

Известия РАН. Серия биологическая, 2020, № 4, стр. 361-374
Полиморфизм устьиц листьев яблони Malus domestica Borkh. в горах и на равнине
А. В. Бабоша 1, *, Т. Х. Кумахова 2, А. С. Рябченко 1, Г. И. Комарова 1
1 Главный ботанический сад им. Н.В. Цицина Российской академии наук
127276 Москва, ул. Ботаническая, 4, Россия
2 Российский государственный аграрный университет – МСХА им. К.А. Тимирязева
127550 Москва, Тимирязевская ул., 49, Россия
* E-mail: phimmunitet@yandex.ru
Поступила в редакцию 31.07.2018
После доработки 11.10.2018
Принята к публикации 19.10.2018
Аннотация
Методом сканирующей электронной микроскопии исследован полиморфизм устьиц листьев яблонь, произрастающих в горах и на равнине. Листья яблони гипостоматного типа с аномоцитными устьицами и радиально расходящимися кутикулярными складками, объединенными в пучки – кутикулярные тяжи, число которых, как правило, соответствовало количеству окружающих устьица эпидермальных клеток. Установлено, что средние размеры устьиц линейно возрастали с увеличением числа кутикулярных тяжей, в более благоприятных условиях (600 м над уровнем моря) наблюдали высокую плотность устьиц более округлой формы. Отмечено, что стрессовые условия – от высоких температур и засухи на равнине (300 м) до высокой инсоляции и перепада температур в горах (1200 м) – оказывали одинаковое воздействие на микроморфологию поверхности листьев, а влияние высотного фактора проявлялось в уменьшении размеров эпидермальных клеток.
Состояние устьичного аппарата существенно влияет на продуктивность и рост растений, а также играет важную роль в адаптации к условиям внешней среды (Hetherington, Woodward, 2003; Mizutani, Kanaoka, 2017). Проблема включает в себя как физиологический аспект, связанный с хорошо известной оперативной регуляцией апертуры замыкающих клеток, так и морфологический – изменение количества (плотности распределения устьиц в расчете на единицу поверхности листа или число клеток), размеров и формы устьиц у растений, выращиваемых в различных условиях. Последнее обусловливает попытки использования параметров морфологии устьичного аппарата для экологического мониторинга.
Вполне ожидаемо, что параметры устьичного аппарата изменяются в условиях различного обеспечения влагой (Василевская, 1954). Удалось также зафиксировать уменьшение размеров устьиц и увеличение плотности их распределения в холодном климате (Stenström et al., 2002). Увеличение интенсивности освещения при выращивании растений Sinapis alba приводило к удвоению числа устьиц, но вызывало только небольшие изменения их размеров (Wild, Wolf, 1980). У ряда древесных пород средние размеры устьиц уменьшались по мере увеличения степени антропогенного загрязнения (Сунцова, Иншаков, 2007).
Наиболее интересные данные касаются влияния содержания углекислого газа (СО2). При выращивании растений в условиях высокого содержания СО2 размеры устьиц возрастают, а их число уменьшается (Franks, Beerling, 2009). В среднем удвоение концентрации СО2 сопровождается уменьшением плотности распределения устьиц на 11% (Hetherington, Woodward, 2003) и в большей степени зависит от концентрации СО2 в тканях растения, чем от его содержания в атмосфере (Šantrůček et al., 2014). Высокая концентрация CO2, свойственная некоторым историческим эпохам, в большой выборке видов растений коррелировала с уменьшением плотности устьиц на поверхности листьев (Royer, 2001; Franks, Beerling, 2009).
Другая интересная закономерность – достаточно стабильно наблюдаемая обратная корреляция между размерами устьиц и их числом (Blanke et al., 1994; Franks et al., 2009). Увеличение плотности распределения более мелких устьиц – один из способов увеличения устьичной проводимости и адаптации к “CO2-голоданию” при его низких концентрациях в атмосфере (Franks, Beerling, 2009). Подобная обратная корреляция между размерами и плотностью распределения устьиц решает проблему максимизации устьичной проводимости за счет наиболее плотной их упаковки на поверхности листа с учетом поддержания нужной механической прочности всей конструкции (Franks, Farquhar, 2007).
Использование размеров устьиц в качестве количественного параметра для экологического мониторинга некоторых растений может быть затруднено вследствие их гетерогенности по размерам, возрасту, физиологии и морфологии. Так, в листьях Populus tremula обнаружены устьица сразу трех типов: парацитные, латероцитные и промежуточные между аномоцитными и латероцитными (Паутов, 2009а). В листьях Exbucklandia populnea наблюдали парацитные, латероцитные, энциклоцитные и переходные типы устьиц (Паутов и др., 2015). По мнению Паутова (Паутов, 2009б), гетеростоматность может возникать при изменении в ходе развития листа согласованности в росте замыкающих и околоустьичных клеток, что приводит к закономерной смене преобладающих устьичных типов.
Однако и среди устьиц одного и того же типа можно обнаружить более или менее различающиеся размерные группы. Так, наблюдали более крупные и более мелкие устьица в листьях персика (Ma et al., 2008). Соотношение тех и других изменялось в зависимости от сорта. Наличие устьиц разных размерных групп, которые встречаются одновременно на одних и тех же участках листа, наблюдали у ряда видов рода Malus (Ganeva, Uzunova, 2010), включая некоторые сорта яблони (Warabieda et al., 1997).
Различия между устьицами по размерам вряд ли обусловлены только топологическим фактором, возможностью их наиболее плотного расположения на поверхности листа. Устьица могут различаться функционально. Показано, что более мелкие устьица быстрее закрываются при потере влаги листом по сравнению с более крупными (Giday et al., 2013), т.е. они различаются по чувствительности к изменениям условий внешней среды.
Очевидно, что в выборках устьиц имеются и другие источники гетерогенности, обусловленные неоднородностью поверхности. Так, в листьях яблони наблюдали два типа околоустьичной кутикулярной складчатости: одни складки радиально расходились в разные стороны, другие опоясывали замыкающие клетки устьиц, образуя перистоматические кольца (Ganeva, Uzunova, 2010). Аналогичные структуры в районе устьичных комплексов описаны на листьях Hydrangea macrophylla (Паутов, 2011). По данным этого исследователя, длина радиальных складок коррелировала с размерами, а их число – с шириной побочных клеток. Так как характер складчатости сильно различался в окрестностях разных устьиц одного листа, был сделан вывод, что на организацию данного микрорельефа влияла ширина побочных клеток (Паутов, 2011).
Цель работы – установить влияние условий выращивания в горах и на равнине на микроморфологию поверхности и полиморфизм устьиц листьев яблони.
МАТЕРИАЛЫ И МЕТОДЫ
Культурная яблоня обладает высоким уровнем экологической пластичности, что позволяет выращивание этой культуры в разных эколого-географических зонах России. На Северном Кавказе многие сорта яблони культивируются на высотах до 1200–1750 м над уровнем моря (у. м.) в условиях резких суточных перепадов температуры, высокой инсоляции и разреженности атмосферы (Шидаков, 1991; Шидаков, Шидакова, 2006). В горной зоне Кабардино-Балкарии яблоневые насаждения часто расположены в межсклонных долинах и на пологих южных склонах, что создает также специфические микроклиматические условия.
Были исследованы листья яблони Malus domestica сорта Ранет Симиренко с укороченных побегов средней части кроны деревьев, произрастающих в Кабардино-Балкарии на разных высотах: 300 (степная зона), 600 (лесогорная зона), 1200 и 1500 м над у. м. (горная зона). Сбор материала проводили в 2012, 2014 и 2016 гг.
Степная (равнинная) экологическая зона расположена на высоте 200–300 м над у. м. Климат в этой зоне жаркий, засушливый с неустойчивым увлажнением. Здесь часто наблюдаются длительные суховеи (за вегетационный период – 70 сут).
Горная экологическая зона подразделяется на две подзоны: лесогорную (550–800 м) и горно-степную (900–1500 м). Лесогорная подзона в основном теплая, степень увлажнения высокая, наблюдаются кратковременные суховеи (10–15 сут за вегетационный период), горно-степная – умеренно прохладная, среднесуточная температура воздуха в июле 15.6°С. Подзона характеризуется множеством ясных солнечных дней и высокой инсоляцией. Отмечаются кратковременные суховеи (8 сут за вегетационный период).
Фрагменты листьев (1 см2) вырезали между жилками из средней части листовых пластинок и просматривали в режиме высокого вакуума при –30°С с помощью сканирующего электронного микроскопа LEO-1430VP (Carl Zeiss, Германия), оборудованного замораживающей приставкой “Deben CoolStage” (криоСЭМ).
На 10–30 микрографиях каждого образца в программе Image J измеряли в мкм длину и ширину 100–500 устьиц, среднюю площадь основных клеток адаксиального и абаксиального эпидермиса, а также учитывали число устьиц и число кутикулярных тяжей, радиально расходящихся в стороны от каждой устьицы пучков складок периклинальных стенок соседних клеток. Рассчитывали плотность устьиц на единицу поверхности листа. О достоверности различий судили по результатам применения дисперсионного анализа и t-критерия. В таблицах приведены средние значения и их ошибки.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Независимо от высоты произрастания (300–1500 м над у. м.) листьев яблони сорта Ренет Симиренко гипостоматного типа многочисленные устьица расположены на абаксиальной (нижней) стороне. Устьица относятся к аномоцитному стоматотипу, в котором соседние с замыкающими эпидермальные клетки по своей форме практически не отличаются (рис. 1).
Рис. 1.
Изменение морфологии устьиц яблони у деревьев в зависимости от высоты произрастания над у. м. (криоСЭМ). а – устьица с хорошо выраженными кутикулярными тяжами и перистоматическими кольцами (600 м, 2016 г.). б–е – типичные устьица растений, произрастающих на высотах: б –1500 (2014 г.), в – 300 (2016 г.), г – 1200 (2016 г.), д – 600 (2016 г.), е – 600 м 2012 г.). Масштаб: 10 мкм.
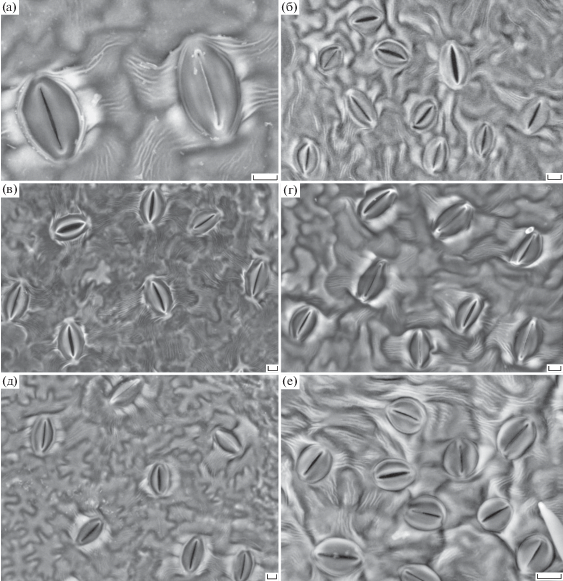
Адаксиальная эпидерма состоит из однотипных плотно расположенных клеток таблитчатой или слегка многоугольной формы, с почти прямыми антиклинальными стенками. Их поверхность покрыта кутикулой в виде длинных прядей и бороздок, которые часто простираются поверх границы нескольких эпидермальных клеток.
Абаксиальная эпидерма в отличие от адаксиальной представлена сложной тканью. В ее состав входят основные эпидермальные и устьичные клетки, а также трихомы (волоски).
В табл. 1 и 2 представлены результаты измерения площади основных эпидермальных клеток абаксиальной и адаксиальной сторон листа. В данных 2012 и 2016 гг. выявляется достаточно отчетливая тенденция уменьшения площади эпидермальных клеток по мере увеличения высоты произрастания растений над у. м., однако сравнение выборок крайних вариантов высоты (300 и 1200 м) в наблюдениях 2016 г. не показало достоверных различий. При этом в 2014 г. сравнение вариантов 300 и 1500 м имеет обратную тенденцию. В обоих исследованиях 2012 и 2016 гг. в варианте высоты 300 м наблюдали более высокие значения площади клеток абаксиальной эпидермы по сравнению со сходными значениями вариантов высот 600 и 1200 м. Для клеток адаксиальной эпидермы вариант высоты 1200 м в те же годы наблюдения имел более низкие значения по сравнению с клетками растений на высотах 300 и 600 м. Плотность устьиц как в 2012, так и в 2016 г. достигала наибольших значений на листьях яблони, собранных на высоте 600 м над у. м. (различия вариантов высот 600 и 300 м достоверны (p = 0.04), в дисперсионном анализе влияние фактора вариантов достоверно при p = 0.04).
Таблица 1.
Размеры клеток адаксиальной и абаксиальной эпидермы и плотность устьиц листьев яблони в горных и равнинных условиях
| Год наблюдений | Высота, м над у.м. | Площадь клетки, мкм2 | Число устьиц/мм2 | Доля первичных устьиц |
|
|---|---|---|---|---|---|
| абаксиальная сторона | адаксиальная сторона | ||||
| 2012 | 300 | 406 ± 25 | 486 ± 21 | 216 ± 18 | 0.038 ± 0.003 |
| 2012 | 600 | 332 ± 24 | 466 ± 18 | 257 ± 76 | 0.022 ± 0.002 |
| 2012 | 1200 | 323 ± 11 | 403 ± 8 | 218 ± 11 | 0.054 ± 0.006 |
| 2014 | 300 | 268 ± 17 | 350 ± 11 | 306 ± 21 | 0.029 ± 0.010 |
| 2014 | 1500 | 371 ± 8 | 499 ± 27 | 311 ± 23 | 0.055 ± 0.012 |
Таблица 2.
Варьирование средних значений параметров устьиц и эпидермальных клеток у листьев, собранных с трех деревьев (повторности) в горных и равнинных условиях в 2016 г.
| Параметр | Высота, м над у.м. | Повторности | Суммарная выборка* | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| Площадь, мкм2, эпидермальной клетки на абаксиальной стороне | 300 | 338 ± 15 | 372 ± 9 | 476 ± 50 | 396 ± 21(42) |
| 600 | 370 ± 23 | 367 ± 20 | 366 ± 30 | 368 ± 13(1) | |
| 1200 | 343 ± 16 | 356 ± 27 | 376 ± 17 | 356 ± 11(10) | |
| Число устьиц/мм | 300 | 286 ± 11 | 232 ± 12 | 215 ± 10 | 242 ± 9(22) |
| 600 | 324 ± 10 | 356 ± 20 | 437 ± 18 | 361 ± 15(33) | |
| 1200 | 270 ± 18 | 286 ± 21 | 254 ± 14 | 268 ± 11(9) | |
| Доля первичных устьиц | 300 | 0.076 ± 0.015 | 0.099 ± 0.019 | 0.077 ± 0.017 | 0.083 ± 0.010(0.005) |
| 600 | 0.077 ± 0.016 | 0.053 ± 0.011 | 0.042 ± 0.010 | 0.055 ± 0.007(0.007) | |
| 1200 | 0.096 ± 0.014 | 0.089 ± 0.016 | 0.077 ± 0.016 | 0.089 ± 0.009(0.004) | |
| Площадь, мкм2, эпидермальной клетки на адаксиальной стороне | 300 | 364 ± 29 | 379 ± 6 | 557 ± 0.3 | 433 ± 40(44) |
| 600 | 451 ± 23 | 447 ± 10 | 378 ± 31 | 426 ± 18(17) | |
| 1200 | 376 ± 22 | 291 ± 32 | 454 ± 19 | 374 ± 25(26) | |
Примечание. * – в скобках приведены ошибки, при расчете которых в качестве повторности использовали средние значения для каждого из деревьев; для табл. 2–4 и 6.
Особенность клеток, окружающих устьица, – наличие закономерно расположенных складок, собранных в пучки (кутикулярные тяжи) от 2 до 7 возле каждой устьицы (рис. 1). Кутикулярные тяжи в отличие от границ клеток эпидермы хорошо видны на электронных микрографиях. У отдельных устьиц на рис. 1 видно, что на каждой соседней клетке располагается не более 1 тяжа, т.е. число тяжей примерно соответствует числу соседних клеток.
По морфологии и размерам можно выделить два основных типа устьиц: первичные (более ранние) и вторичные (более поздние). Первичные относительно редкие, в большинстве случаев очень крупные, обычно они расположены в центре группы устьиц средних размеров (рис. 1б). Вторичные устьица, наоборот, составляли бόльшую часть выборки (рис. 1). Аналогичное разделение устьиц по времени формирования было описано ранее (Паутов, 2009а; Паутов и др., 2014). Также в эпидерме наблюдали немного очень мелких устьиц, которые находились в ювенильной стадии либо были недоразвиты.
Соотношения первичных и вторичных устьиц различались в разных вариантах (300, 600, 1200 и 1500 м), а также варьировали в разные годы наблюдений (табл. 1 и 2). Доля первичных устьиц во все годы наблюдений была существенно меньше у листьев, собранных на высоте 600 м над у. м. Кроме того, в 2012 и 2014 гг. она была заметно больше у плодовых растений, выращенных в горных условиях на высотах 1200 и 1500 м над у. м., по сравнению с вариантом 300 м. При этом в 2016 г. это различие было отмечено между высотами 300 и 1200 м, но было относительно небольшим.
В табл. 3 и 4 представлены средние значения длины и ширины устьиц. При анализе трехлетних данных отчетливо видна тенденция увеличения длины вторичных устьиц с увеличением высоты произрастания деревьев над у. м. Аналогичная тенденция проявилась и для ширины устьиц в 2012 и 2014 гг. Средняя длина первичных устьиц в 2016 г. на всех высотах была одинакова.
Таблица 3.
Средние значения длины (Д) и ширины (Ш) устьиц листьев яблони в горных и равнинных условиях
| Год наблюдения | Высота, м над у.м. | Д, мкм | Ш, мкм | Д/Ш | Повторность* |
|---|---|---|---|---|---|
| 2012 | 300 | 20.3 ± 0.2 | 14.9 ± 0.1 | 1.38 ± 0.01 | 288 |
| 2012 | 600 | 19.5 ± 0.2 | 15.5 ± 0.2 | 1.27 ± 0.01 | 117 |
| 2012 | 1200 | 23.6 ± 0.1 | 15.9 ± 0.1 | 1.52 ± 0.01 | 489 |
| 2014 | 300 | 19.5 ± 0.2 | 14.4 ± 0.2 | 1.38 ± 0.02 | 168 |
| 2014 | 1500** | 22.9 ± 0.2 (1.4) |
15.1 ± 0.2 (1.2) |
1.56 ± 0.02 (0.06) |
295 |
Таблица 4.
Варьирование средних значений длины (Д, мкм) и ширины (Ш, мкм) устьиц листьев яблони, собранных с трех деревьев (повторности) в горных и равнинных условиях в 2016 г.
| Параметр | Высота, м над у.м. | Повторности | Суммарная выборка* | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| Первичные устьица | |||||
| Число устьиц | 300 | 24 | 25 | 17 | 66 |
| 600 | 22 | 19 | 12 | 53 | |
| 1200 | 29 | 26 | 18 | 73 | |
| Д | 300 | 29.2 ± 0.5 | 27.7 ± 0.5 | 28.6 ± 0.9 | 28.5 ± 0.4(0.4) |
| Д | 600 | 28.2 ± 0.8 | 30.1 ± 0.9 | 26.4 ± 0.8 | 28.5 ± 0.5(1.1) |
| Д | 1200 | 29.1 ± 0.4 | 27.6 ± 0.5 | 28.8 ± 0.6 | 28.5 ± 0.3(0.4) |
| Ш | 300 | 16.8 ± 0.4 | 16.4 ± 0.3 | 17.4 ± 0.3 | 16.8 ± 0.2(0.3) |
| Ш | 600 | 17.0 ± 0.3 | 15.9 ± 0.4 | 17.4 ± 0.7 | 16.7 ± 0.3(0.5) |
| Ш | 1200 | 15.7 ± 0.4 | 15.9 ± 0.4 | 16.1 ± 0.4 | 15.9 ± 0.2(0.1) |
| Д/Ш | 300 | 1.75 ± 0.05 | 1.67 ± 0.04 | 1.62 ± 0.05 | 1.69 ± 0.03(0.04) |
| Д/Ш | 600 | 1.45 ± 0.09 | 1.85 ± 0.03 | 1.45 ± 0.04 | 1.57 ± 0.05(0.13) |
| Д/Ш | 1200 | 1.94 ± 0.05 | 1.70 ± 0.04 | 1.77 ± 0.04 | 1.80 ± 0.03(0.07) |
| Вторичные устьица | |||||
| Число устьиц | 300 | 193 | 129 | 150 | 472 |
| 600 | 165 | 177 | 126 | 468 | |
| 1200 | 119 | 104 | 84 | 307 | |
| Д | 300 | 22.8 ± 0.2 | 22.6 ± 0.2 | 24.2 ± 0.2 | 23.2 ± 0.1(0.5) |
| Д | 600 | 24.0 ± 0.2 | 24.7 ± 0.2 | 21.7 ± 0.2 | 23.7 ± 0.1(0.9) |
| Д | 1200 | 24.9 ± 0.3 | 23.5 ± 0.3 | 25.0 ± 0.3 | 24.4 ± 0.2(0.5) |
| Ш | 300 | 14.3 ± 0.1 | 14.2 ± 0.1 | 14.5 ± 0.1 | 14.3 ± 0.1(0.1) |
| Ш | 600 | 14.8 ± 0.1 | 14.3 ± 0.1 | 15.4 ± 0.2 | 14.7 ± 0.1(0.3) |
| Ш | 1200 | 13.5 ± 0.2 | 13.5 ± 0.2 | 14.3 ± 0.2 | 13.7 ± 0.1(0.3) |
| Д/Ш | 300 | 1.52 ± 0.01 | 1.60 ± 0.02 | 1.64 ± 0.02 | 1.59 ± 0.01(0.04) |
| Д/Ш | 600 | 1.57 ± 0.02 | 1.62 ± 0.02 | 1.39 ± 0.02 | 1.54 ± 0.01(0.07) |
| Д/Ш | 1200 | 1.87 ± 0.03 | 1.78 ± 0.03 | 1.70 ± 0.02 | 1.78 ± 0.02(0.05) |
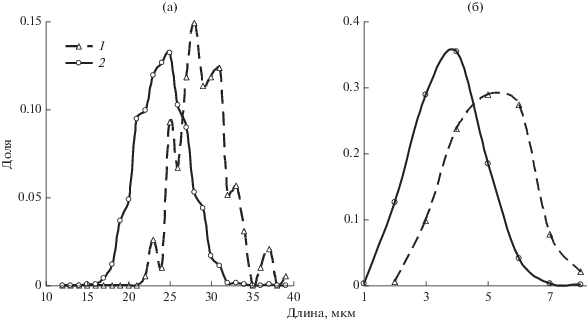
Средние размеры первичных и вторичных устьиц сильно различались (табл. 4, рис. 2а). Заметно различаются и распределения устьиц этих групп по числу тяжей (рис. 2б). Аналогично выглядят соответствующие графики распределений обеих групп устьиц по ширине (данные не представлены). Длина и ширина первичных устьиц, а также число тяжей в среднем во всех вариантах были существенно выше, чем у вторичных. Тем не менее графики частоты распределений по длине и числу тяжей у обеих групп заметно перекрываются (рис. 2), т.е. имеется большое число устьиц, которые различаются морфологически, но одинаковы по размерам и имеют одинаковое число тяжей. Высокая трансгрессия пиков не позволяет использовать эти параметры в качестве диагностических признаков.
Рис. 2.
Распределение первичных (1) и вторичных (2) устьиц по длине (а) и числу тяжей (б) (сумма вариантов 2016 г.).
На рис. 1 видно, что устьичные клетки на высотах 1200 и 1500 м более вытянуты, а на высоте 600 м имеют более округлую форму. Различия формы устьиц между высотами можно оценить количественно, рассчитав отношение длины к ширине (Д/Ш). Отношение этих параметров у растений, произрастающих на большой высоте, во все годы наблюдений существенно выше по сравнению с другими вариантами (табл. 3 и 4). Наоборот, в варианте 600 м оно ближе к единице, что особенно четко проявилось в 2012 г. Данные 2016 г. (табл. 4) свидетельствуют, что указанные закономерности примерно в одинаковой степени проявляются как для вторичных, так и для первичных устьиц.
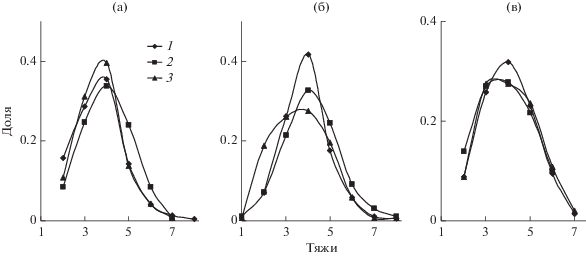
На рис. 3 и 4 представлены графики распределения устьиц по числу околоустьичных тяжей. Полученные данные свидетельствуют о том, что соотношение устьиц с различным числом тяжей, а значит и с различным числом соседних клеток, достаточно лабильно. В 2012 и 2014 гг. среди устьиц у растений, которые произрастают на высоте 1200 м, преобладало относительно большое число тяжей, а распределения в вариантах 300 и особенно 600 м были заметно сдвинуты в сторону устьиц с небольшим их числом (рис. 3). Наоборот, в 2016 г. во всех вариантах были наиболее многочисленны устьица с четырьмя тяжами. Очень близки между собой и распределения, соответствующие отдельным деревьям (рис. 4).
Рис. 3.
Распределение устьиц на листьях яблони, произрастающих в равнинных и горных условиях по числу кутикулярных тяжей (средние данные по вариантам 300 (1), 600 (2), 1200 (3) и 1650 (4) м над у. м. в 2012 (а), 2014 (б) и 2016 гг. (в)).
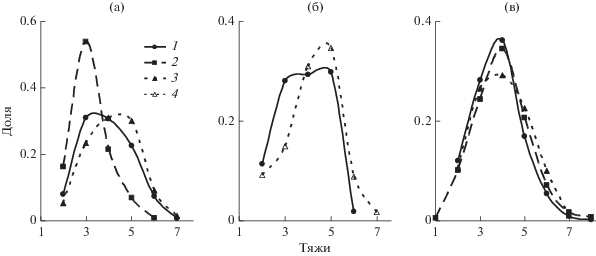
Рис. 4.
Варьирование формы кривой распределения устьиц по числу тяжей у разных деревьев (1–3) в равнинных и горных условиях (варианты 300 (а), 600 (б) и 1200 (в) м над у. м., 2016 г.).
Как свидетельствуют данные рис. 2, первичные устьица имеют большую длину и одновременно в среднем имеют большее число тяжей по сравнению с вторичными. Между числом околоустьичных тяжей и размерами устьиц имеется заметная корреляция, поэтому зависимость между этими показателями была исследована более подробно. На рис. 5 представлены графики зависимости средней длины первичных и вторичных устьиц в 2016 г. от числа тяжей. Во всех вариантах устьица с большим числом тяжей в среднем имели и большую длину. Сходные графики можно построить и для зависимости от числа тяжей средней ширины устьиц. Расположение экспериментальных точек позволяет предположить, что зависимость в обоих случаях близка к линейной, а отклонения носят случайный характер. При этом большей повторности у вторичных устьиц по сравнению с первичными соответствовало и более близкое к прямой линии расположение точек. Поэтому были рассчитаны параметры a и b линии регрессии Y = b × X + a (Y – длина или ширина устьиц, X – число кутикулярных тяжей) и ее доверительный интервал. Поскольку количество устьиц в группах с разным числом тяжей сильно различалось (рис. 5), при построении регрессии и вычислении ее параметров использовали весовой метод (Себер, 1980). В качестве весов использовали повторность, с которой были получены средние значения длины или ширины для устьиц с определенным числом тяжей.
Рис. 5.
Зависимость длины устьиц от числа тяжей для первичных (а–в) и вторичных (г–е) устьиц на листьях яблони в горных и равнинных условиях. Регрессия (Y = b × X + a) и ее доверительный интервал (p = 0.05) – 1 и 2 соответственно (суммарные данные трех деревьев в 2016 г.). 3 – средние значения длины устьиц с определенным числом тяжей. Параметры линии регрессии рассчитаны весовым методом с использованием числа устьиц для каждой градации числа тяжей в качестве веса. а, г – 300; б, д – 600; в, е – 1200 м над у. м.
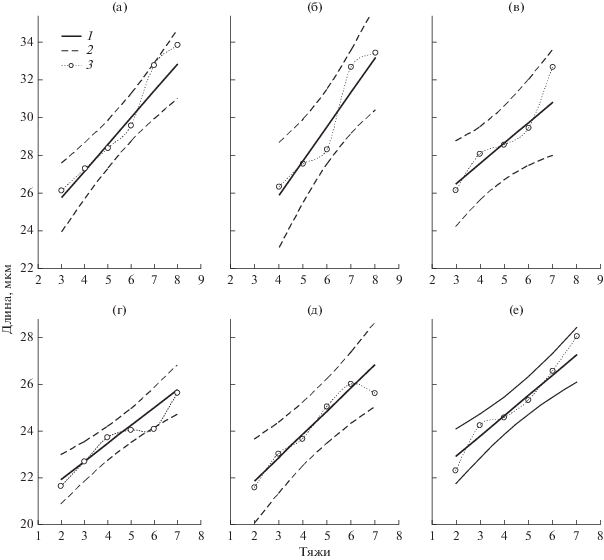
В табл. 5 и 6 приведены значения параметров линейной регрессии зависимости длины и ширины устьиц от числа кутикулярных тяжей. Судя по значениям b (коэффициент наклона кривой), увеличение числа тяжей на 1 приводило к возрастанию длины устьица на 0.6–2 мкм и ее ширины на 0.1–0.6 мкм. Коэффициенты b для длины достигали достоверных значений у обоих типов устьиц почти для всех деревьев. Это свидетельствует о том, что зависимость длины устьиц от числа тяжей действительно существует. Во всех вариантах значения b первичных устьиц превышали значения вторичных, а в вариантах высот 300 и 600 м это превышение было достоверным (р = 0.021 и 0.048 соответственно). В наблюдениях 2012 и 2016 гг., в которых были исследованы растения, находящиеся на высоте 600 м над у. м., в этом варианте отмечена тенденция увеличения коэффициента b. Однако в целом достоверных различий между высотами не было, т.е. линии регрессии в разных вариантах почти параллельны одна другой. Вероятно, невозможность их статистического различения обусловлена высокими отклонениями от средних значений у отдельных деревьев из одной и той же высоты. Вариабельность коэффициента b для ширины устьиц у разных деревьев была еще больше. Ни варианты, ни группы устьиц достоверных различий не показали, а сами значения b для ширины устьиц были достоверными лишь в отдельных случаях (это свидетельствует о слабой зависимости ширины устьиц от числа тяжей). Отношение Д/Ш от числа тяжей не зависело.
Таблица 5.
Параметры a и b линии регрессии Y = b × X + a (Y – длина или ширина устьиц, X – число кутикулярных тяжей) у листьев яблони в горных и равнинных условиях
| Год наблюдения | Высота, м над у.м. | Длина | Ширина | ||
|---|---|---|---|---|---|
| b | a | b | a | ||
| 2012 | 300 | 0.73 ± 0.13 | 17.5 ± 1.0 | 0.37 ± 0.07 | 13.5 ± 0.5 |
| 2012 | 600 | 1.06 ± 0.10 | 16.1 ± 0.4 | 0.22 ± 0.12 | 14.9 ± 0.5 |
| 2012 | 1200 | 0.95 ± 0.11 | 19.6 ± 1.1 | 0.18 ± 0.16 | 15.1 ± 1.6 |
| 2014 | 300 | 1.20 ± 0.17 | 15.0 ± 1.1 | 0.65 ± 0.08 | 11.9 ± 0.5 |
| 2014 | 1500* | 1.25 ± 0.13 | 17.7 ± 1.1 | 0.77 ± 0.16 | 11.8 ± 1.2 |
Таблица 6.
Варьирование параметров a и b линии регрессии Y = b × X + a (Y – длина или ширина устьиц, X – число кутикулярных тяжей) устьиц листьев яблони, собранных с трех деревьев (повторности) в горных и равнинных условиях в 2016 г.
| Параметр | Высота, м над у.м. | Повторности | Суммарная выборка | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| Первичные устьица | |||||
| Длина (a) | 300 | 22.6 ± 1.0 | 21.4 ± 0.7 | 20.8 ± 0.6 | 21.5 ± 0.7(0.5) |
| Длина (a) | 600 | 17.4 ± 0.7 | 20.5 ± 1.5 | 18.5 ± 0.7 | 18.6 ± 1.6(0.9) |
| Длина (a) | 1200 | 25.9 ± 1.1 | 22.9 ± 1.2 | 20.1 ± 0.7 | 23.4 ± 1.2(1.7) |
| Длина (b) | 300 | 1.30 ± 0.39 | 1.32 ± 0.27 | 1.60 ± 0.33 | 1.41 ± 0.20(0.10) |
| Длина (b) | 600 | 2.16 ± 0.29 | 1.65 ± 0.64 | 1.43 ± 0.38 | 1.83 ± 0.40(0.22) |
| Длина (b) | 1200 | 0.63 ± 0.41 | 1.04 ± 0.38 | 1.72 ± 0.31 | 1.06 ± 0.31(0.32) |
| Ширина (a) | 300 | 13.6 ± 0.8 | 15.9 ± 0.6 | 15.6 ± 0.6 | 14.8 ± 1.1(0.7) |
| Ширина (a) | 600 | 15.7 ± 0.7 | 13.9 ± 0.2 | 14.1 ± 0.6 | 15.5 ± 0.5(0.6) |
| Ширина (a) | 1200 | 15.1 ± 0.4 | 15.9 ± 0.6 | 15.6 ± 0.6 | 15.5 ± 0.9(0.2) |
| Ширина (b) | 300 | 0.62 ± 0.31 | 0.11 ± 0.26 | 0.37 ± 0.32 | 0.40 ± 0.29(0.15) |
| Ширина (b) | 600 | 0.25 ± 0.28 | 0.34 ± 0.10 | 0.60 ± 0.30 | 0.21 ± 0.13(0.10) |
| Ширина (b) | 1200 | 0.31 ± 0.16 | 0.11 ± 0.26 | 0.37 ± 0.32 | 0.25 ± 0.24(0.08) |
| Вторичные устьица | |||||
| Длина (a) | 300 | 19.2 ± 1.1 | 19.7 ± 0.4 | 22.1 ± 0.7 | 20.7 ± 1.1(0.9) |
| Длина (a) | 600 | 20.6 ± 1.2 | 21.1 ± 0.6 | 19.1 ± 0.7 | 19.8 ± 0.9(0.6) |
| Длина (a) | 1200 | 21.1 ± 1.4 | 20.5 ± 0.9 | 22.4 ± 0.6 | 21.2 ± 1.3(0.6) |
| Длина (b) | 300 | 1.02 ± 0.19 | 0.76 ± 0.08 | 0.57 ± 0.14 | 0.77 ± 0.13(0.13) |
| Длина (b) | 600 | 0.89 ± 0.22 | 0.89 ± 0.11 | 0.76 ± 0.14 | 1.00 ± 0.11(0.04) |
| Длина (b) | 1200 | 1.00 ± 0.28 | 0.83 ± 0.18 | 0.67 ± 0.15 | 0.87 ± 0.16(0.10) |
| Ширина (a) | 300 | 12.2 ± 0.3 | 12.3 ± 1.1 | 13.4 ± 0.8 | 12.7 ± 0.3(0.4) |
| Ширина (a) | 600 | 14.2 ± 0.6 | 13.6 ± 0.5 | 14.7 ± 0.5 | 14.4 ± 0.5(0.3) |
| Ширина (a) | 1200 | 12.5 ± 0.8 | 13.1 ± 0.2 | 13.0 ± 0.9 | 12.9 ± 0.2(0.2) |
| Ширина (b) | 300 | 0.60 ± 0.05 | 0.49 ± 0.20 | 0.29 ± 0.16 | 0.46 ± 0.03(0.09) |
| Ширина (b) | 600 | 0.14 ± 0.10 | 0.16 ± 0.09 | 0.20 ± 0.10 | 0.10 ± 0.07(0.02) |
| Ширина (b) | 1200 | 0.28 ± 0.17 | 0.10 ± 0.04 | 0.32 ± 0.21 | 0.23 ± 0.02(0.07) |
Коэффициент a в уравнении регрессии имеет физический смысл гипотетической длины (ширины) устьиц, не имеющих тяжей. Значение этого коэффициента как для длины, так и ширины меньше средних размеров устьиц, но имеет тот же порядок. Варьирование этого показателя, очевидно, также обусловлено в большей степени различиями биологии отдельных деревьев. Варианты высоты произрастания статистически неразличимы, однако порядок вариантов по значению a (длина устьица) одинаков во все годы наблюдений и для обеих групп устьиц (600 < 300 < 1200 м).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Особенности микрорельефа листьев яблони в данной работе были изучены в трех основных местах произрастания (300, 600 и 1200 м над у. м). В предгорьях Северного Кавказа условия варианта 600 м были наиболее благоприятны по температурному режиму и влажности для культивирования яблони сорта Ренет Симиренко и получения высокого урожая плодов хорошего качества (Шидаков, 1991; Шидаков, Шидакова, 2006). В горной и степной зонах данного региона растения подвергались воздействию комплекса специфических для каждой точки стрессоров. В этих условиях плоды яблони мельчают, что приводит к снижению экономических показателей при культивировании данного сорта. Степные растения варианта 300 м, как правило, страдают от высоких температур и засухи. Наоборот, климат в горной зоне вариантов 1200 и 1500 м более прохладный и влажный. Помимо этого на высоте 1200 м атмосферное давление снижается в среднем на 100 мм рт. ст., содержание кислорода с 21 до 18%, влажность уменьшается до 60%, а ультрафиолетовая (УФ) инсоляция увеличивается более чем в 1.5 раза по сравнению с соответствующими значениями над у. м. моря (Суслов и др., 1999). Высокие дозы УФ-облучения могут приводить к повреждению фотосинтетического аппарата, способствуют возникновению окислительного стресса, вызывают морфологические изменения: уменьшение поверхности и увеличение толщины листьев (Frohnmeyer, Staiger, 2003; Nawkar et al., 2013). С увеличением высоты местообитания над у. м. в листьях ряда луговых растений наблюдали увеличение палисадной и губчатой ткани, абаксиального и адаксиального эпидермиса, что приводило к общему утолщению листа (Чхубианишвили и др., 2009).
У растений разных жизненных форм с увеличением высоты произрастания над у. м. плотность устьиц уменьшалась (Körner et al., 1986; Gou et al., 2005), а их размеры увеличивались (Yang et al., 2014). В других работах в условиях горной местности у разных видов растений было отмечено как увеличение, так и уменьшение числа устьиц (Kofidis et al., 2003; Kessler et al., 2007; Kouwenberg et al., 2007).
Изменения параметров микроморфологии в наших исследованиях в целом аналогичны опубликованным ранее (Elias, 1995; Мотылёва, Кузнецова, 2010). Так, было отмечено наибольшее значение плотности устьиц в варианте выращивания яблони без орошения. Это позволяет предположить, что уменьшение этого параметра с высотой произрастания в приведенных выше работах (Körner et al., 1986; Gou et al., 2005) может быть обусловлено повышенной влажностью в горных условиях. Однако в отличие от данных (Elias, 1995), как свидетельствуют полученные нами данные (табл. 1 и 2), наибольшей в течение 2 лет плотность устьиц была в варианте (высота 600 м) с наиболее благоприятными для яблони условиями, в том числе по обеспеченности влагой. На высотах 300 и 1200 м этот показатель был заметно меньшим, что соответствует преимущественно неспецифическому снижению плотности устьиц в стрессовых условиях.
Уменьшение размеров клеток некоторые исследователи считают одной из важнейших особенностей растений холодного климата, способствующих повышению их устойчивости (Мирославов, Кравкина, 1990; Мирославов, 1994). В наших наблюдениях у яблони проявились достаточно устойчивые тенденции уменьшения площади эпидермальных клеток на адаксиальной и абаксиальной сторонах листовой пластинки с увеличением высоты произрастания плодовых деревьев (табл. 1 и 2). Также происходило заметное удлинение замыкающих клеток вторичных устьиц, которое, однако, либо не было монотонным в 2012 г. (табл. 3), либо было совсем невелико в 2016 г. (табл. 4). При этом средняя длина первичных устьиц в 2016 г. во всех вариантах была одинакова. Высокую вариабельность наблюдали как в разные годы, так и у разных деревьев, произрастающих на одной и той же высоте. Это свидетельствует о том, что в определении характера микроморфологии поверхности листьев влияние высотного фактора, вероятно, не имеет решающего значения и сравнимо с влиянием локальных факторов, в частности специфических условий, которые возникают на склонах и проявление которых мало зависит от высоты над у. м.
Большей стабильностью обладал показатель отношения длины к ширине устьиц (табл. 3 и 4). В более благоприятных для яблони условиях (высота 600 м) устьица имели более округлую форму Д/Ш. Наоборот, листья горных растений (на высоте 1200 м) имели более вытянутые устьица и достоверные отличия параметра Д/Ш (для вторичных устьиц) даже в условиях 2016 г., в котором многие другие параметры всех вариантов были сходными. Более вытянутая форма замыкающих клеток горных растений (на высоте 1200 м) также была отмечена как у вторичных, так и у первичных устьиц. Следует отметить, что в наблюдениях 2012 и 2016 гг. значения Д/Ш на высоте 300 м занимали промежуточное положение. Можно предположить, что значения этого показателя определялись в большей степени суммарным неспецифическим воздействием комплекса стрессоров.
Отделить собственно адаптивные реакции от симптомов повреждения достаточно трудно. На наш взгляд, правдоподобным выглядит предположение, что более округлые устьица (на высоте 600 м) более функциональны. Структурное разнообразие эпидермы может быть обусловлено механическими напряжениями и деформациями в ювенильной стадии (при делении и растяжении клеток). В этом случае удлиненные устьица двух других вариантов возникают в результате достаточно резких нарушений нормального развития листа. Вместе с тем нельзя исключить, что в условиях засухи или разреженного горного воздуха большое число нормально функционирующих устьиц могло бы приводить к чрезмерным потерям воды.
Вычисление средних значений параметров не вызывает трудностей при сравнении относительно однородных выборок. Наши данные свидетельствуют о том, что в случае устьиц растений яблони эти выборки не однородны и могут быть разделены на несколько групп. Во-первых, даже визуально можно выделить некоторое число устьиц, находящихся, как правило, в центре группы других устьиц и названных поэтому первичными. В отличие от остальных (вторичных) устьиц первичные устьица более крупные, имеют в целом больше тяжей, а увеличение их размеров с увеличением числа тяжей происходит быстрее. Разделение устьиц на первичные и вторичные, вероятно, соответствует размерным группам устьиц у яблони, а также у некоторых других древесных культур, которые наблюдали ранее (Warabieda et al., 1997; Ganeva, Uzunova, 2010). Во-вторых, устьица с разным числом кутикулярных тяжей также имеют свои особенности, в частности средние размеры устьиц возрастают при увеличении числа тяжей с зависимостью, близкой к линейной (рис. 5).
Очевидно, что увеличение доли вторичных устьиц, а также доли устьиц с небольшим числом тяжей и, соответственно, меньшими размерами будет сказываться на их средних значениях. Подобная неоднородность в изучаемых выборках устьиц приводит к тому, что различия или сходство вариантов по средним размерам могут иметь разную природу. В этом случае, вероятно, полезно сравнивать не средние размеры устьиц, а взаимное расположение кривых зависимости длины от числа тяжей или параметры линии регрессии (a и b), которые не зависят от распределения устьиц по числу тяжей.
Как правило, на одной соседней клетке может располагаться не более одного кутикулярного тяжа. По мнению Паутова (Паутов, 2009б), полиморфизм устьиц связан с их разным возрастом: ранние устьица в массиве клеток на стадии деления могут достигнуть необходимых размеров только за счет деления побочных клеток. По мере развития листа и смене фазы преимущественного деления растяжением у более поздних устьиц появляется возможность достигнуть функционального размера за счет растяжения соседних клеток и без увеличения их числа. В этом случае число кутикулярных тяжей, которое коррелирует с числом соседних клеток, может служить своеобразным индикатором возраста или, точнее, времени закладки устьиц. Естественно предположить, что изменение доли устьиц разного типа и с разным числом тяжей (рис. 3) в наших экспериментах связано с изменениями в характере развития листа.
Вариант (высота 600 м) с оптимальными условиями произрастания отличался существенно более низкой долей первичных устьиц, а у горных растений (высота 1200 м) она во все годы наблюдений была наивысшей (табл. 1 и 2). Соотношение устьиц с разным числом тяжей варьировало в разные годы наблюдений. Можно предположить, что в 2016 г. климатические условия на разных высотах были сходными, что обусловило идентичные распределения устьиц по числу тяжей (рис. 3). В 2012 г. стрессовые явления, характерные для степных и горных территорий, проявились в полной мере и обусловили более раннюю закладку устьиц с большим числом кутикулярных тяжей. Наоборот, при благоприятных условиях варианта (высота 600 м) большая часть устьиц возникла на более поздних этапах при наступлении фазы растяжения основных клеток эпидермы.
Таким образом, стрессоры независимо от их природы оказывают заметное и сходное влияние на микроморфологию поверхности листьев яблони. Это проявлялось в отличиях (достоверных или в виде устойчивой тенденции) варианта (высота 600 м) с наиболее благоприятными для яблони условиями от двух других вариантов с меньшей и большей высотами (300 и 1200 м) произрастания по показателям плотности, длины и формы устьиц, а также доли первичных устьиц. Изменения, которые можно достаточно однозначно интерпретировать как результат более или менее специфического воздействия высотного фактора, проявились только в виде тенденции монотонного уменьшения размеров эпидермальных клеток на обеих сторонах листа с увеличением высоты над у. м. Неспецифичность воздействия высотного фактора, вероятно, одна из причин неоднозначных результатов в приведенных выше литературных источниках.
Другой причиной, по крайней мере, у растений с устьицами аномоцитного типа может быть их неоднородность. Нами показано, что средние размеры устьиц яблони зависят от числа тяжей и линейно возрастают с его увеличением. Кроме того, можно выделить хорошо различающиеся первичные и вторичные устьица. Поэтому выборки могут отличаться не только собственно размерами устьиц, но также соотношением подгрупп с различными характеристиками в их составе. В этом случае необходимо применять регрессионный анализ, а также учитывать данные о распределении устьиц по группам. Эти дополнительные параметры, по нашему мнению, могут оказаться информативными при изучении механизмов развития устьичного аппарата и его адаптации к условиям внешней среды.
Список литературы
Василевская В.К. Формирование листа засухоустойчивых растений. Ашхабад: Изд-во АН ТССР, 1954. 183 с.
Мирославов Е.А. Структурные адаптации растений к холодному климату // Бот. журн. 1994. Т. 79. № 2. С. 20–26.
Мирославов Е.А., Кравкина Е.М. Сравнительная анатомия листа растений, произрастающих в горах на разных высотах // Бот. журн. 1990. Т. 75. № 3. С. 368–375.
Мотылёва С.М., Кузнецов М.Н. Микроморфология листьев триплоидных иммунных к парше сортов яблони // Соврем. садоводство – Contemporary horticulture. 2010. № 1. С. 72–78.
Паутов А.А. Закономерности филоморфогенеза вегетативных органов растений. СПб.: Изд-во СПбГУ, 2009а. 219 с.
Паутов А.А. Роль морфогенетических корреляций в возникновении гетеростоматности // Бот. журн. 2009б. Т. 94. № 6. С. 785–792.
Паутов А.А. Расположение складок микрорельефа на побочных клетках устьиц Hydrangea macrophylla (Thunb.) Ser.(Hydrangeaceae) // Вестн. СПбГУ. Сер. 3. Биология. 2011. № 2. С. 39–44.
Паутов А.А., Васильева В.А., Крылова Е.Г. Полимофизм устьиц в эпидерме листа Exbucklandia populnea (Hamamelidaceae) и его возможное значение // Бот. журн. 2015. Т. 100. № 6. С. 540–549.
Паутов А.А., Сапач Ю.О., Иванова О.В., Крылова Е.Г. Микрорельеф поверхности листьев цветковых растений: устьичные кольца и выступы // Бот. журн. 2014. Т. 99. № 6. С. 625–640.
Себер Дж. Линейный регрессионный анализ. М.: Мир, 1980. 456 с.
Сунцова Л.Н., Иншаков Е.М. Древесные растения в условиях техногенной среды города Красноярска // Хвойные бореальной зоны. 2007. Т. 24. № 1. С. 95–99.
Суслов Ф.П., Гиппенрейтер Е.Б., Холодов Ж.К. Спортивная тренировка в условиях среднегорья. М.: РГАФК, 1999. 202 с.
Чхубианишвили Е.И., Чанишвили Ш.Ш., Качарава Н.Ф., Бадридзе Г.Ш. Структурно-функциональные особенности листьев луговых растений в условиях высокогорий Малого Кавказа // Физиология и биохимия культурных растений. 2009. Т. 41. № 2. С. 132–139.
Шидаков Р.С. Сортимент яблони и совершенствование его путем селекции в предгорьях Северного Кавказа. Нальчик: Эльбрус, 1991. 303 с.
Шидаков Р.С., Шидакова А.С. Экономическая оценка сортов и элит яблони в агроландшафтах в разных экологических зонах Северного Кавказа // Успехи соврем. естествознания. 2006. № 11. С. 12–15.
Blanke M.M., Höfer M., Pring R.J. Stomata and structure of tetraploid apple leaves cultured in vitro // Ann. Bot. 1994. V. 73. P. 651–654.
Elias P. Stomata density and size of apple trees growing in irrigated and non irrigated conditions // Biol. Bratislava. 1995. V. 50. P. 115–118.
Franks P.J., Beerling D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time // Proc. Nat. Acad. Sci. 2009. V. 106. P. 10343–10347.
Franks P.J., Farquhar G.D. The mechanical diversity of stomata and its significance in gas-exchange control // Plant Physiol. 2007. V. 143. P.78–87.
Franks P.J., Franks P.J., Drake P.L., Beerling D.J. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: an analysis using Eucalyptus globulus // Plant, Cell Environ. 2009. V. 32. P. 1737–1748.
Frohnmeyer H., Staiger D. Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection // Plant Physiol. 2003. V. 133. P. 1420–1428.
Ganeva T., Uzunova K. Comparative leaf epidermis study in species of genus Malus Mill. (Rosaceae) // Bot. Serbica. 2010. V. 34. P. 45–49.
Giday H., Kjaeк K.H., Fanourakis D., Ottosen C.O. Smaller stomata require less severe leaf drying to close: a case study in Rosa hydrida // J. Plant Physiol. 2013. V. 170. P. 1309–1316.
Gou X., Gou X., Chen F., Yang M., Li J., Peng J., Jin L. Climatic response of thick leaf spruce (Picea crassifolia) tree-ringwidth at different elevations over Qilian Mountains, northwestern China // J. Arid Environ. 2005. V. 61. P. 513–524.
Hetherington A.M., Woodward F.I. The role of stomata in sensing and driving environmental change // Nature. 2003. V. 424. P. 901–908.
Kessler M., Siorak Y., Wunderlich M., Wegner C. Patterns of morphological leaf traits among pteridophytes along humidity and temperature gradients in the Bolivian Andes // Functional Plant Biol. 2007. V. 34. P. 963–971.
Kofidis G., Bosabalidis A.M., Moustakas M. Contemporary seasonal and altitudinal variations of leaf structural features in oregano (Origanum vulgare L.) // Ann. Bot. 2003. V. 92. P. 635–645.
Kouwenberg L., Kürschner W.M., McElwain J.C. Stomatal frequency change over altitudinal gradients: prospects for paleoaltimetry // Rev. Mineral. Geochem. 2007. V. 66. P. 215–241.
Körner C., Bannister P., Mark A.F. Altitudinal variation in stomatal conductance, nitrogen content and leaf anatomy in different plant life forms in New Zealand // Oecologia. 1986. V. 69. P. 577–588.
Ma Z.S., Jia Y.Y., Wang Y.H., Xuan L.F. Observation of leaf stomata shapes and distribution of large and small stomatas of peach (Prunus persica L.)[J] // J. Agricult. Univ. Hebei. 2008. V. 6. P. 018.
Mizutani M., Kanaoka M.M. Environmental sensing and morphological plasticity in plants // Seminars in cell & developmental biology. N.Y.: Acad. Press, 2017. V. 83. P. 69–77.
Nawkar G.M., Maibam P., Park J.H., Sahi V.P., Lee S.Y., Kang C.H. UV-induced cell death in plants // Intern. J. Mol. Sci. 2013. V. 14. P. 1608–1628.
Royer D.L. Stomatal density and stomatal index as indicators of paleoatmospheric CO2 concentration // Rev. Palaeobot. Palynol. 2001. V. 114. P. 1–28.
Šantrůček J., Vráblová M., Šimková M., Hronková M., Drtinová M., Květoň J., Neuwithová J. Stomatal and pavement cell density linked to leaf internal CO2 concentration // Ann. Bot. 2014 V. 114. P. 191–202.
Stenström A., Jónsdóttir I.S., Augner M. Genetic and environmental effects on morphology in clonal sedges in the Eurasian Arctic //Am. J. Bot. 2002. V. 89. P. 1410–1421.
Warabieda W., Olszak R.W., Dyki B. Morphological and anatomical characters of apple leaves associated with cultivar susceptibility to spider mite infestation // Acta Agrobot. 1997. V. 50. P. 53–64.
Wild A., Wolf G. The effect of different light intensities on the frequency and size of stomata, the size of cells, the number, size and chlorophyll content of chloroplasts in the mesophyll and the guard cells during the ontogeny of primary leaves of Sinapis alba // Z. Pflanzenphysiol. 1980. V. 97. P. 325–342.
Yang X., Yang Y., Ji C., Feng T., Shi Y., Lin L., He J.S. Large-scale patterns of stomatal traits in Tibetan and Mongolian grassland species // Basic Appl. Ecol. 2014. V. 15. P. 122–132.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая