.9 ± 46.46 | 1–445 | 103.2 | 47.7 | 0.83 | –0.47 | |
| 100 семян | 86.8 ± 10.54 | 25.0–144.7 | 42.1 | 17.6 | –0.07 | –0.53 | |
| Кпц, % | 42.2 ± 8.94 | 4–84.2 | 73.3 | 28.2 | 0.02 | –1.59 | |
| Кэ, % | 50.3 ± 5.49 | 6.3–73 | 37.8 | 19.9 | –1.10 | 1.59 | |
Показатели асимметрии в целом положительные, т.е. в распределении преобладают более низкие значения признака. Лишь коэффициент эффективности плода отрицательный.
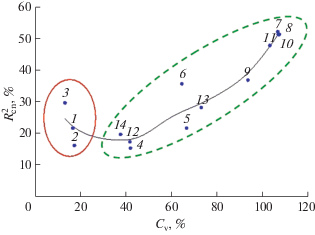
Коэффициент вариации свидетельствует об очень высоком варьировании дискретных, весовых и индексных признаков семенной продуктивности, а также о среднем изменении линейных признаков генеративного побега и числа междоузлий. Графическое изображение индивидуальной изменчивости по Cv и детерминации $\left( {R_{{{\text{ch}}}}^{2}} \right)$ наглядно показывает вклад генотипической и экологической компонент в изменчивость морфологических признаков (рис. 3). В целом эти два коэффициента отобразили высокую положительную корреляционную связь между собой (r = 0.85), кривая регрессии, пстроенная методом наименьших квадратов, выявила две совокупности признаков. В первую группу вошли признаки с наименьшей изменчивостью по Cv и $R_{{{\text{ch}}}}^{2};$ к ней относятся признаки конструкции генеративного побега. Во вторую группу вошли признаки семенной продуктивности.
Рис. 3.
Двумерная проекция индивидуальной изменчивости (Cv) и средней сопряженности $\left( {R_{{{\text{ch}}}}^{2}} \right)$ морфологических признаков. 1 – высота растения, 2 –длина растения до главного соцветия, 3 – число междоузлий, 4 – длина главного соцветия, 5 – число цветков главного соцветия, 6 – число цветков на растении, 7 – число плодов, 8 – число листовок, 9 – число семян, 10 – масса плодов, 11 – масса семян, 12 – масса 100 семян, 13 – Кпц, 14 – Кэ; r = 0.85.
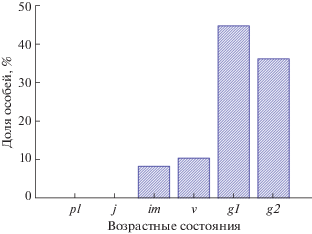
Возрастной состав популяции неполночленный, с преобладанием молодых генеративных особей (g1). Молодая часть представлена только имматурными и виргинильными (19.1%) особями, проростки и ювенильные особи не обнаружены (рис. 4). Для этого вида выделение старых генеративных особей без проведения специальных анатомических исследований каудекса визуально не представляется возможным.
Рис. 4.
Возрастной состав Delphinium macropogon в Талгинском ущелье. p – проростки, j – ювенильные особи, im – имматурные особи, v – виргинильные особи, g1 – молодые генеративные особи, g2 – зрелые генеративные особи.
Определение виталитета изучаемой популяции проводилось сначала на основе метода главных компонент, по результатам которого были выделены два главных фактора. В двумерной проекции по первому фактору все особи распределились по признакам генеративного побега, по второму – семенной продуктивности. Между самыми информативными признаками был проведен корреляционный анализ, затем визуально были рассчитаны классы растений. По итогам анализа было выявлено преобладание мелких и средних особей в популяции (а = 7, b = 14, c = 12); IVC = 0.88, Iв = 0.24. Такое критическое соотношение молодых и генеративных особей может частично объясняться высокой элиминацией проростков и ювенильных особей при относительно низкой семенной продуктивности и полевой всхожести семян.
По критерию Δ–ω популяция относится к зреющему типу. Вычисленные индексы наглядно показывают, что в местах наибольшего сосредоточения происходят процессы заселения территории, но низкая эффективная плотность (Ме) показывает, что этот процесс происходит на низком уровне плотности, чему препятствуют слабый прирост за счет молодой фракции, низкая семенная продуктивность и всхожесть семян.
ЗАКЛЮЧЕНИЕ
Популяция D. macropogon характеризуются сильной разреженностью. Плотность этого вида в типичных местах произрастания составляет 19.3 особи на 25 м2. Размещение особей относится к контагиозному типу. Встречаемость вида на учетных площадках составляет 48%, из которых 32.0% – единичные особи. Основное эколого-ценотическое условие, необходимое для существования этого вида, – высокая сомкнутость крон (75–100%), способствующая сильному затенению и, соответственно уменьшению проективного покрытия травяного яруса, сохранению влаги, что в конечном счете благоприятствует прорастанию семян и уменьшению межвидовой конкуренции в аридном климате Талгинского ущелья.
Семенная продуктивность вида невысока, в среднем на одно растение приходится 21.9 цветка, а коэффициент плодоцветения – 42.2%. Одно растение в среднем продуцирует 167.4 семян при массе 100 семян 86.8 мг.
Возрастной состав популяции неполночленный с преобладанием молодых генеративных особей (g1). Молодая часть представлена только имматурными и виргинильными (19.1%) особями, проростки и ювенильные особи не обнаружены, что может свидетельствовать о высокой гибели в молодом возрасте в совокупности с низкой семенной продуктивностью и всхожестью семян.
С помощью Iв = 0.24 и IVC = 0.88 установлена депрессивность демографических показателей. По критерию Δ–ω популяция относится к зреющему типу. Вычисленные индексы наглядно показывают, что в местах наибольшего сосредоточения происходят процессы заселения территории, но низкие значения Ме показывают, что этот процесс происходит на низком уровне плотности вследствие слабого прироста за счет молодой фракции, низких семенной продуктивности и всхожести семян.
Автор выражает благодарность Р.А. Муртазалиеву и Р.М. Османову (сотрудникам лаборатория флоры и растительных ресурсов) за помощь в проведении полевых исследований и геоботанических описаниях учетных площадок.
Список литературы
Акаев Б.А., Атаев З.В., Гаджиев Б.С. Физическая география Дагестана: уч. пособие. М.: Школа, 1996. 384 с.
Анатов Д.М. Виды рода Delphinium L. в Дагестане и особенности их распространения: Тез. докл. межд. науч. конф. “Изучение флоры Кавказа”. Пятигорск: РИА-КМВ, 2010. С. 13–14.
Андреева Е.Н., Баккал Ю.И., Горшков В.В., Лянгузова И.В., Мазная Е.А., Нешатаев В.Ю., Нешатаева В.Ю., Ставрова И.Н., Ярмишко В.Т., Ярмишко М.А. Методы изучения лесных сообществ. СПб.: НИИхимии. СПбГУ, 2002. 240 с.
Блинова И.В. Эколого-ценотические и онтогенетические стратегии редких видов сосудистых растений Мурманской области // Вестн. КольскНЦ РАН. 2014. Т. 19 (4). С. 83–95.
Вайнагий И.В. О методике изучения семенной продуктивности растений // Ботан. журн. 1974. Т. 59. № 6. С. 826–831.
Галушко А.И. Род Delphinium L. // Флора Северного Кавказа. Определитель. Т. 1. Ростов н/Д.: Изд-во Рост. ун-та, 1978. С. 274–276.
Гроссгейм А.А. Род Delphinium L. // Флора Кавказа. Т. 4. М.; Л.: Изд-во АН СССР, 1950. С. 24–33.
Димитрова В.Н. Новые виды живокости из Дагестана // Новости систематики высших растений. М.; Л.: Наука, 1966. С. 94–107.
Димитрова В.Н. Oligophyllon – новый подрод рода Delphinium L. // Новости систематики высших растений. 1969. Вып. 6. С. 64–66.
Димитрова В.Н. Обзор кавказских видов рода Delphinium L. // Тр. ДагРБО. 2013. Вып. 2. С. 67–70.
Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. № 1. С. 3–7.
Злобин Ю.А. Принципы и методы изучения ценотических популяций растений. Казань: Изд-во КГУ, 1989. 148 с.
Злобин Ю.А. Популяционная экология растений: современное состояние, точки роста: Монография. Сумы: Универ. книга, 2009. 263 с.
Лакин Г.Ф. Биометрия. М.: Высш. шк., 1980. 293 с.
Литвинская С.А., Муртазалиев Р.А. Кавказский элемент во флоре Северного Кавказа: география, созология, экология. Краснодар: Просвещение–Юг, 2009. С. 246–252.
Магомедова М.А., Гасанова О.О. Анализ флористических комплексов в ландшафтах Талгинского ущелья аридных предгорий Дагестана // Естеств. и матем. науки в соврем. мире. 2014. № 15. С. 154–160.
Малютин Н.И. Дельфиниумы. М.: Агропромиздат, 1992. 56 с.
Мамаев С.А. Основные принципы методики исследования внутривидовой изменчивости древесных растений // Тр. Ин-та экологии растений и животных УНЦ АН СССР. 1975. Вып. 94. С. 3–14.
Марков М.Н. Популяционная биология растений. М.: КМК, 2012. 388 с.
Муртазалиев Р.А. Карта флористических районов Дагестана: Матер. VI межданар. конф. “Биологическое разнообразие Кавказа”. Нальчик: КБГУ, 2004. С. 187–188.
Муртазалиев Р.А. Конспект флоры Дагестана. Т. 1. Махачкала: Эпоха, 2009. 356 с.
Онтогенетический атлас лекарственных растений: уч. пособие. Йошкар-Ола: МарГУ, 2000. 268 с.
Плотников В.В. Эволюция структуры растительных сообществ. М.: Наука, 1979. 276 с.
Проханов Я.И. Новые виды из Северо-Восточного Дагестана // Ботан. материалы гербария БИН АН СССР. 1961. № 21. С. 417–437.
Работнов Т.А. Жизненные циклы многолетних травянистых растений в луговых ценозах // Тр. Ботан. ин-та АН СССР. М.; Л.: 1950. Сер. 3. Вып. 6. С. 5–204.
Работнов Т.А. Изучение ценотических популяций в целях выяснения “стратегии жизни” видов растений // Бюл. МОИП. 1975. Т. 80. Вып. 2. С. 5–17.
Работнов Т.А. О типах стратегий растений // Экология. 1985. № 3. С. 3–12.
Ростова Н.С. Корреляции: структура и изменчивость. СПб.: Изд-во СПбГУ, 2002. 308 с.
Тахтаджян А.Л. Редкие и исчезающие виды флоры СССР. Л.: Наука, 1981. 261 с.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергии волновых процессов // Биол. науки. 1975. № 2. С. 7–34.
Ходачек Е.А. Семенная продуктивность и урожай семян растений в тундрах Западного Таймыра // Ботан. журн. 1970. Т. 55. № 7. С. 995– 1010.
Ходачек Е.А. Особенности репродукции цветковых растений Арктики и их репродуктивные стратегии: Матер. ХII Съезда РБО “Фундаментальные и прикладные проблемы ботаники в начале XXI века”. Ч. 1. Петрозаводск: КНЦ РАН, 2008. С. 300–302.
Ценопопуляции растений (основные понятия и структура). М.: Наука, 1976. 217 с.
Ценопопуляции растений. Развитие и взаимоотношения. М.: Наука, 1977. 134 с.
Ценопопуляции растений (очерки популяционной биологии). М.: Наука, 1988. 184 с.
Greig-Smith P. The use of pattern analysis in ecological investigations // Recent Advances in Botany. Toronto: Univ. Press, 1961. P. 1354–1358.
Grime J.P. Vegetation classification by reference to strategies // Nature (London). 1974. V. 250. P. 26–31.
Harper J.L. Population biology of plants. London; New York: Acad. Press, 1977. 892 p.
Wang Wencai, Michael J. Warnock. Delphinium L. Flora of China. 2001. V. 6. P. 223–274.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая