

Известия РАН. Серия биологическая, 2021, № 2, стр. 184-192
Влияние фактора искусственных насаждений на восстановление вторичной целины в агролесоландшафтах сухих степей
А. М. Пугачёва *
Федеральный научный центр агроэкологии, комплексных мелиораций и защитного лесоразведения РАН
400062 Волгоград, Университетский пос. 97, Россия
* E-mail: nir-1@mail.ru
Поступила в редакцию 07.04.2018
После доработки 07.11.2018
Принята к публикации 29.11.2018
Аннотация
Впервые с использованием математических методов оценено восстановление вторичной целины на залежных землях сухих степей под влиянием разных типов искусственных защитных лесных насаждений – обязательного компонента современных агролесоландшафтов. Установлено достоверное влияние насаждений на видовой состав растительных сообществ, подтвержденное результатами дисперсионного анализа, ранговой корреляцией Спирмена и методом корреляционных плеяд. Отмечено, что большее воздействие куртин на состав растительных сообществ наблюдается вблизи насаждений. Обнаружено максимальное значение коэффициента сходства с целиной на удаленных от насаждений объектах. Показано, что при линейном размещении насаждений залежи их опушечных зон имеют сходство видового состава с фитоценозами прилегающих территорий.
Процесс движения земель сельскохозяйственного назначения и их перевод из одной категории использования в другую в течение длительного времени распространялись на все почвенно-климатические зоны России и различались продолжительностью, динамикой и масштабами. Последствия этих трансформаций – залежный клин земель сельскохозяйственного назначения площадью 35 млн га (Иванов, 2015). Перевод земель из категории сельскохозяйственного использования в категорию залежь характеризуется их самозарастанием и формированием вторичных фитоценозов, по видовому составу не всегда соответствующих зональным сообществам (Титлянова и др., 1993). Степная зона наиболее сильно затронута трансформационными процессами, связанными с аграрной эксплуатацией земель. Этим объясняется практически полное уничтожение степного биома (Национальная …, 2001). Залежные земли в Российской Федерации – основной территориальный резерв его восстановления (Смелянский, 2012). Из-за большой площади распашки восстановление осложняется территориальной отдаленностью редко расположенных очагов инспермации, являющихся основным источником пополнения банка семян степных видов растений, а также биологией самих видов. Особенности жизненного цикла аборигенных представителей флоры, зависимость репродуктивной активности, например, плотнодерновинных злаков от погодных факторов, в частности от соотношения тепла и влаги, непосредственно влияют на формирование генеративных побегов и сказываются на скорости и интенсивности их расселения (Левыкин, 2006). Отклонение от климатических оптимумов приводит к нарушению цикла их репродукции, удлиняя период восстановления терминальных фитоценозов. Закономерные смены растительности на залежах зависят как от внутренних процессов, происходящих в растительных сообществах, так и от внешних факторов, которые оказывают влияние на их продолжительность.
Изучением этих трансформаций занимаются многие исследователи. В основном они рассматривают внутренние процессы в сообществах (Александрова, 1964; Титлянова и др., 1993; Абаимов и др., 2012; Сергеева и др., 2015), а также изучают сукцессионные смены под влиянием внешних факторов (Гусев, 2008, 2009, 2012, 2015; Котлярова, 2008; Казанцева и др., 2010). Результаты исследований по Центрально-Черноземной зоне России, юго-востока Белоруссии доступны. Для сухих степей юга Приволжской возвышенности (зона исследований) изучение восстановления растительных сообществ в постагрогенных лесоландшафтах отличается новизной и характеризуется большой значимостью. Сухие степи занимают в России площадь 28 млн га (Орлов, Бирюкова, 1995). По мнению И.Н. Сафроновой, они подвергались сильному изменению и практически полностью распаханы (Сафронова, 2006).
В национальной стратегии сохранения биоразнообразия России признанным способом сбережения природных популяций значится их восстановление in situ. Один из путей реставрации биосистем – размещение на землях сельскохозяйственного назначения искусственных защитных лесных насаждений (ЗЛН), формирующих агролесомелиоративные ландшафты. ЗЛН – признанные стабилизаторы экологической составляющей экосистем, влияющие на процессы, происходящие на сопряженных территориях, в том числе на формирование видового состава сообществ постагрогенной растительности (Пугачёва, 2015; Кулик, Пугачёва, 2016). В настоящее время на территории РФ площадь под искусственными ЗЛН разного назначения составляет 204.5 млн га, 80% из которых – спелые насаждения, достигшие оптимальных параметров своего развития и оказывающие максимальное и разнообразное влияние на прилегающие территории (Кулик и др., 2018).
Цель работы – оценка восстановления вторичной целины на залежах агролесоландшафтов сухих степей юга Приволжской возвышенности с использованием математических методов, а также определение роли внешних факторов в восстановлении отдельных ценотических групп растений на одновозрастных залежах и влияние насаждений разных типов в сукцессионных рядах.
МАТЕРИАЛЫ И МЕТОДЫ
Создание в сухих степях юга Приволжской возвышенности эталонного агролесоландшафта (системы ЗЛН “Качалинское”, расположенной в Иловлинском р-не Волгоградской обл.) обусловлено активным развитием деградационных процессов территории, связанных с эрозионными явлениями, в первую очередь с дефляцией. Регион в силу своего географического расположения попадает под активное влияние воздушных масс различного географического типа (арктического, полярного и тропического воздуха), определяющих формирование пыльных бурь (Сажин, 2017). Общая площадь землепользования 7297 га; на ней в 1982 г. были заложены линейные полезащитные ЗЛН (ПЗЛН) насаждения, расположенные вдоль полей севооборотов, и куртинные (противоэрозионные), приуроченные к понижениям рельефа. По дистанционному мониторингу лаборатории аэрокосмических методов исследований ФНЦ агроэкологии РАН на данной территории были определены имеющиеся понижения мезо- и микрорельефа, которые отличаются лучшими почвенными характеристиками и условиями влагообеспеченности; с их учетом и были заложены куртинные насаждения (Рулев, Пугачёва, 2014). Наличие множества понижений на ограниченной территории характеризует ее специфику, что обусловливает дополнительные возможности облесения засушливых регионов. Часть пашни агролесоландшафта в 1992 г. была выведена из сельскохозяйственного использования, на ней более 25 лет происходят процессы самозарастания и самовосстановления. По данным 2010 г. площадь залежей составила 30% общей площади территории. Почвы района исследований тяжелосуглинистые, комплексные, с содержанием гумуса, не превышающим 2%, за исключением понижений с лугово-каштановыми почвами, где показатели почвенного плодородия достигают 4%. Климат характеризуется резкой континентальностью (сумма годового количества осадков до 400 мм) и гидротермическим коэффициентом в пределах 0.6. Контролем был участок целины, также расположенный на изучаемой территории.
ПЗЛН занимают площадь 122.7 га. Они созданы по универсальной схеме, характерной для зон недостаточного увлажнения. Схема смешения четырехрядная: два ряда – внутренние основные древесные породы и два ряда – внешние кустарники (сопутствующие породы). Расстояние между рядами 3, ширина закраек 3, общая ширина полос 15 м. Изучались залежи опушечных зон (закраек) вдоль лесных полос с участием следующих древесных видов растений: Quercus robur L. и Ribes aureum Pursh. (см-Д-Д-см) – объект L1; Pinus nigra subsp. pallasiana (Lamb.) и Lonicera tatarica L. (ж-Ск-Ск-ж) – объект L2; Ulmus pumilla L. и Ribes aureum Pursh. (см-Вз-Вз-см) – объект L3. Из-за специфики изучаемых залежных объектов закраек, представляющих собой полосу шириной 3 м вдоль насаждений, пробные площадки составляли 3 × 9 м2, что соответствует допустимому размеру для изучения растительности в сухих степях (Миркин и др., 2008). Так как сходство флор залежей изучалось сравнением их ботанических списков с видовым составом растительных сообществ прилегающих территорий, в исследованиях рассматриваются дополнительно три объекта: L4 – залежь начального этапа зарастания, бурьянистая стадия (граничит с L2); L5 – агроценоз (с L3); Z – целина (контроль) с L1.
Площадь объектов, находящихся под влиянием насаждений куртинного типа (противоэрозионных), 135 га. Их породный состав представлен разнообразным ассортиментом декоративных, плодовых и селекционно-улучшенных форм деревьев и кустарников. В состав входят: Q. robur L. var. pyramidalis, Robinia pseudoacacia L., P. nigra subsp. рallasiana (Lamb.) Holmboe, Fraxinus lanceolata Borkh., Ulmus laevis Pall., Pyrus communis L., Hippophae rhamnoides L., Prunus padus L., Aronia melanocarpa (Michx.) Elliott, R. aureum Pursh, Cotinus coggygria Scop. (Rhus cotinus L.), L. tatarica L. и др. Названия видов представлены по флористической сводке (Черепанов, 1995).
Для достижения поставленной цели математическому анализу подверглись сформированные ранее ботанические списки залежных объектов, размещенных на разных расстояниях от куртинных насаждений (30, 60, 90, 120 м для К1, К2, К3, К4 соответственно) – пробные площадки размером 10 × 10 м2, по 10 площадок на каждом варианте. Исследования растительных сообществ проводили с 2008 по 2016 г. по стандартной методике (Программа…, 1974; Методика…, 1985). При сопоставлении флоры изучаемых объектов особое внимание уделялось видовому составу и ценотическому спектру. Сукцессии залежной растительности рассматривались с позиции их детерминированности (за основу взяты характерные для залежей сухих степей сукцессионные ряды) (Титлянова, Самбуу, 2014; Кулик, Пугачёва, 2016).
Полученные и опубликованные ранее результаты по видовому составу растительных сообществ и их обилию позволили сделать вывод о стадиях развития залежей под влиянием насаждений разных типов (Кулик, Пугачёва, 2016). Для подтверждения достоверного влияния насаждений на развитие растительных сообществ на примере степных и сорных видов, доминирующие позиции которых в сообществах характеризуют разные циклы зарастания, применялся дисперсионный анализ. Методом корреляционных плеяд при выявлении сходства видов, определены объекты залежей, имеющие сходство с целиной и приближающиеся в своем развитии к терминальной стадии. Из множества существующих коэффициентов сходства флор выбран коэффициент Серенсена–Чекановского (KSC). По квадратной корреляционной матрице посредством построения графического изображения проведен анализ структуры статистически значимых связей. Основателем метода предложено несколько способов анализа; в данной работе выбран метод корреляционных граф (Терентьев, 1959). Индикатором являлся состав растительных сообществ целины, на основе сходства с ним построены дендриты максимального корреляционного пути и выделены корреляционные плеяды (Шмидт, 1984).
Для расчета показателей сходства систематической структуры флор между ранжированными рядами ведущих семейств был использован коэффициент ранговой корреляции Спирмена, выбранный в связи с наличием связанных рангов во всех ранжированных рядах.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Растительный покров климаксовых ценозов Волгоградской обл. хорошо изучен. В основном данные относятся к концу XIX–началу XX в. Геоботанические экспедиции здесь осуществляли И.И. Лепехин, И.Г. Гмелин, П.С. Паллас, исследования проводили А.Н. Краснов, И.К. Пачоский, Б.А. Келлер, Г.Н. Высоцкий, К.М. Залесский, И.В. Новопокровский, Е.М. Лавренко и другие ученые. Недавно проведенные локальные исследования касаются прежде всего особо охраняемых природных территорий (Бялт и др., 2012; Паршутина, 2015).
После антропогенного нарушения почвенного покрова изучаемого агролесоландшафта, связанного с закладкой самих насаждений и выращиванием на территории в течение 10 лет сельскохозяйственных культур Tríticum aestívum L., начался обратный процесс, связанный с выводом земель из оборота (процесс естественного самозарастания и самовосстановления). После 25-летнего восстановительного периода на объектах залежи сформировались растительные сообщества, которые различались видовым составом и характеризовались разными стадиями зарастания. По результатам проведенных ранее исследований залежных объектов в изучаемом агролесоландшафте были обнаружены как виды, свойственные фитоценозам сухих степей (Stipa lessingiana Trin & Rupr., Festuca valesiaca Gaudin), так и редкие виды (Tulipa gesneriana L. (T. schrenkii Regel)) – представители коренных ценозов изучаемой территории.
По анализу ботанических списков, включающих в себя 114 видов высших сосудистых растений, из 29 семейств и 72 родов основными были – Poaceae, Asteraceae, Fabaceae, Brassicaceae, Lamiaceae, но доли их участия различались. В табл. 1 представлено долевое участие семейств по числу видов. Несмотря на одинаковый период восстановления, отмечаются различия в составах растительных сообществ, что подтверждает разные стадии их развития.
Таблица 1.
Доля семейств в общем числе видов во флорах объектов исследований, %
| Семейство | Целина (контроль) |
Залежные объекты | ||||||
|---|---|---|---|---|---|---|---|---|
| под влиянием куртинных насаждений | под влиянием линейных насаждений | |||||||
| обозначения объектов исследований | ||||||||
| Z | K1 | K2 | K3 | K4 | L1 | L2 | L3 | |
| Poaceae | 24.0 | 3.4 | 8.9 | 25.3 | 25.6 | 26.2 | 16.7 | 19.4 |
| Asteraceae | 20.3 | 25.6 | 23.6 | 22.7 | 20.2 | 21.3 | 26.2 | 30.6 |
| Fabaceae | 8.1 | 13.9 | 12.4 | 9.3 | 8.1 | 8.2 | 2.4 | 8.3 |
| Brassicaceae | 9.5 | 10.4 | 11.2 | 6.7 | 9.4 | 6.6 | 16.7 | 13.9 |
| Lamiaceae | 6.7 | 6.9 | 5.6 | 2.7 | 5.4 | 8.2 | 7.1 | 5.5 |
| Caryophyllaceae | 2.7 | 3.4 | 2.3 | 1.3 | 1.4 | 3.4 | – | – |
| Rubiaceae | – | 1.2 | 1.1 | 1.3 | 1.4 | – | – | – |
| Apiaceae | 1.4 | 1.2 | 1.1 | 1.3 | 1.4 | – | – | – |
| Scrophulariaceae | 4.1 | 3.4 | 3.5 | 4.2 | 4.0 | 4.9 | 4.7 | 5.5 |
| Ranunculaceae | 1.4 | 2.3 | 2.3 | 2.7 | 2.7 | – | – | – |
| Umbelliferae | 1.4 | 1.2 | 1.1 | 1.3 | 1.4 | 1.6 | 2.4 | – |
| Cyperaceae | – | – | – | 1.3 | – | – | – | – |
| Euphorbiaceae | 1.4 | 2.3 | 2.3 | 1.3 | 1.4 | 1.6 | 2.4 | 2.8 |
| Chenopodiaceae | – | 4.7 | 4.5 | 1.3 | – | – | 4.7 | 2.8 |
| Convolvulaceae | 1.4 | 1.2 | 1.1 | – | 1.4 | 1.6 | 2.4 | 2.8 |
| Polygonaceae | – | 1.2 | 1.1 | – | – | – | – | – |
| Rosaceae | 1.4 | 1.2 | 1.1 | 1.3 | 1.4 | 3.4 | 2.4 | 2.8 |
| Aristolochiaceae | – | 1.2 | 1.1 | – | – | – | – | – |
| Plantaginaceae | 1.4 | 2.3 | 2.3 | 2.7 | 1.4 | 1.6 | 4.7 | 2.8 |
| Dipsacaceae | – | 1.2 | 1.1 | 1.3 | – | – | – | – |
| Boraginaceae | 2.7 | 5.8 | 5.6 | 6.7 | 4.0 | 3.4 | – | – |
| Amaranthaceae | – | 4.2 | 1.1 | – | – | 1.6 | 2.4 | – |
| Orobanchaceae | 1.4 | 1.2 | 1.1 | – | 1.4 | 1.6 | – | – |
| Liliaceae | 2.7 | 1.2 | 1.1 | 1.3 | 2.7 | – | – | – |
| Linaceae | 1.4 | 1.2 | 1.1 | 1.3 | – | 1.6 | – | – |
| Plumbaginaceae | 2.7 | 1.2 | 2.3 | 2.7 | 2.7 | – | – | – |
| Labiatae | 1.3 | – | – | – | 1.3 | 1.6 | 2.4 | 2.8 |
| Crassulaceae | 1.3 | – | – | – | – | 1.6 | 2.4 | – |
| Amaryllidaceae | 1.3 | – | – | – | 1.3 | – | – | – |
Примечание. “–” – отсутствие данного вида на пробных площадках; для табл. 1 и 2.
На залежных объектах наблюдаются начальные стадии зарастания: бурьянистая – К1, L2 (доля степных видов составляет здесь 44–51%); корневищная – К2 (доля степных видов достигает 55%), а также завершающие: рыхлодерновинная и плотнодерновинная – К4, L1 (доля степных видов в них возрастает до 72%).
В вариантах, где развитие сообществ соответствует завершающим стадиям развития, максимальная доля растений семействa Poaceae составляет 26.2%, в остальных вариантах основные позиции занимает семейство Asteraceae – К1, К2, L2, L3 (до 30.6%). К основным относятся также семейства Brassiсaceae (до 16.7%), Fabaceae (до 13.9%), Lamiaceae (до 8.2%). Доля оставшихся 24 семейств в сообществах составляет 1.1–4.1%.
На рис. 1 представлена диаграмма наличия в залежных фитоценозах под влиянием куртинных насаждений степной и сорной ценотических групп, характеризующих начало (сорная и степная) и завершение сукцессионного ряда. Оценка их присутствия в фитоценозах выявила сильную обратную корреляционную зависимость (r = − 0.91), характеризующуюся по мере удаленности объектов от насаждений К1 → К4 увеличением числа степных видов от 51 до 72% с коэффициентом аппроксимации R2 = 0.971 при уменьшении сорных от 20 до 8% (R2 = 0.7975) (рис. 1).
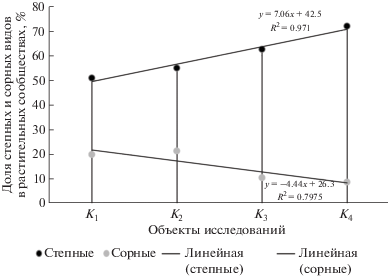
С применением однофакторного дисперсионного анализa для проверки гипотезы влияния куртинных насаждений на содержание видов в растительных сообществах доказана достоверность его влияния с 96%-ным уровнем, влияние неучтенных факторов – 4% (табл. 2). Полученные результаты подтверждены критерием Фишера, значения которого (F = 45.7 и 129.5 для степных и сорных видов) превышают критические.
Таблица 2.
Результаты дисперсионного анализа влияния фактора куртинных насаждений на содержание степных и сорных видов в залежных фитоценозах
| Разнообразие | Дисперсии (суммы квадратов) |
V | Q2 | Проверка достоверности фактора | ||||
|---|---|---|---|---|---|---|---|---|
| степные | сорные | степные | сорные | степные | сорные | степные | сорные | |
| Факториальное (межгрупповое) | 523 | 583 | 3 | 3 | 174 | 194.3 | $\eta _{х}^{2}$ = 0.96 ± 0.0064 Ф = $\frac{{0.96}}{{0.0064}}$ =150 F = $\frac{{174}}{{3.8}}$ = 45.7 |
$\eta _{х}^{2}$ = 0.96 ± 0.0075 Ф = $\frac{{0.96}}{{0.0075}}$ = 128 F = $\frac{{194.3}}{{1.5}}$ = 129.5 |
| Случайное (внутригрупповое) | 70 | 24 | 18 | 16 | 3.8 | 1.5 | ||
| Общее | 593 | 607 | 21 | 19 | – | – | Fst = (126.5 – 26.7 – 8.7) | Fst = (127.7 – 26.9 – 8.7) |
Коэффициенты ранговой корреляции (табл. 3) свидетельствуют о сходстве целины с объектом К4, достоверность подтверждается значением, превышающим уровень существенности 0.01. Для выявления связанных одним признаком групп, в нашем исследовании это видовой состав растительных сообществ, схожий с составом в целинном объекте, применен метод корреляционных плеяд. В табл. 4–6 в матрицах рассчитанных коэффициентов сходства флор постагрогенных объектов под влиянием насаждений разных типов по диагонали указано число выявленных в них видов. По матрице табл. 4 построен дендрит сходства флор изучаемых объектов (рис. 2).
Таблица 3.
Коэффициенты Спирмена ранговой корреляции Ps между вариантами под влиянием куртинных насаждений
| Объекты для сравнения | K1 | K2 | K3 | K4 |
|---|---|---|---|---|
| Z | 0.32 | 0.30 | 0.25 | 0.75** |
Примечание. Связь между параметрами значима * – при Ps > > 0.05; ** – при Ps > 0.01; для табл. 3 и 5.
Таблица 4.
Матрица сходства флор по коэффициенту Серенсена-Чекановского (KSC) объектов под влиянием куртинных защитных насаждений и целины
| Название объекта | Z | K1 | K2 | K3 | K4 |
|---|---|---|---|---|---|
| Z | 75 | 0.49 | 0.51 | 0.58 | 0.83 |
| K1 | 0.49 | 86 | 0.80 | 0.68 | 0.56 |
| K2 | 0.51 | 0.80 | 89 | 0.70 | 0.56 |
| K3 | 0.58 | 0.68 | 0.70 | 75 | 0.72 |
| K4 | 0.83 | 0.56 | 0.56 | 0.72 | 74 |
Рис. 2.
Дендрит и корреляционные плеяды флор залежных объектов и целины под влиянием куртинных защитных насаждений.
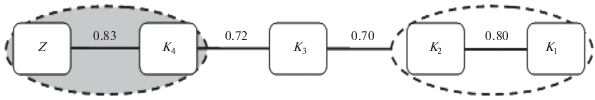
Обособленную плеяду наибольшего сходства с целиной (KSC ≥ 0.83) имеют удаленные пробные площадки залежи, находящиеся на расстоянии 120 м (К4) от насаждений. Данные варианты характеризуются рыхлодерновинной стадией зарастания с переходом в плотнодерновинную и приближаяются по видовому составу к терминальным сообществам. Также выделяется плеяда с показателем KSC ≥ 0.80, характеризующая сходство между собой залежных объектов К1 и К2, находящихся на расстояниях 30 и 60 м от куртин. Сходство близко расположенных от насаждений участков свидетельствует о воздействии насаждений на происходящие в растительных сообществах процессы. Объекты К1 и K2 находятся на начальных стадиях зарастания (несмотря на период восстановления >20 лет). Большая доля сходства видового состава в варианте K4 и целине свидетельствует о завершающей стадии развития растительного сообщества, что характерно для сухих степей после 25 лет восстановления без внешнего воздействия (в наших исследованиях – куртинных защитных насаждений).
Анализ систематической структуры флор залежных объектов под влиянием линейных насаждений выявил иные характеристики растительных сообществ на залежах (рис. 3). Сходство имеют как видовой состав сообществ на расположенных рядом территориях, так и их ценотические группы, что свидетельствует об особенностях развития залежных сообществ под влиянием линейных насаждений с индивидуальной характеристикой сукцессионных рядов.
Рис. 3.
Соотношение степных и сорных видов на расположенных рядом объектах под влиянием линейных насаждений, шт.
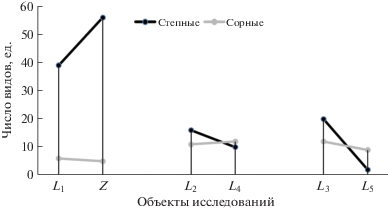
Рис. 4.
Дендрит и корреляционные плеяды сходства флор между изучаемыми объектами и целиной под влиянием линейных защитных насаждений.
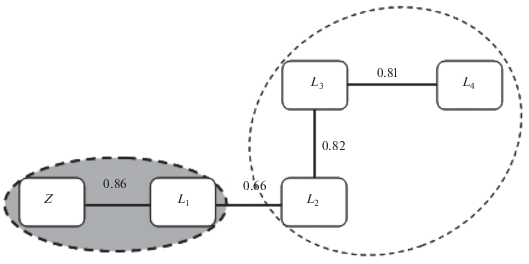
При сравнении систематической структуры флор изучаемых объектов под влиянием насаждений этого типа получены достоверные коэффициенты их сходства (табл. 5). Максимальное сходство с целиной имеет залежный объект прилегающей территории. Размещенные рядом объекты, находящиеся на начальных этапах сукцессионного развития также характеризуются максимальными коэффициентами соответствия.
Таблица 5.
Коэффициенты ранговой (семейства) корреляции Спирмена между вариантами исследований
| Объекты для сравнения | L1 | L2 | L3 |
|---|---|---|---|
| Z | 0.65** | 0.46* | 0.50** |
| L4 | 0.53** | 0.83** | 0.95** |
| L5 | 0.53** | 0.75** | 0.89** |
Таблица 6.
Матрица сходства флор по коэффициенту Серенсена-Чекановского залежных объектов под влиянием линейных защитных насаждений
| Название объекта | Z | L1 | L2 | L3 | L4 | L5 |
|---|---|---|---|---|---|---|
| Z | 74 | 0.86 | 0.47 | 0.53 | 0.27 | 0.16 |
| L1 | 0.86 | 61 | 0.55 | 0.66 | 0.42 | 0.18 |
| L2 | 0.47 | 0.55 | 36 | 0.82 | 0.81 | 0.47 |
| L3 | 0.53 | 0.66 | 0.82 | 42 | 0.68 | 0.42 |
| L4 | 0.27 | 0.42 | 0.81 | 0.68 | 28 | 0.55 |
| L5 | 0.16 | 0.18 | 0.47 | 0.42 | 0.55 | 15 |
Дендрит, построенный по матрице (табл. 6) с последующим выделением корреляционных плеяд наиболее сходных объектов по систематической структуре флор, свидетельствует о том, что, несмотря на одинаковый период восстановления залежей, сходством с целиной отличается только объект L1 (КSC ≥ 0.86), расположенный на прилегающей территории.
Также выделяется плеяда объектов L2 и L4 (КSC ≥ ≥ 0.81), характеризующая начальную бурьянистую стадию зарастания растительного сообщества, что свидетельствует о сходстве расположенных рядом объектов. Можно предположить, что, достигнув проектной высоты (у отдельных древесных видов она составляет 18 м), деревья создают естественные барьеры на пути распространения семян с отдаленных участков при свободном доступе семян с расположенных рядом территорий. Достоверность полученных результатов подтверждается и ранговой корреляцией Спирмена.
Применение метода корреляционных плеяд позволило наглядно оценить сходство изучаемых фитоценозов и сделать вывод о соответствии той или иной стадии развития растительных сообществ под влиянием насаждений разных типов сукцессионному ряду залежей сухих степей. Дисперсионный анализ подтвердил достоверное воздействие на эти процессы внешних факторов, которыми в наших исследованиях являются искусственные защитные насаждения.
Защитные лесные насаждения разных типов (неотъемлемая часть современных агролесоландшафтов, а также один из способов реставрации экосистем in situ) непосредственно влияют на скорость восстановления постагрогенных фитоценозов. Их воздействие зависит от типа насаждений.
Куртинные защитные насаждения, располагаясь на территории в понижениях микрорельефа, снижают влияние на видовые смены на залежах по мере удаленности объектов от насаждений. Варианты вблизи насаждений К1 и К2 (30–60 м от куртин) характеризуются начальными циклами зарастания (с коэффициентом соответствия целине КSC = 0.49–0.58). На расстоянии 120 м (К4) сукцессионные смены происходят закономерно для сухих степей, что характеризуется наличием 72% степных видов растений в фитоценозах КSC ≥ 0.83.
Залежи линейных объектов имеют максимальные коэффициенты сходства с фитоценозами прилегающих территорий, что предполагает наличие у линейных насаждений барьерного эффекта на пути привноса семян с отдаленных участков при беспрепятственном доступе с территорий, расположенных рядом. Это приводит к растягиванию на неопределенное время формирования вторичной целины.
Дисперсионный анализ подтвердил достоверность влияния фактора куртинных насаждений на содержаниe степных и сорных ценотических групп в сообществах залежной растительности на 96%. Критерий Фишера при этом превышает критические значения (F = 45–129).
Проведенный анализ ранговой корреляции Спирмена подтвердил полученные выводы примененных математических методов, указав на специфику влияния насаждений разных типов на восстановление видов в растительных сообществах залежей.
Список литературы
Абаимов В.Ф., Ледовский Н.В., Ходячих Н.В. Демутационные процессы растительности на залежах в сухостепной зоне Южного Урала // Изв. Оренбург. гос. аграрн. ун-та. 2012. № 3(35). С. 73–75.
Александрова В.Д. Изучение смен растительного покрова. М; Л.: Наука, 1964. Т. 3. С. 300–447.
Бялт В.В., Сагалаев В.А., Фирсов Г.А. Природная и адвентивная дендрофлора Нижнехоперского природного парка // Бюл. главного ботан. сада. 2012. № 3. С. 35–45.
Гусев А.П. Сукцессионная система как основа фитоиндикации динамики ландшафтов (на примере Полесской ландшафтной провинции) // Природ. ресурсы. 2008. № 2. С. 51–62.
Гусев А.П. Особенности начальной стадии восстановительной сукцессии в антропогенном ландшафте (на примере юго-востока Белоруссии) // Экология. 2009. № 3. С. 174–179.
Гусев А.П. Особенности сукцессий растительности в ландшафтах, нарушенных деятельностью человека (на примере юго-востока Белоруссии) // Сиб. экол. журн. 2012. № 2. С. 231–236.
Гусев А.П. Антропогенная трансформация ландшафтов и сукцессии растительности // Вестн. Тюмен. гос. ун-та. Экология и природопользование. 2015. Т. 1. № 2(2). С. 103–110.
Иванов А.Л. Рациональное использование и охрана земельных (почвенных) ресурсов Российской Федерации // Вестн. Рос. с.-х. науки. 2015. № 1. С.7–10.
Казанцева Т.И., Бобровская И.И., Тищенко В.В. Особенности восстановления залежной растительности луговых степей Центрального Черноземья (Воронежская область) // Арид. экосистемы. 2010. Т. 16. № 2(42). С. 76–86.
Котлярова Е.Г. Расширение биологического разнообразия в ландшафтных системах // Земледелие. 2008. № 2. С. 21–24.
Кулик К.Н., Пугачёва А.М. Структура растительных сообществ залежных земель в системе куртинных защитных лесных насаждений в сухих степях // Арид. экосистемы. 2016. Т. 22. № 1(66). С. 77–85.
Кулик К.Н., Иванов А.Л., Рулев А.С., Свинцов И.П., Павловский Е.С., Петров В.И., Барабанов А.Т., Манаенков А.С., Васильев Ю.И., Жданов Ю.М., Зыков И.Г., Кулик Н.Ф., Крючков С.Н., Маланина З.И., Семенютина А.В., Сухоруких Ю.И., Шульга В.Д., Юферев В.Г. Стратегия развития защитного лесоразведения в Российской Федерации на период до 2025 года. Волгоград: ВНИАЛМИ, 2018. 34 с.
Левыкин С.В. Особенности генеративной активности степных фитоценозов // Изв. Оренбург. гос. аграр. ун-та. 2006. Т. 3. № 11–1. С. 151–153.
Методика системных исследований лесоаграрных ландшафтов. М.: ВАСХНИЛ, 1985. 112 с.
Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности: Учебник. М.: Логос, 2001. 264 с.
Национальная стратегия сохранения биоразнообразия России. М.: РАН, Мин-во природ. ресурсов РФ, 2001. 76 с.
Орлов Д.С., Бирюкова О.Н. Запасы углерода органических соединений в почвах Российской Федерации // Почвоведение. 1995. № 1. С. 21–32.
Паршутина Л.П. Степи Западной окраины Нижнего Поволжья (Волгоградская область) // Ботан. журн. 2015. Т. 100. № 9. С. 886–908.
Программа и методика биоценотических исследований / Под ред. Дылиса Д.Ф. М.: Наука, 1974. 403 с.
Пугачёва А.М. Сукцессии залежной растительности под влиянием линейных защитных насаждений (на примере юга Приволжской возвышенности) // Проблемы регион. экологии. 2015. № 1. С. 44–47.
Рулев А.С., Пугачёва А.М. Почвенно-геоморфологические исследования ландшафтов Юга Прикаспийской возвышенности // Аграрн. науч. журн. 2014. № 12. С. 30–33.
Сажин А.Н., Кулик К.Н., Васильев Ю.И. Погода и климат Волгоградской области. Изд. 2-е, перераб. и доп. Волгоград: ФНЦ агроэкологии РАН, 2017. 334 с.
Сафронова И.Н. Можно ли подзону опустыненных степей называть экотоном? // Арид. экосистемы. 2006. Т. 12. № 30–31. С. 20–27.
Сергеева И.В., Шевченко Е.Н., Зябирова М.М. Особенности флоры и растительности разновозрастных залежных земель южной части Приволжской возвышенности Саратовской области // Аграрн. науч. журн. 2015. № 10. С. 26–28.
Смелянский И.Э. Сколько в степном регионе России залежей? // Степной бюл. 2012. № 36. С. 4–7.
Терентьев П.В. Метод корреляционных плеяд // Вестн. ЛГУ. 1959. № 9. С. 137–141.
Титлянова А.А., Самбуу А.Д. Детерминированность и синхронность залежной сукцессии в степях Тувы // Изв. РАН. Сер. биол. 2014. № 6. С. 621–630.
Титлянова А.А., Афанасьев Н.А., Наумова Н.Б., Андриевский B.C., Артамонова В.С., Булавко Г.И., Гантимурова Н.И., Косинова Л.Ю., Косых Н.П., Миронычева-Токарева Н.П., Мордкович Г.Д., Наумов А.В., Напрасникова Е.В., Половинко Г.П., Стебаева С.К., Якутин М.В. Сукцессии и биологический круговорот. Новосибирск: Наука Сибирская издательская фирма, 1993. 157 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
Шмидт В.М. Математические методы в ботанике: Учебное пособие. Л.: Изд-во ЛГУ, 1984. 288 с.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая