

Известия РАН. Серия биологическая, 2021, № 5, стр. 521-527
Изменение соотношения типов реореакции у рыб при голодании
Д. С. Павлов 1, В. В. Костин 1, *, В. Ю. Паршина 1, Е. Д. Павлов 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский проспект, 33, Россия
* E-mail: povedenie@yandex.ru
Поступила в редакцию 18.11.2020
После доработки 09.12.2020
Принята к публикации 09.12.2020
Аннотация
Проведен анализ изменения мотивационной компоненты реореакции (соотношения типов реореакции) при голодании у ряда видов рыб. Показано, что характер изменения мотивационной компоненты реореакции у рыб в ответ на голодание во многом определяется наличием у них перед голоданием мотивации к миграции. У всех исследованных рыб семейств Cyprinidae и Salmonidae с изначально резидентным поведением наблюдается однотипный ответ на голодание: на 2-е–5-е сутки у них увеличивается предпочтение к движению по течению, а на 7-е–12-е сутки увеличивается предпочтение к движению против течения, и соотношение типов реореакции приближается к исходному. У молоди черноморской кумжи (Salmo trutta labrax) с мотивацией к покатной миграции наблюдается иной ответ на голодание – начиная с двух суток монотонно увеличивается движение особей по течению.
Реореакция (отношение рыб к течению) – это врождённая реакция, связанная с жизнью рыб в подвижной среде. Она проявляется в выборе рыбами участка водоема с оптимальными (для актуального поведения) скоростями течения и вектора собственного движения в нем. Реореакция является одним из основных механизмов поведения рыб в текучих водоемах, в том числе, их распределения, миграций и кочевок (Павлов 1979; Павлов и др., 2007).
Проявление реореакции определяют три компоненты: ориентационная (восприятие стимула), мотивационная (выбор типа реореакции), локомоторная (реализация реореакции выбранного типа) (Павлов, 1979; Pavlov et al., 2010; Павлов и др., 2020б). К показателям мотивационной компоненты относятся: реопреферендум рыб (выбор канала с течением или без него), избираемые скорости течения в реоградиенте (Павлов, 1979), соотношение типов реореакции (Maclean, Gee, 1971; Pavlov et al., 2010; Павлов и др., 2020 б). Рассмотрены три типа реореакции: положительный (ПТР) – движение против течения; отрицательный (ОТР) – движение по течению; компенсаторный (КТР) – сохранение своего положения относительно неподвижных ориентиров. Эти типы отношения рыб к течению отмечали и другие авторы (Johnston et al., 2017).
Изменения реореакции под воздействием различных факторов среды имеют разные причины. Известно, что мотивационная компонента реореакции в период миграции молоди с нерестилищ к местам нагула быстро (минуты, часы) и кардинально изменяется в зависимости от освещенности (Павлов и др., 2007; Павлов и др., 2019) и химического состава воды (Звездин, 2016). В этих случаях особи постоянно находятся в миграционном состоянии, а изменение реореакции определяется сменой их актуального поведения, например, контранатантной миграции на денатантную у молоди нерки Oncorhynchus nerka при отсутствии “запаха озера” или смене в сумерки резидентного поведения на миграционное у молоди многих видов при покатной миграции в реке. Кроме того, реореакция молоди изменяется при длительном воздействии неблагоприятных факторов различной природы. В этом случае у рыб, которые изначально были без мотивации к миграции, изменение реореакции определяется формированием миграционного состояния (Павлов и др., 2010; Пономарева, 2014).
В естественных условиях трофический фактор является наиболее частой причиной кочевок и миграций рыб (Olsson et al., 2006; Павлов и др., 2007; Flecker et al., 2010; Ferguson et al., 2019). При голодании рыб происходит быстрое (дни, недели) формирование миграционного поведения, что изменяет их реореакцию (Павлов и др., 2010). При этом у рыб модифицируются все три компоненты реореакции. В ряде специальных исследований были изучены изменения ориентационной (Gaffron, 1934; Harden Jones, 1963; Павлов, 1979) и локомоторной (Этинген, 1964; MacLeod, 1967; Laurence, 1972; Сабуренков, 1973; Павлов, 1979 и др.) компонент реореакции рыб под влиянием голода. Специальных исследований изменения количественных параметров мотивационной компоненты реореакции рыб в литературе нами не найдено. Некоторые сведения о ее изменении получены при исследовании физиологических и поведенческих механизмов миграционного поведения рыб (Павлов, 1979; Павлов и др., 2007; 2010; 2016 и др.).
Цель данной работы – экспериментальная оценка динамики соотношения типов реореакции при голодании рыб.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проводили в лаборатории ИПЭЭ РАН в 2018 г. Использовали особей 4-х видов рыб: верховка (Leucaspius delineatus), средняя длина тела 31.7 ± 0.25 (28–36) мм; данио рерио (Danio rerio) 26.5 ± 0.27 (24–33) мм; серебряный карась (Carassius auratus) 94.6 ± 1.14 (78–116) мм; микижа (радужная форель, Parasalmo mykiss (=Oncorhynchus mykiss)) 105.6 ± 0.97 (88–120) мм. Латинские названия видов приводятся по (Аннотированный каталог…, 1998). Верховку отловили мальковой волокушей в речках, а карася – в прудах Ярославской обл. Радужная форель была взята на подмосковном рыборазводном заводе. Данио рерио (беспородные) взяты из аквариальной ИПЭЭ РАН.
До начала экспериментов рыб кормили один раз в сутки, давая избыточное количество корма. Данио рерио и верховке давали корм для аквариумных рыб Prodac Tropical Fish Flakes, а радужной форели и серебряному карасю – искусственный корм BioMar. После начала экспериментов кормление рыб полностью прекращали, последний раз рыб кормили в 0-е сутки за час до начала тестирования. Рыб содержали в аэрируемых аквариумах при плотности посадки: 1 экз/7 л воды (данио рерио и верховка) и 1 экз/10 л воды (карась и радужная форель). Температуру поддерживали с помощью терморегулятора на уровне 18°C (карась, радужная форель, верховка) и 22°C (данио рерио). Освещeнность в лаборатории с 10 до 19 ч составляла 450–500 лк; в остальное время суток искусственное освещение выключали.
Для определения соотношения типов реореакции использовали прямоточные гидродинамические установки типа “Рыбоход”, позволяющие определять три типа реореакции – ПТР, КТР и ОТР (Pavlov et al., 2010). Такая установка представляла собой лоток, разделенный поперечными перегородками на 9 отсеков, средний отсек (№ 5) был стартовым. В зависимости от размера рыб использовали два варианта установки: для серебряного карася и радужной форели длина “Рыбохода” составляла 200 см, ширина – 60 см; для данио рерио и верховки – 110 и 20 см соответственно.
Опыты проводили на группах из 6 рыб. В начале эксперимента рыб помещали в стартовый отсек (№ 5), предварительно изолированный съемными сетками. После 20 мин акклимации снимали сетки стартового отсека, и начинался опыт. Через 30 мин подсчитывали число рыб в отсеках установки.
Для сравнимости с предыдущими публикациями опыты проводили на 0-е, 2-е, 5-е, 7-е, 10-е и 12-е сутки после последнего кормления.
На рыбах каждого вида было проведено по 60 опытов и использовано по 60 экз.
По экспериментальным данным рассчитывали частоты проявления каждого типа реореакции:
где: N – число исследованных рыб, экз., ni – число рыб в 1–4 отсеках рыбохода, экз., n5 – число рыб в 5 отсеке (стартовом), экз., nj – число рыб в 6–9 отсеках, экз., i, j – номер отсека в диапазоне 1–4 и 6–9 соответственно.
Кроме соотношения типов реореакции рассчитывали и индекс контранатантности (Ik), который отражает соотношение динамических типов реореакции (ПТР и ОТР). Индекс контранатантности оценивает перемещение рыб относительно стартового отсека, приведенное к шкале –1…+1. Расчет этого показателя проводили по формуле:
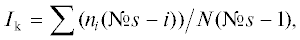
где: ni – число рыб в отсеке №i, №s – номер стартового отсека, i – номер отсека, N – число всех рыб в эксперименте.
Кроме того, для анализа и обобщения результатов в работе использованы исходные данные по контрольным особям, полученные нами в исследованиях физиолого-биохимических механизмов миграций кумжи (Salmo trutta), черноморской кумжи (Salmo trutta labrax) и радужной форели (Павлов и др., 2010, 2018, 2020а; Павлов Е. и др., 2016). Эти особи не были подвергнуты экспериментальному воздействию (гормональными препаратами или лишением доступа к убежищам) ни до начала, ни в течение голодания.
Для статистического анализа использовали непараметрический дисперсионный анализ (H‑критерий Краскела–Уоллиса) с использованием результатов каждого опыта и критерий Стьюдента для долей (Лакин, 1973), проведенный по суммарным данным для каждого вида. Как и любые соотношения, соотношения типов реореакции достоверно различаются, когда достоверно различаются частоты проявлений хотя бы одного из типов реореакции.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Непараметрический дисперсионный анализ (H-критерий Краскела–Уоллиса) показал, что голодание достоверно (p < 0.05) влияет на частоту проявления динамических типов реореакции (ПТР, ОТР), но не КТР (р = 0.46). Не выявлено достоверного совместного влияния двух факторов (вида рыб и длительности их голодания) на рассматриваемые показатели. То есть при голодании характер изменения соотношения типов реореакции сходен у рыб исследованных видов.
У неголодавших рыб (0-е сутки, табл. 1) преобладают динамические типы реореакции (83.3–96.7%), при этом частоты их проявления (ПТР и ОТР) примерно равны (р > 0.70, критерий Стьюдента для долей). Такое соотношение типов реореакции наблюдается у рыб с резидентным поведением, использующих динамический способ сохранения местоположения (Пономарева и др., 2017). Этому соответствует и близкое к нулю значение индекса контранатантности. В естественных условиях такое соотношение типов реореакции наблюдалось у немигрирующих рыб (Павлов и др., 2010, 2020б; Pavlov et al., 2010). Следовательно, у подопытных рыб до начала голодания не было миграционного состояния – в экспериментальной установке они проявляли резидентное поведение с динамическим способом сохранения местоположения.
Таблица 1.
Частота проявления типов реореакции в “Рыбоходе” при голодании у 4-х видов рыб
| Вид рыб | Сутки голодания |
ПТР | КТР | ОТР |
|---|---|---|---|---|
| D. rerio | 0 | 40.0 | 3.3 | 56.7 |
| 2 | 16.7 | 3.3 | 80.0 | |
| 5 | 31.7 | 1.7 | 66.7 | |
| 7 | 35.0 | 8.3 | 56.7 | |
| 10 | 31.7 | 0.0 | 68.3 | |
| 12 | 45.0 | 3.3 | 51.7 | |
| P. mykiss (=O. mykiss) | 0 | 45.0 | 15.0 | 40.0 |
| 2 | 28.7 | 10.0 | 61.3 | |
| 5 | 23.3 | 13.3 | 63.3 | |
| 7 | 40.0 | 21.7 | 38.3 | |
| 10 | 38.3 | 6.7 | 55.0 | |
| 12 | 46.7 | 1.7 | 51.7 | |
| L. delineatus | 0 | 48.3 | 3.3 | 48.3 |
| 2 | 33.3 | 3.3 | 63.3 | |
| 5 | 23.3 | 1.7 | 75.0 | |
| 7 | 33.3 | 3.3 | 63.3 | |
| 10 | 48.3 | 0.0 | 51.7 | |
| 12 | 40.0 | 0.0 | 60.0 | |
| C. auratus | 0 | 43.3 | 16.7 | 40.0 |
| 2 | 45.0 | 5.0 | 50.0 | |
| 5 | 36.7 | 5.0 | 58.3 | |
| 7 | 66.7 | 3.3 | 30.0 | |
| 10 | 51.7 | 10.0 | 38.3 | |
| 12 | 46.7 | 0.0 | 53.3 |
На вторые–пятые сутки голодания у всех исследованных рыб частота ОТР стала больше, чем ПТР (табл. 1). Такое соотношения типов реореакции в естественных условиях указывает на перемещение (миграцию или откочевку) рыб по течению (Pavlov et al., 2010; Павлов и др., 2010, 2020б; Звездин, 2016). Затем у рыб повышалась мотивация к движению против течения. В результате на 7-е–12-е сутки голодания у исследованных рыб соотношение типов реореакции приближалось к своему начальному значению, и разница в частотах ПТР и ОТР вновь стала недостоверной (табл. 1).
Индекс контранатантности (Ik) дает наиболее наглядную оценку динамики изменения поведения рыб, связанного с миграциями. В табл. 2 представлены как результаты данной работы, так и ранее полученные на особях радужной форели, кумжи и ее подвида – черноморской кумжи (Павлов и др., 2010, 2018, 2020а; Павлов Е. и др., 2016). Для всех рыб характерно в начале голодания увеличение мотивации к движению по течению, а затем – против течения. К концу эксперимента Ik приближается к исходным значениям.
Таблица 2.
Средние по суткам голодания индексы контранатантности у исследованных рыб
| Вид рыб | Длительность голодания, сут. | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 5 | 7 | 10 | 12 | |
| L. delineatus | –0.01 | –0.28 | –0.47 | –0.20 | 0.00 | –0.06 |
| D. rerio | –0.14 | –0.59 | –0.23 | –0.15 | –0.05 | 0.00 |
| P. mykiss (=O. mykiss) | 0.00 | –0.38 | –0.21 | –0.11 | –0.22 | 0.01 |
| C. auratus | 0.10 | –0.14 | 0.03 | 0.19 | 0.08 | –0.03 |
| P. mykiss (=O. mykiss) (Павлов и др., 2020а) | –0.14 | –0.57 | –0.30 | –0.23 | –0.25 | 0.05 |
| P. mykiss (=O. mykiss) (Павлов Е. и др., 2016) | 0.13 | –0.21 | –0.20 | –0.03 | –0.15 | 0.00 |
| S. truttalabrax (Павлов и др., 2010) | –0.08 | –0.18 | –0.22 | –0.09 | –0.01 | – |
| S. trutta (Павлов и др., 2018) | –0.43 | –0.62 | –0.68 | –0.58 | –0.42 | – |
Дисперсионный анализ показал, что Ik, как и соотношение типов реореакции зависит: а) от длительности голодания (р = 0.002), что указывает на изменение средней (по видам) величины Ik в течение опыта; б) от вида рыб (р = 0.006), т.е. средняя (за все сутки голодания) величина Ik различна у разных видов (рис. 1а). Совместное влияние двух факторов (вид рыб и длительность голодания) оказалось недостоверным (р = 0.26), что свидетельствует о сходной динамике изменения Ik у различных видов рыб при голодании (рис. 1б).
Рис. 1.
Изменение индекса контранатантности (Ik) при голодании карповых и лососевых рыб с резидентным поведением до начала голодания. (а) – регрессионные кривые по видам: 1 – C. auratus, 2 – P. mykiss (=O. mykiss) (Павлов Е. и др., 2016), 3 – S. trutta labrax (Павлов и др., 2010), 4 – P. mykiss (=O. mykiss), 5 – L. delineatus, 6 – P. mykiss (=O. mykiss) (Павлов и др., 2020а), 7 – D. rerio, 8 – S. trutta (Павлов и др., 2018); (б) – общая схема динамики Ik.
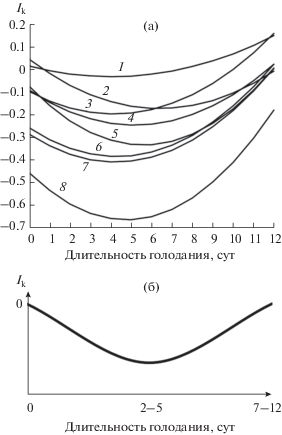
У исследованных рыб в разных масштабах наблюдается сходная динамика Ik. От начала голодания ко 2–5 суткам величина Ik уменьшается, что указывает на преимущественное движение рыб по течению. После достижение минимума Ik, от 2–5 сут к 7–12 сут голодания рыбы начинают больше двигаться против течения, и величина Ik приближается к исходной. В этот период зафиксированы минимальные значения частоты КТР (табл. 1), что свидетельствует о большей подвижности рыб на 7–12 сутки голодания.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате анализа полученных материалов у рыб с исходным резидентным поведением (без мотивации к миграции) выявлены два этапа поведенческого ответа на голодание. На первом этапе (2–5 сут голодания) у рыб усиливается мотивация к движению по течению, что в естественных условиях обеспечивает их покатную миграцию или откочевку по течению. То есть на первом этапе ответом рыб на голодание является поиск пищи с минимальными затратами энергии. Известно, что недостаток пищи мотивирует молодь рыб к покатным миграциям (Павлов, 1979; Olsson et al., 2006; Павлов Е. и др., 2007; Flecker et al., 2010; Ferguson et al., 2019). На втором этапе (7–12 сут голодания) начинает усиливаться мотивация к движению против течения. И это приводит к примерно одинаковому движению рыб по и против течения, при этом у них увеличивается подвижность. То есть на втором этапе ответ рыб на голодание заключается в поиске корма в обоих направлениях относительно течения.
Сроки и характер изменения мотивационной компоненты реореакции при отсутствии корма соответствуют срокам и характеру изменений других компонент реореакции. Например, время начала оптомоторной реакции у молоди ельца (Leuciscus leuciscus), не кормленной 48 ч, увеличивалось в 32 раза, а оценка силы реакции значительно снижалась. Аналогичные результаты были получены и на других видах рыб (Павлов, 1979). При голодании также изменяется и локомоторная компонента реореакции рыб. В экспериментах первоначальной реакцией на снижение рациона было повышение активности рыб – особи чаще двигались в потоке и реже отдыхали в специальных затишных зонах (Этинген, 1964; Sogard, Olla, 1996). Также на молоди рыб различных видов было показано, что их крейсерские скорости плавания уменьшаются на 1–6 сут голодания (MаcLeod, 1967; Павлов, 1979; Laurence, 1972; Faria et al., 2011). У осетровых (Acipenseridae) голодание приводило к снижению числа рыб, активно сопротивлявшихся течению, например, у молоди севрюги (Acipenser stellatus) через один день – до 49%, через 4 дня – до 15–25%, а через 6 дней – до 2% рыб (Павлов, 1979). Бросковые (Сабуренков, 1973) и критические (Faria et al., 2011) скорости течения для молоди рыб не изменяются при голодании до 30 и 3 сут соответственно.
Уменьшение крейсерских скоростей рыб некоторые авторы связывают со снижением выносливости при голодании (Faria et al., 2011), однако это объяснение, по нашему мнению, не всегда обосновано. Для личинок плотвы Rutilus rutilus было показано, что на показатели плавательной способности рыб существенное влияние оказывает наличие или отсутствие у них мотивации к покатной миграции. При дневной освещенности, когда в реке нет миграции плотвы, плавательная способность мигрантов была выше, чем у резидентов. А при ночной освещенности, когда в реке идет покатная миграция, плавательная способность мигрантов стала ниже, чем у резидентов (Павлов и др., 2007). На сеголетках плотвы было показано, что у рыб с мотивацией к покатной миграции снижается критическая скорость течения (Костин и др., 2016). Поэтому мы считаем, что снижение экспериментальных показателей плавательной способности рыб при начале голодания может быть вызвано появлением мотивации к движению по течению, а не только изменением их физиологических возможностей. Это согласуется с данными литературы, например, у молоди минтая Theragra chalcogramna переход от активного поискового поведения к снижению двигательной активности происходил на 30–54 сутки голодания (Sogard, Olla, 1996).
Ранее нами были получены материалы по влиянию голодания на соотношение типов реореакции у рыб с мотивацией к покатной миграции. На черноморской кумже были проведены исследования по изучению условий формирования жизненных (миграционных) стратегий (Павлов и др., 2010; Пономарева, 2014). Для этого вида генетически детерминирован скат в море части особей из популяции (Jonsson, Jonsson, 2011). При воздействии неблагоприятных условий содержания различной природы у черноморской кумжи начинала формироваться повышенная мотивация к покатной миграции. В этих исследованиях было показано, что у таких рыб наблюдали иной ответ на голодание – начиная со вторых суток монотонно увеличивалась мотивация рыб к движению по течению. Видимо, ответ особей на голодание во многом определяется их мотивацией к миграции, сформировавшейся перед началом голодания.
Таким образом, выявленные изменения мотивационной компоненты реореакции показали, что рыбы без мотивации к миграции сначала будут покидать участок обитания с минимальными затратами энергии – скатываться по течению. С увеличением длительности голодания рыбы начнут активный поиск корма, двигаясь как по, так и против течения. У особей с мотивацией к покатной миграции поведение существенно отличается – они будут постоянно скатываться вниз по течению.
Авторы благодарят Д.А. Прозорова за участие в проведении опытов, А.О. Касумяна (МГУ) за ценные замечания по тексту рукописи.
Сбор материала выполнен в 2018 г. при поддержке Программы Президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России”, его обработка, анализ и подготовка статьи при финансовой поддержке РНФ (грант 19-14-00015).
Настоящая статья не содержит каких-либо исследований с участием людей в качестве объектов исследований.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Список литературы
Аннотированный каталог круглоротых и рыб континентальных вод России / Под ред. Решетникова Ю.С. М.: Наука, 1998. 218 с.
Звездин А.О. Реореакция ранней молоди нерки Oncorhynchus nerka (Walb.) в период расселения с нерестилищ: Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 2016. 28 с.
Костин В.В., Звездин А.О., Павлов Д.С., Голованов В.К., Маврин А.С., Мартемьянов В.И., Капшай Д.С. Сравнение поведенческих и физиолого-биохимических показателей у сеголетков плотвы Rutilus rutilus (L.) из реки Ильдь и Ильдинского залива Рыбинского водохранилища // Биология внутренних вод. 2016. № 3. С. 79–88. https://doi.org/10.7868/S0320965216030128
Лакин Г.Ф. Биометрия. М.: Высшая школа, 1973. 343 с.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука, 1979. 319 с.
Павлов Д.С., Лупандин А.И., Костин В.В. Механизмы покатной миграции молоди речных рыб. М.: Наука, 2007. 213 с.
Павлов Д.С., Костин В.В., Пономарева В.Ю. Поведенческая дифференциация сеголеток черноморской кумжи Salmo trutta labrax: реореакция в год, предшествующий смолтификации // Вопр. ихтиологии. 2010. Т. 50. № 2. С. 251–261.
Павлов Д.С., Павлов Е.Д., Костин В.В., Ганжа Е.В. Реореакция молоди радужной форели при воздействии сурфагона // Онтогенез. 2016. Т. 47. № 2. С. 110–116. https://doi.org/10.7868/S0475145016020075
Павлов Д.С., Павлов Е.Д., Ганжа Е.В., Зарипов П.И., Костин В.В., Ручьёв М.А. Влияние сурфагона на реореакцию молоди кумжи Salmo trutta // Вопр. ихтиологии. 2018. Т. 58. № 6. С. 740–744. https://doi.org/10.1134/S004287521806019X
Павлов Д.С., Костин В.В., Звездин А.О., Прозоров Д.А., Подоляко С.А. Реореакция молоди некоторых карповых рыб (Cyprinidae) в период осенней контранатантной миграции // Вопр. ихтиологии. 2019. Т. 59. № 6. С. 716–723. https://doi.org/10.1134/S0042875219060122
Павлов Д.С., Павлов Е.Д., Ганжа Е.В., Костин В.В. Изменение реореакции и содержания тиреоидных гормонов в крови молоди радужной форели Oncorhynchus mykiss при голодании // Вопр. ихтиологии. 2020а. Т. 60. № 2. С. 229–234. https://doi.org/10.31857/S0042875220020186
Павлов Д.С., Паршина В.Ю., Костин В.В., Прозоров Д.А. Сравнение экспериментальных методов оценки мотивационной компоненты реореакции рыб (соотношения типов реореакции) // Вопросы ихтиологии. 2020б. Т. 60. №. 4. С. 643–652. https://doi.org/10.1134/S0032945220040153
Павлов Е.Д., Ганжа Е.В., Пономарева В.Ю., Костин В.В., Павлов Д.С., Во Тхи Ха. Влияние метилтестостерона на физиологическое состояние и реореакцию радужной форели Parasalmo mykiss (=Oncorhynchus mykiss) при неблагоприятных условиях содержания // Вопросы ихтиологии. 2016. Т. 56. № 6. С. 740–752.
Пономарева В.Ю. Поведенческие механизмы внутрипопуляционной дифференциации молоди некоторых лососевых рыб: Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 2014. 28 с.
Пономарева В.Ю., Павлов Д.С., Костин В.В. Разработка и апробирование методики исследования соотношения типов реореакции рыб в кольцевом гидродинамическом лотке // Биология внутренних вод. 2017. № 1. С. 100–108. https://doi.org/10.7868/S0320965217010156
Сабуренков Е.Н. Скорость и длительность плавания рыб как элементы их поведения: Автореф. дис. … канд. биол. наук. М.: ИЭМЭЖ, 1973. 23 с.
Этинген Р.М. О реакции рыб на течение в зависимости от степени накормленности // Вопросы ихтиологии. 1964. Т. 4. № 4. С. 624–631.
Faria A., Chícharo M.A., Gonçalves E.J. Effects of starvation on swimming performance and body condition of pre-settlement Sparus aurata larvae // Aquat. Biol. 2011. V. 12. P. 281–289. https://doi.org/10.3354/ab00345
Ferguson A., Reed T.E., Cross T.F. et al. Anadromy, potamodromy and residency in brown trout Salmo trutta: the role of genes and the environment // J. Fish Biol. 2019. P. 1. https://doi.org/10.1111/jfb.14005
Flecker A.S., McIntyre P.B., Moore J.W., Anderson J.T., Taylor B.W., Hall R.O. Migratory fishes as material and process subsidies in riverine ecosystems // Am. Fish. Soc. Symp. 2010. V. 73. P. 559–592.
Gaffron M. Untersuchungen über das Bewegungssehen bei Libellenlarven, Fliegen und Fischen // Z. f. vergl. Physiologie. 1934. V.20. P. 299–337. https://doi.org/10.1007/BF00361440
Jonsson B., Jonsson N. Ecology of atlantic salmon and brown trout: habitat as a template for life histories // Fish Fisheries Ser. 2011. V. 33.
Johnston M.E., Kelly J.T., Lindvall M.E. et al. Experimental evaluation of the use of vision and barbels as references for rheotaxis in green sturgeon // J. Exp. Mar. Biol. Ecol. 2017. V. 496. P. 9. https://doi.org/10.1016/j.jembe.2017.04.002
Harden Jones F.R. The reaction of fish to moving backgrounds // J. Exp. Biol. 1963. V. 40. № 3. P. 437–446.
Laurence G.C. Comparative swimming abilities of fed and starved larval largemouth bass (Micropterus salmoides) // J. Fish Biol. 1972. V. 4. № 1. P. 73–78. https://doi.org/10.1111/j.1095-8649.1972.tb05654.x
Maclean J.A., Gee J.H. Effect of temperature on movements of pre-spawning brook sticklebacks, Culaea inconstans, in the Roseau River, Manitoba // J. Fish. Res. Board Can. 1971. V. 28. P. 919. https://doi.org/DOI:10.1139/f71-133
MacLeod J.C. A new apparatus for measuring maximum swimming speeds of small fish // J. Fish. Res. Board Can. 1967. V. 24. № 6. P. 1241–1252. https://doi.org/10.1139/f67-106
Olsson I.C., Greenberg L.A., Bergman E., Wysujack K. Environmentally induced migration: The importance of food // Ecology Letters. 2006. V. 9. P. 645–651. https://doi.org/10.1111/j.1461-0248.2006.00909.x
Pavlov D.S., Kostin V.V., Zvezdin A.O., Ponomareva V.Yu. On methods of determination of the rheoreaction type in fish // J. Ichthyology. 2010. V. 50. № 11. P. 977. https://doi.org/10.1134/s0032945210110020
Sogard S.M., Olla B.L. Food deprivation affects vertical distribution and activity of a marine fish in a thermal gradient: potential energy-conserving mechanisms // Mar. Ecol. Prog. Ser. 1996. 133. P. 43–55. https://doi.org/10.3354/meps133043
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая