

Известия РАН. Серия биологическая, 2021, № 2, стр. 193-199
Фитоиндикационный индекс разнообразия условий среды
Л. А. Животовский 1, *, Г. О. Османова 2
1 Институт общей генетики РАН
119991 Москва, ул. Губкина, 3, Россия
2 Марийский государственный университет
424000 Йошкар-Ола, пл. Ленина, 1, Россия
* E-mail: levazh@gmail.com
Поступила в редакцию 28.04.2018
После доработки 03.08.2020
Принята к публикации 07.08.2020
Аннотация
Введен фитоиндикационный индекс разнообразия среды (ФИРС), основанный на нормированном перекрытии диапазонов экологических шкал всех видов растений, представленных в данном местообитании. ФИРС оценивает сколь разнообразны экологические ниши, освоенные растениями в данном местообитании в данный момент времени. Индекс может быть применен индивидуально к каждой шкале, а также представлять собой усредненные данные по набору шкал. Метод продемонстрирован с использованием экологических шкал Цыганова на примере различных местообитаний: от относительно однородных до крайне разнородных. Показано отсутствие причинной связи между значением ФИРС и числом видов в местообитании. Установлено, что наряду с другими показателями популяционного состава ФИРС может тестировать изменения условий местообитаний.
Растительный покров наиболее доступен для наблюдения и чутко реагирует на изменения окружающей среды. О его роли как индикаторе условий среды было отмечено еще в работах А. Гумбольдта, А. Декандоля, А. Гризебаха и других ботаников-географов, определивших тесную связь растительного покрова со средой (цит. по: Булохов, 2001). Это направление исследований было названо индикационной геоботаникой (Викторов и др., 1962), а затем фитоиндикацией (Корчагин, 1971).
Фитоиндикация (определение экологически важных параметров местообитания по видовому спектру произрастающих на нем растений) – основа геоботанических исследований в XX в. (Braun-Blanquet, 1964). Она позволяет оценить условия местообитания по почвенным, климатическим и другим экологическим шкалам. Каждая шкала для конкретного вида может быть представлена точечной оценкой – оптимумом вида по данной шкале (Landolt, 1977; Ellenberg et al., 1991) – либо как диапазон (интервал, амплитуда) условий по этой шкале, в пределах которых обитает данный вид (Раменский и др., 1956; Цыганов, 1983). Как показывает сравнительный анализ, разные шкалы имеют свои особенности, связанные с набором видов, их зональностью и обильностью (Зубкова, 2011). Рассмотрим амплитудные шкалы Цыганова (Цыганов, 1983), разработанные для лесных фитоценозов европейской части России, хотя подход к оценке разнообразия местообитаний применим также к шкалам Раменского для луговых фитоценозов и иным диапазонным шкалам.
Ранее (Цыганов, 1983, стр. 125–130) были предложены три метода обработки флористических списков по диапазонным шкалам. Первый – метод “экстремальных границ”, который позволяет оценить местообитания по трем параметрам: левой (L) и правой (R) границам перекрытия диапазонов всех видов и середине между ними. Условие применимости этого метода: ${\text{max(}}L{\text{)}} < {\text{min(}}R{\text{)}}$ (рис. 1). Если же общего перекрытия не существует, то выбирают наибольшее число видов, для которых общий интервал перекрытия существует, – это так называемый метод пересечения большинства интервалов. Третий метод – метод середины интервалов, основанный на усреднении середин диапазонов всех видов (с учетом или без учета степени участия вида). Этот метод применим даже в отсутствие перекрытия. Все эти методы и ассоциированные с ними статистики представлены в программном пакете EcoScaleWin (Зубкова и др., 2008).
Рис. 1.
Условный пример границ перекрытия диапазонов трех видов по шкале освещенности–затенения (Цыганов, 1983). Диапазоны условий для каждого вида представлены сплошной линией с левой (L) и правой (R) границами, интервал их перекрытия показан штриховой линией; для рис. 1 и 2.
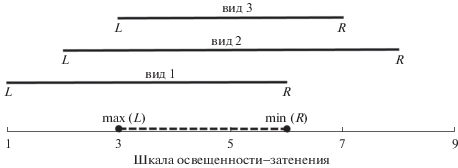
Все указанные методы дают средние оценки местообитания на соответствующей шкале. Вместе с тем важно оценить, насколько разнообразно освоено растениями данное местообитание по условиям среды, потому что даже на небольшой по размеру пробной площадке могут быть микропонижения, микроповышения, изменения кислотности или светового режима и т.п. Если все виды данного местообитания имеют значительный общий интервал перекрытия по разным шкалам, то это можно интерпретировать как освоение растениями небольшого разнообразия ниш данного местообитания. Напротив, если перекрытие небольшое или перекрытия нет, то это свидетельствует о том, что освоены более разнообразные ниши данного местообитания.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследования послужили флористические списки геоботанических описаний 11 местообитаний редкого в Республике Марий Эл растения – башмачка настоящего, или желтого Cypripedium calceolus L. (Красная …, 2013). Исследования C. calceolus были проведены в Волжском, Куженерском, Моркинском и Новоторъяльском районах Республики Марий Эл в разных фитоценозах (Животовский, Османова, 2018, 2020). Мы также проанализировали списки видов по шести экогеографическим агрегациям (Османова и др., 2019) и данные по всем видам (Цыганов, 1983). Материал обрабатывали с помощью четырех почвенных шкал Цыганова (увлажнения (Hd), солевого режима (Tr), богатства азотом (Nt), кислотности (Rc)), а также шкалы освещенности–затенения (Lc).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Фитоиндикационный индекс разнообразия среды. Для того чтобы определить индекс разнообразия среды, мы допускаем отрицательное значение перекрытия диапазонов, т.е. когда ${\text{max(}}L{\text{)}} < {\text{min(}}R{\text{)}}$ (рис. 2). Отметим, что абсолютное значение перекрытия не превышает размера шкалы. Поэтому мы вводим фитоиндикационный индекс разнообразия среды (ФИРС, Φ) данного местообитания по данной шкале s на основе нормированного значения перекрытия диапазонов всех видов, представленных в данном местообитании:
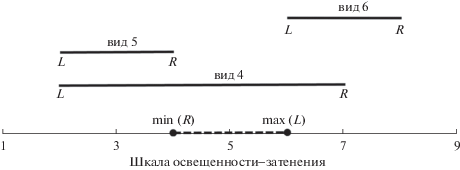
Чем менее разнообразны условия среды по отношению к данной экологической шкале, тем больше перекрытие диапазонов видов в данном местообитании и тем меньше значение ФИРС по этой шкале. Напротив, чем разнообразнее условия среды по этой шкале, тем меньше перекрытие диапазонов обитающих видов и тем больше значение ФИРС. Теоретически наименьшее значение ФИРС – 0, которое могло бы быть достигнуто, если бы левые края диапазона всех видов местообитания были бы равны 1, а их правые края – размеру шкалы $\left( {{{D}_{s}}} \right)$. Фактически же это малореально, хотя таких видов немало для разных шкал. Например, среди использованных нами шкал Цыганова имеются 50 таких видов из 2325 по шкале Lc, 9 видов из 1536 по шкале Nt, 104 вида из 1640 по шкале Rc; по шкалам Hd и Tr таких видов нет, однако есть виды, диапазоны которых занимают бóльшую часть этих шкал. Теоретически максимальное значение ФИРС – 1, если в данном фитоценозе имеются по крайней мере два вида с длиной диапазона 1, причем один из них представяет крайний левый край шкалы, а другой – правый. (Для одновидового сообщества максимальное значение ФИРС – 0.5). Однако по почвенным шкалам Цыганова оно недостижимо, так как длины диапазонов всех видов не меньше 3, за исключением армерии альпийской Armeria alpine Willd с длиной диапазона 2 по шкале Tr (Цыганов, 1983).
Наряду с частными значениями ФИРС по отдельным шкалам введем общее значение индекса как среднее по $S$ использованным шкалам:
Доверительный интервал оценки ФИРС. Из представленных выше формул можно получить точечную оценку ФИРС. При выборе метода определения доверительного интервала значений ФИРС важно учесть вероятностную природу его возможных случайных колебаний для данного местообитания. Действительно, в пределы выбранной пробной площадки могут случайно не попасть какие-то виды данного биоценоза ввиду малого размера пробного участка либо из-за невозможности охватить всю средовую гетерогенность из-за редкости видов. Поэтому подходящие статистические процедуры должны следовать такой структуре случайностей. Одна из них – это так называемая процедура складного ножа (СН) (Sokal, Rolf, 1995). Согласно этой процедуре (применительно к индексу ФИРС) из списка k видов убирается поочередно по одному виду и вычисляется оценка индекса по оставшимся $k - 1$ видам. Опишем ее детально.
Пусть Φ – значение какого-либо индекса, определяемого по списку k видов. Это может быть значение ФИРС по отдельной шкале s $({{\Phi }_{s}})$, либо среднее значение по нескольким шкалам $\left( {\bar {\Phi }} \right)$, либо любая другая их функция. Пусть ${{\Phi }_{{(i)}}}$ – моделируемое значение этого индекса, которое определяется по тому же списку, но с вычеркнутым из него видом под номером i. Таких моделируемых значений будет k (по числу видов в списке). Определим по набору этих модельных значений их СН-среднее $\left( {\tilde {\Phi }} \right)$ и стандартное отклонение (СН-ошибку, ${{s}_{\Phi }}$):
Значение $\tilde {\Phi }$, определенное методом складного ножа, может быть близким к исходной оценке $\Phi $, полученной по всем k видам, или значительно отклоняться от нее. В последнем случае исходная оценка $\Phi $ является смещенной, вместо нее берется значение $\tilde {\Phi }$. Стандартное отклонение (${{s}_{\Phi }}$) трактуется как его статистическая ошибка.
Особое внимание следует обратить на случаи повторов одних и тех же видов в флористическом списке данного местообитания. Это относится в основном к древесным растениям, когда вид представлен в двух или более ярусах. При анализе проективного покрытия ярусность важно учитывать, однако для анализа диапазонных характеристик этого не требуется, так как оцениваются условия среды, позволяющие данному виду существовать в любом статусе. Введение повторов видов не повлияет на точечную оценку значений ФИРС, так как их диапазоны идентичны по всем шкалам, но может повлиять на оценку доверительного интервала, так как СН-процедура основана на сокращении списка на один вид. При наличии повторов эта процедура убирает вид из одного яруса, но оставляет его в другом ярусе, и поэтому повторы в этой процедуре фактически не участвуют. Поэтому при оценке доверительного интервала методом СН предлагаем представлять вид однократно вне зависимости от его встречаемости в разных ярусах. По той же причине нельзя использовать бутстреп-процедуру, так как она приводит к появлению повторов видов из-за того, что любой вид может попасть несколько раз в моделируемую бутстреп-выборку.
Отметим возможные проблемы в СН-оценках. Если по данной шкале имеются два вида или более с одинаковым максимальным значением левых границ диапазонов и два вида или более с одинаковым минимальным значением их правых границ, то процедура СН окажется неэффективной, так как все ${{\Phi }_{{(i)}}}$ будут равны оценке Φ, из-за чего Φ будет совпадать с $\tilde {\Phi }$, а стандартная ошибка будет равна 0. При оценке по нескольким шкалам возможность такого совпадения невелика, но по отдельным шкалам возможна. В таком случае можно применить более сложную СН-процедуру с вычеркиванием двух или более видов (здесь не описана из-за ее сложности).
ФИРС для разных типов местообитаний с разным уровнем гетерогенности. Мы рассчитали и проанализировали оценки ФИРС $\left( {\bar {\Phi }} \right)$ по указанным выше пяти шкалам Цыганова для разных местообитаний (табл. 1). Согласно полученным значениям ФИРС, местообитания сгруппировались и расположились ступенчато (рис. 3). Для местообитания “болото”, где разнообразие условий среды наиболее однородное среди изученных нами местообитаний, значение ФИРС оказалось минимальным (0.381) (табл. 1). На рис. 3 это местообитание занимает первую ступень. Действительно, анализ флористического списка видов этого местообитания (багульник болотный Ledum palustre L., вахта трехлистная Menyanthes trifoliate L., вереск обыкновенный Calluna vulgaris (L.) Hull, водяника черная Empetrum nigrum L., голубика Vaccinium uliginosum L., клюква болотная Oxycoccus palustris Pers., морошка арктическая Rubus chamaemorus L., подбел многолистный Andromeda polifolia L., пушица влагалищная Eriophorum vaginatum L., росянка круглолистная Drosera rotundifolia L., сабельник болотный Comarum palustre L., хамедафна болотная Chamaedaphne calyculata (L.) Moench) показал, что по всем четырем почвенным шкалам Цыганова (увлажнения, солевого режима, богатства азотом и кислотности), а также по шкале освещенности–затенения диапазоны всех видов перекрываются, а в ряде случаев полностью совпадают.
Таблица 1.
Характеристики местообитаний и значения фитоиндикационного индексa разнообразия среды (ФИРС)
| Местообитания | Ассоциация | Число видов |
ФИРС | СН-оценка ФИРС |
|---|---|---|---|---|
| “Болото” | Кустарничково-сфагновая | 12 | 0.381 | 0.377 ± 0.0238 |
| Местообитания C. calceolus | ||||
| 1 | Молодой березняк елово-сосновый мшистый | 29 | 0.526 | 0.524 ± 0.0246 |
| 2 | Ельник липово-осоковый | 37 | 0.510 | 0.509 ± 0.0267 |
| 3 | Сосново-елово-лещиновый ландышево-коротконожко-орляковый лес | 9 | 0.429 | 0.424 ± 0.0139 |
| 4 (2009 г.) | Березово-елово-сосновый лес | 8 | 0.452 | 0.447 ± 0.0200 |
| 4 (2018 г.) | То же | 27 | 0.508 | 0.506 ± 0.0279 |
| 5 | Березняк с елью и сосной шершаво кульбабо-кошачьелапково-мшистый | 14 | 0.502 | 0.498 ± 0.0279 |
| 6 | Сосняк березово-ландышево-костяничный | 25 | 0.470 | 0.469 ± 0.0321 |
| 7 | Ельник ландышево-осоко-костяничный | 32 | 0.523 | 0.520 ± 0.0271 |
| 8 | Ельник березовый разнотравно-костяничный | 32 | 0.484 | 0.484 ± 0.0075 |
| 9 | Сосняк березово-еловый ландышево-костянично-папоротниковый | 39 | 0.488 | 0.487 ± 0.0212 |
| 10 | Сосняк елово-зеленомошный | 20 | 0.454 | 0.453 ± 0.0129 |
| 11 | Сосняк осоково-кисличный | 32 | 0.487 | 0.487 ± 0.0044 |
| Экогеографические агрегации | ||||
| ЭГА-1 | Экогеографические единицы, территориально близкие друг к другу и обитающие в сходных почвенных и иных условиях, объединены в экогеографические агрегации | 107 | 0.573 | 0.573 ± 0.0094 |
| ЭГА-2 | 114 | 0.573 | 0.573 ± 0.0100 | |
| ЭГА-4 | 45 | 0.582 | 0.581 ± 0.0240 | |
| ЭГА-5 | 110 | 0.558 | 0.558 ± 0.0069 | |
| ЭГА-10 | 104 | 0.570 | 0.569 ± 0.0225 | |
| ЭГА-11 | 118 | 0.565 | 0.564 ± 0.0108 | |
| Тотальные данные (Цыганов, 1983) | Сосудистые растения подзоны хвойно-широколиственных лесов европейской части России | 2362 | 0.775 | 0.775 |
Рис. 3.
Значения фитоиндикационного индекса разнообразия среды (ФИРС) $(\bar {\Phi })$ по четырем почвенным шкалам и шкале освещенности–затенения для различных типов местообитаний. Пунктирной линией отмечены средние значения ФИРС по группам местообитаний.
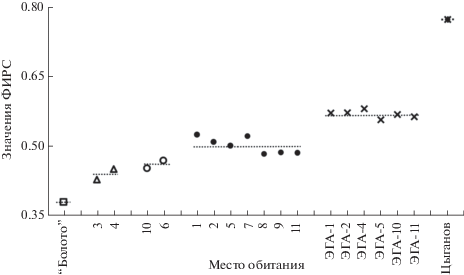
Так как местообитание “болото” относительно однородно по условиям среды, то для сравнения был проанализирован максимально разнородный материал по почвенным шкалам и шкале освещенности–затенения: 2362 вида сосудистых растений подзоны хвойно-широколиственных лесов европейской части России (Цыганов, 1983). Сравниваемые диапазоны шкал в большинстве случаев не перекрываются. Следовательно, это местообитание (большая по площади территория) неоднородно. Подтверждением этого стало высокое значение ФИРС – 0.775 (табл. 1). На рис. 3 это самая верхняя ступень. Между этими местообитаниями, крайними по гетерогенности условий среды и значениям ФИРС, расположились другие исследованные нами местообитания.
Анализ флористических списков 11 местообитаний редкого вида C. calceolus по шкалам Цыганова и значения ФИРС $(\bar {\Phi })$ позволили распределить эти местообитания на три группы (рис. 3). В первую группу попали местообитания 3 и 4 C. calceolus со значениями ФИРС 0.429 и 0.452 соответственно, они наиболее однородны. Причиной их меньшей гетерогенности стало влияние антропогенного фактора. В местообитании 3 (Куженерский р-н, ур. Нолькин Камень, склон южной экспозиции к левому притоку р. Нолька) была проведена выборочная рубка древостоя. Растительный покров был нарушен, а флористический состав был представлен всего девятью видами (табл. 1). Местообитание 4 (Моркинский р-н, близ д. Коркатово, склон северо-западной экспозиции горы Коркан-Курык, отвалы каменного карьера из мелкого щебня, мергеля, извести) было обследовано в 2009 г. В 2008 г. это местообитание C. calceolus в результате расширения дороги и очистки площади для собственных нужд работниками Коркатовского каменного карьера было нарушено, поэтому видовой состав стал очень бедным (табл. 1). Семь местообитаний C. calceolus (1, 2, 5, 7, 8, 9, 11) представляют собой ненарушенные фитоценозы с бóльшим разнообразием условий среды, значение ФИРС для них варьирует от 0.484 до 0.526 (табл. 1). Местообитания 6 и 10 оказались промежуточными между предыдущими группами по значениям ФИРС (0.470 и 0.454 соответственно) из-за особенностей почвенных условий (они расположены в зоне Марийско-Вятского увала).
Были проанализированы также списки видов растений, входящих в экогеографические агрегации (ЭГА), каждая из которых состояла из нескольких экогеографических единиц (ЭГЕ), территориально близких друг к другу и обитающих в сходных почвенных и иных условиях (Османова и др., 2019). Несмотря на то что по макропараметрам условий среды ЭГА внутренне однородны (Османова и др., 2019), значения ФИРС для них оказались более высокими (табл. 1, рис. 3), что указывает на наличие микрогетерогенности занимаемой ими территории, поскольку ЭГА занимают бóльшие площади, чем площади ранее рассмотренных местообитаний.
Таким образом, приведенные выше примеры наглядно показывают, что значение ФИРС оценивает гетерогенность местообитания по совокупности видов растений с использованием экологических шкал.
ФИРС и число видов. Можно было бы предположить, что значение ФИРС определяется числом видов (рис. 3). На самом же деле причинной положительной связи между ними нет. Действительно, различия между группами на рис. 3 по значениям ФИРС вызваны либо нарушением местообитаний, либо увеличением площади местообитания, что ведет к увеличению как гетерогенности условий, так и числа видов. Причинно же ФИРС и число видов не взаимосвязаны: ФИРС представляет собой совсем иной, независимый от числа видов, параметр местообитания. Из табл. 1, например, следует, что число видов в местообитании “болото” больше, чем в местообитаниях 3 и 4, однако значения ФИРС там меньше. Далее, в ЭГА-4 число видов намного меньше, чем в других ЭГА, однако значение ФИРС для него наибольшее среди ЭГА (табл. 1). Наконец, внутригрупповые положительные корреляции между значениями ФИРС и числом видов практически отсутствуют (рис. 4).
Рис. 4.
Отсутствие положительной взаимосвязи значений фитоиндикационного индекса разнообразия среды и числа видов. Обозначения местообитаний те же, что на рис. 3: черные кружочки – ненарушенные ассоциации с C. calceolus, крестики – ЭГА (знаком “плюс” отмечена ЭГА-4, включающая в себя всего 45 видов); пунктиром отмечены линии регрессии; R2 – коэффициент детерминации (для ЭГА вычислен без учета ЭГА-4).
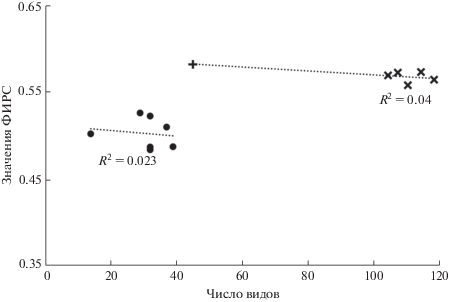
Следующий пример показывает, что местообитания с одинаковым числом видов могут значительно различаться по гетерогенности условий среды и, соответственно, по значениям ФИРС. Нами были заложены небольшие пробные площадки в двух разных фитоценозах (в ельнике и в экотонном сообществе), содержащие по 14 видов. Значения ФИРС для них оказались равными 0.410 и 0.523 соответственно (СН-оценки 0.410 ± 0.0042, 0.515 ± 0.0495). Этот пример свидетельствует о том, что значение ФИРС может увеличиться при внедрении чужеродных видов, если местообитание имеет соответствующие ниши с подходящими условиями.
Следует также иметь в виду, что действие общего экологического фактора может привести к одновременному изменению обоих параметров – числа видов и значения ФИРС. Например, в 2018 г. в местообитании 4 мы повторно провели геоботаническое описание. Число видов за десятилетие, прошедшее после антропогенного воздействия в 2008 г., увеличилось с 8 до 27, возросло и значение ФИРС – 0.508 (табл. 1), т.е. возобновляемый растительный покров стал “заполнять” микрониши этого местообитания видами с разными экологическими диапазонами, в результате чего увеличилось и число видов, и значение ФИРС.
ЗАКЛЮЧЕНИЕ
Индекс ФИРС оценивает в терминах диапазонных шкал, сколь разнообразны экологические ниши, освоенные растениями в данном местообитании в данный момент времени. Если значения индекса невелики, это еще не значит, что данное местообитание однородно по условиям среды, однако это значит, что по какой-то причине там обитают растения со сходными диапазонами экологических шкал, т.е. ими освоены ниши со сходными условиями среды. Если же значения индекса значительны, то это можно интерпретировать как большее число освоенных ниш к данному моменту времени.
ФИРС оценивается соответственно по максимальным и минимальным значениям левых и правых концов диапазонов видов данного местообитания. Можно ввести и другие индексы разнообразия по диапазонным шкалам, например, используя не крайние элементы порядковых рядов, как для ФИРС, а номера Т:
Для исследованного выше индекса ФИРС Т = 1. Взяв Т = 2, получим подобный ФИРС индекс, основанный на вторых по рангу величинах L и R. Можно взять Т = 3 (третьи по порядку L и R) и т.д. Предварительный анализ показал, что значения ФИРС при Т = 1 более контрастные, чем при Т > 1.
Таким образом, индекс ФИРС позволяет оценить разнообразие ниш, освоенных растениями в данном местообитании.
Работа выполнена в рамках гостемы ГЗ 0112-2019-0002 (подтема “Эколого-генетическая структура вида”).
Список литературы
Булохов А.Д. Травяная растительность Юго-Западного Нечерноземья России. Брянск: БГУ, 2001. 296 с.
Викторов С.В., Востокова Е.А., Вышивкин Д.Д. Введение в индикационную геоботанику. М.: Изд-во МГУ, 1962. 227 с.
Животовский Л.А., Османова Г.О. Эколого-географический подход к выявлению популяционной структуры вида у растений // Экология и география растений и растительных сообществ: Труды конф. Екатеринбург: Гуманитарный ун-т, 2018. С. 282–285.
Животовский Л.А., Османова Г.О. Экогеографические единицы и охрана внутривидового разнообразия // Изв. РАН. Сер. биол. 2020. № 2. С. 124–136.
Зубкова Е.В. О некоторых особенностях диапазонных экологических шкал растений Д.Н. Цыганова // Изв. СамарНЦ РАН. 2011. Т. 13. № 5. С. 48–53.
Зубкова Е.В., Ханина Л.Г., Грохлина Т.И., Дорогова Ю.А. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы “EcoScaleWin”. Йошкар-Ола: Изд-во Мар. гос. ун-та, 2008. 96 с.
Корчагин А.А. Теоретические вопросы фитоиндикации. Л.: Наука, 1971. 216 с.
Красная книга Республики Марий Эл. Т. Растения. Грибы / Сост. Богданов Г.А., Абрамов Н.В., Убранавичюс Г.П., Богданова Л.Г. Йошкар-Ола: Мар. гос. ун-т, 2013. 324 с.
Османова Г.О., Богданов Г.А., Животовский Л.А. Выделение многовидовых экогеографических агрегаций редких видов растений в целях организации охраняемых природных территорий (на примере флоры Республики Марий Эл) // Экология. 2019. № 5. С. 373–377.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипов Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 472 с.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 197 с.
Braun-Blanquet J. Pflanzensociologie. 3 Aufl. Wien; New York: Springer-Verlag, l964. 865 s.
Ellenberg H., Weber H.E., Dull R., Wirth V., Werner W., Paulisen D. Zeigerwerte von Pflanzen in Mitteleuropa [Indicator values of plants in Central Europe] // Scripta Geobotanics. V. 18. Gottingen: Verlag Erich Goltze KG, 1991. 248 s.
Landolt E. Okologische Zeigerwerts zur Sweizer Flora / Veroff. Geobot. Inst. ETH. Zurich. 1977. H. 64. S. 1–208.
Sokal R.R., Rolf F.J. Biometry. 3rd ed. N.Y.: W.H. Freeman & Co., 1995. 887 p.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая