

Известия РАН. Серия биологическая, 2022, № 1, стр. 46-53
Пространственно-типологическая структура и организация населения птиц Алданского нагорья
Е. В. Шемякин 1, Л. Г. Вартапетов 2, *
1 Институт биологических проблем криолитозоны СО РАН
677980 Якутск, пр. Ленина, 41, Россия
2 Институт систематики и экологии животных СО РАН
630091 Новосибирск, ул. Фрунзе, 11, Россия
* E-mail: lvartapetov@yandex.ru
Поступила в редакцию 26.04.2020
После доработки 19.06.2020
Принята к публикации 19.06.2020
- EDN: NZMCBQ
- DOI: 10.31857/S1026347022010127
Аннотация
На основе многолетних учетов изучено население птиц одного из наименее обследованных регионов Якутии – Алданского нагорья. Впервые составлена наиболее полная и подробная иерархическая классификация сообществ птиц. Выявлена пространственно-типологическая структура населения птиц, оценена значимость факторов среды в формировании орнитокомплексов. Установлены региональные особенности формирования орнитокомплексов.
Алданское нагорье – северо-восточная часть Байкальской горной страны, его территория занимает 189 тыс. км2. Этот регион характеризуется сложной структурой высотной поясности и значительным ландшафтным разнообразием. Рассматриваемая территория отличается широким диапазоном природных условий, которые определяются сложным сочетанием горных хребтов и межгорных котловин. Алданское нагорье соседствует с таежными равнинными пространствами Лено-Алданского плато с северо-запада и горными территориями Верхоянского хребта с северо-востока, а также – Станового хребта с юга, без сколько-нибудь значительных зоогеографических барьеров, что обусловливает высокий уровень его биологического разнообразия.
До недавнего времени Алданское нагорье, особенно его верхние высотные пояса, гольцово-тундровый и подгольцово-редколесный, оставались мало обследованными. Орнитокомплексы этих поясов стали изучать в наибольшей степени после публикации наших первых результатов (Вартапетов и др., 2012). Поэтому данная работа в значительной степени дополняет и расширяет опубликованные ранее представления о классификации и пространственно-типологической структуре населения птиц этого региона.
Результат исследования явяляется значимым не только с фундаментальных позиций орнитогеографии и синэкологии, но и для решения региональных эколого-географических проблем, в том числе связанных с сохранением биоразнообразия сообществ птиц. Количественные оценки населения птиц, в том числе выполненные на ландшафтной основе, служат для развития международных систем орнитологического мониторинга, в основном направленных на выявление популяционных трендов гнездящихся и мигрирующих птиц (Gregory, 2000; Svensson, 2000; Sanderson et al., 2006; Gregory et al., 2007). В России ландшафтно-типологический подход в орнитологическом мониторинге направлен на решение общих задач экологического мониторинга, а также сохранения и восстановления биологических ресурсов и биоразнообразия (Гармаев и др., 2016).
Основные задачи данной работы – на основе иерархической классификации населения птиц Алданского нагорья определить главные направления его территориальных изменений (пространственно-типологическую структуру), а также оценить и сопоставить значимость основных факторов среды для формирования населения птиц или пространственную организацию орнитосообществ. Кроме этого, установить региональную специфику населения птиц Алданского нагорья в сопоставлении с другими равнинными и горными природно-географическими областями.
МАТЕРИАЛЫ И МЕТОДЫ
Пространственная неоднородность фауны и населения птиц Алданского нагорья выявлена на основе собственных неопубликованных данных, а также с использованием материалов из банка данных коллективного пользования Института систематики и экологии животных СО РАН, ранее опубликованных авторами и другими специалистами (Егоров и др., 2002; Исаев и др., 2006; Вартапетов и др., 2008, 2012; Шемякин и др., 2014, 2016). Использованы результаты учетов птиц в первой половине лета (с конца мая до середины июля). В этот период завершается пролет и прилет птиц, а гнездящиеся виды уже приступили к размножению. Таким образом, результаты маршрутных учетов в этот период позволяют описать относительно стабильное гнездовое население птиц. Доля пролетных особей (в основном, гусеообразных и ржанкообразных), в этот период незначительна.
Маршрутные учеты проведены без ограничения ширины трансекта с последующим раздельным пересчетом полученных данных на площадь по средним групповым дальностям обнаружения, рассчитанным интервальным методом (Равкин, Ливанов, 2008). В период с 2000 по 2016 гг. пешими и водными маршрутами на территории Алданского нагорья пройдено 3272 км. Всего проанализировано 84 варианта населения птиц. Вариант населения – это список видов с указанием обилия каждого из них для каждого из обследованных местообитаний (показатели усреднены за первую половину лета).
Классификация населения проведена с помощью одного из методов кластерного анализа (Трофимов, Равкин, 1980) с использованием коэффициента сходства Жаккара (Jaccard, 1902) для количественных признаков (Наумов, 1964). Суть метода сводится к разделению совокупности рассматриваемых объектов (вариантов населения) на незаданное число групп по степени сходства каждой пробы со всеми остальными. Крупные группы орнитокомплексов подразделяли на подгруппы, пока для каждой из них можно было установить природный режим, определяющий их выделение (Вартапетов, 1998). Результаты первого разделения считали делением на типы населения, а последующих – делениями на подтипы и классы. В итоге была составлена иерархическая классификация населения птиц. Основное назначение таких классификаций – выявить взаимосвязи между признаками фаун или животного населения и факторами окружающей среды (Харвей, 1974; Hengeveld, 1990).
Пространственно-типологическая структура построена по матрице средних коэффициентов сходства на уровне подтипа населения. Порог значимости связей определяли так, чтобы полученная схема наилучшим образом отображала выявленные тренды орнитокомплексов и ее можно было построить в двухмерном пространстве. Оценка силы связи факторов среды и неразделимых сочетаний (природно-антропогенных режимов) с пространственной неоднородностью населения птиц проведена с помощью линейной качественной аппроксимации по выделенным градациям факторов (Равкин и др., 1978).
К фоновым отнесены виды, доля которых в населении птиц по обилию составляла не менее одной особи на 1 км2 (Кузякин, 1962). Лидирующими считали первые пять видов, преобладающих по обилию (Равкин, Ливанов, 2008). Названия типов фаун даны по Б.К. Штегману (1938) с некоторыми дополнениями. Видовые названия птиц приведены по Л.С. Степаняну (2003), с некоторыми дополнениями из “Списка птиц Российской Федерации” (Коблик и др., 2006). Для расчета биомассы использованы сведения из монографии “Птицы Советского Союза” (1951–1954). Показатели рассчитаны как суммарная сырая масса особей, приходящих на 1 км2.
КЛАССИФИКАЦИЯ НАСЕЛЕНИЯ
В приведенной ниже классификации для каждого из выделенных таксонов приведено по пять лидирующих по обилию (преобладающих) видов с указанием их доли (в %) в сообществах, а также суммарное обилие/биомасса и число встреченных/фоновых видов. Кроме того, в классификацию включены сведения о фаунистическом составе населения с указанием доминирующих типов фауны и их доли по количеству особей (в %).
1. Лесной тип населения (лесов и гарей) преобладает по обилию – юрок (10), пятнистый конек (8), буроголовая гаичка (7), корольковая пеночка (6), зеленая пеночка (5); плотность населения/биомасса – 201 особь/км2/10 кг/км2; всего видов/фоновых видов – 114/38; доля по количеству особей, в %: сибирского типа фауны – 65, китайского – 24, транспалеарктов – 6 и европейского – 4.
1.1. Подтип населения приречных лесов: юрок (14), зеленая пеночка (8), корольковая пеночка (6), буроголовая гаичка (6), пятнистый конек (5); 228/11; 88/39; сибирского типа фауны – 65, китайского – 24, транспалеарктов – 6 и европейского – 5.
Классы населения:
1.1.1 – ивово-березово тополевых пойменных лесов: юрок (28), зеленая пеночка (11), пятнистый конек (8), седоголовая овсянка (5), бурая пеночка (4); 233/8; 45/31; китайского типа фауны – 31, сибирского – 6, транспалеарктов – 4;
1.1.2 – надпойменных лиственнично-елово-березовых лесов: буроголовая гаичка (8), корольковая пеночка и зеленая пеночка (по 7), юрок (6), синехвостка (5); 226/12; 83/41; сибирского типа фауны – 66, китайского – 20, транспалеарктов и европейского по 7.
1.2. Подтип населения не приречных лесов: пятнистый конек (10), буроголовая гаичка (8), корольковая пеночка и юрок (по 7), синехвостка (5); 174/8; 82/35; сибирского типа фауны – 68, китайского – 23, европейского – 5, транспалеарктов – 4.
Классы населения:
1.2.1 – сосново-лиственничных лесов: пятнистый конек (14), юрок (13), седоголовая овсянка (12), пеночка-таловка (7), корольковая пеночка (6); 251/8; 34/30; сибирского типа фауны – 63, китайского – 31, европейского и транспалеарктов по 3;
1.2.2 – лиственнично-сосновых лесов: корольковая пеночка и буроголовая гаичка (11), пятнистый конек (10), овсянка-ремез (8), пеночка-зарничка (5); 194/11; 68/28; сибирского типа фауны – 70, китайского – 18, европейского – 8, транспалеарктов – 4;
1.2.3 – лиственничных лесов: синехвостка (11), дубровник (10), зеленая пеночка (9), рыжая овсянка и соловей-красношейка (по 7); 160/8; 36/28; сибирского типа фауны – 63; китайского – 31, транспалеарктов – 5;
1.2.4 – сосновых лесов: сероголовая гаичка (18), овсянка-ремез (16), малая мухоловка (10), буроголовая гаичка (9), пятнистый конек (6); 115/6; 43/19; сибирского типа фауны – 76; китайского – 12, транспалеарктов – 9, европейского – 3;
1.2.5 – еловых лесов: юрок (20), буроголовая гаичка (12), корольковая пеночка (9), сибирская завирушка и пеночка-таловка (по 8); 77/2; 22/20; сибирского типа фауны – 80, китайского – 13, европейского – 4.
1.3. Подтип населения гарей по сосново-лиственничным и лиственничным лесам: пятнистый конек (12), юрок и пеночка-зарничка (по 9), буроголовая гаичка и черноголовый чекан (по 6); 224/18; 79/40; сибирского типа фауны – 60; китайского – 24, транспалеарктов – 11.
Классы населения:
1.3.1 – молодых (свежих) гарей: пятнистый конек (33), буроголовая гаичка и трехпалый дятел (по 9), овсянка-ремез (7), соловей-красношейка (5); 108/9; 37/23; сибирского типа фауны – 53, китайского – 37, транспалеарктов – 3;
1.3.2 – старых и средневозрастных гарей: пятнистый конек и пеночка-зарничка (по 10), юрок (9), черноголовый чекан (7), буроголовая гаичка (6); 252/20; 71/40; сибирского типа фауны – 61, китайского – 23, транспалеарктов – 12.
2. Редколесный тип населения (лиственничных редколесий в сочетании с марями, кедровым стлаником и кустарниками, с участками луговин): бурая пеночка (14), пятнистый конек, зеленоголовая трясогузка, пеночка-таловка и буроголовая гаичка (по 8); 141/5; 84/30; сибирского типа фауны – 55, китайского – 29, транспалеарктов – 9, европейского – 3.
2.1. Подтип населения ерниковых редколесий: бурая пеночка (18), зеленоголовая трясогузка (13), юрок, седоголовая овсянка и пеночка-таловка (по 5); 145/5; 72/26; сибирского типа фауны – 50, китайского – 34, транспалеарктов – 10, европейского – 3;
2.2. Подтип населения редколесий с зарослями кедрового стланика и кустарников: бурая пеночка и буроголовая гаичка (по 12), пеночка-таловка и пятнистый конек (по 10), соловей-красношейка (6); 161/4; 47/30; сибирского типа фауны – 63, китайского – 29, европейского – 4, транспалеарктов – 3;
2.3. Подтип населения мезотрофных кустарничково-осоково-сфагновых болот с участками редколесий: черноголовый чекан (30), белокрылый клест (17), зеленоголовая трясогузка (16), пятнистый конек (9), соловей-красношейка (7); 144/10; 25/17; сибирского типа фауны – 48, транспалеарктов – 38, китайского – 12;
2.4. Подтип населения зарослей кедрового стланика и кустарников с участками луговин и каменистых осыпей: бурая пеночка (15), пятнистый конек (13), соловей-красношейка (9), рогатый жаворонок и зеленоголовая трясогузка (по 8); 90/4; 33/21; сибирского типа фауны – 49, китайского – 31, арктического – 11, транспалеарктов и европейского по 3.
3. Горно-тундровый тип населения: альпийская завирушка (32), зеленоголовая трясогузка и горный конек (по 14), каменка (12), сибирский горный вьюрок (10); 54/3; 16/7; тибетского типа фауны – 46, сибирского – 17, транспалеарктов – 14, арктического – 11, неясного происхождения – 10.
4. Марево-луговой тип населения: бурая пеночка (19), певчий сверчок (16), обыкновенная чечевица (11), седоголовая овсянка (9), пятнистый сверчок (5); 328/18; 62/34; китайского типа фауны – 47, сибирского – 22, монгольского – 16.
4.1. Подтип населения закустаренных марей: бурая пеночка (27), певчий сверчок (23), пятнистый сверчок (7), дубровник (6), азиатский бекас (5); 466/20; 41/34; китайского типа фауны – 47, сибирского и монгольского – по 23, транспалеарктов – 7;
4.2. Подтип населения пойменных лугов, кустарников и озер: бурая пеночка (17), певчий сверчок (14), чечевица (11), толстоклювая камышевка (8), азиатский бекас (6); 268/19; 42/33; китайского типа фауны – 46, сибирского – 25, транспалеарктов – 15, монгольского – 14;
4.3. Подтип населения антропогенных лугов: чечевица (25), седоголовая овсянка (21), горная трясогузка (11), соловей-красношейка и белая трясогузка (по 7); 219/16; 25/22; китайского типа фауны – 50, транспалеарктов – 23, сибирского – 18, европейского – 7, монгольского – 3.
5. Селитебный тип населения (поселков): домовый воробей (35), воронок (21), белая трясогузка (19), бурая пеночка (5), чечевица (3); 301/24; 21/18; транспалеарктов – 80, китайского типа фауны – 10, европейского – 6, сибирского – 3.
6. Водно-околоводный тип населения (рек, озер и их берегов): перевозчик (17), речная крачка (10), горная трясогузка и гоголь (по 8), сизая чайка (7); 55/20; 58/16; транспалеарктов – 61, сибирского типа фауны – 27, голарктов – 8.
6.1. Подтип населения крупных рек: речная крачка (31), сизая чайка (15), озерная чайка (12), перевозчик (9), кряква (3); 3/11; 32/0; транспалеарктов – 70, голарктов – 15, сибирского типа фауны – 8, европейского – 4;
6.2. Подтип населения средних и малых рек: перевозчик – 41, гоголь и горная трясогузка (по 16), воронок – 6, речная крачка – 3; 63/17; 34/8; транспалеарктов – 74, сибирского типа фауны – 20, голарктов – 3;
6.3. Подтип населения озер и стариц: речная крачка (13), зеленоголовая трясогузка (11), сизая чайка (10), кряква (8), большой улит (7); 63/27; 45/16; транспалеарктов – 53, сибирского типа фауны – 31, голарктов – 11, арктического – 3.
Рассмотренная классификация отражает различия в населении птиц незастроенной и застроенной суши, а также рек и озер. Незастроенные местообитания представлены четырьмя типами, десятью подтипами и девятью классами, застроенные – одним типом, а водно-околоводные – одним типом и тремя подтипами населения. При дифференциации типов населения птиц прослежено воздействие облесенности, высотной поясности и продуктивности биоценозов, которые определяют выделение первых трех типов. Формирование четвертого типа населения (марей и лугов) связано с воздействием увлажнения и закустаренности. Выделение пятого типа связано с антропогенным воздействием, проявляющемся в основном как застроенность, а выделение шестого типа определяется наличием водоемов. На более низком классификационном уровне выделение подтипов населения птиц определяется также совместным воздействием облесенности, высотной поясности, различий в рельефе, продуктивности и влияния пожаров. Кроме того, в формировании подтипов орнитокомплексов (2.1; 2.2; 2.3; 2.4 и в подтипах 4.1; 4.2 и 4.3) заметно воздействие заболоченности, закустаренности и антропогенного воздействия, а в подтипах (6.1; 6.2 и 6.3) – размеров водотоков и различий между реками и озерами.
Лесной тип населения наиболее дробный. Только он, кроме подтипов, подразделяется и далее – на классы. Он занимает наиболее широкий набор местообитаний в связи с обширным распространением на территории Алданского нагорья лесных сообществ и их формаций, но при этом определяющим становится не только разнообразие лесообразующих пород, но и уровень абсолютных высот. Так, с ростом абсолютных высот местности класс приречных пойменных лесов (1.1.1), сменяется таковым на склонах долин и надпойменных террас (класс 1.1.2), далее – на водоразделах (классы 1.2.1–1.2.5). Изменение орнитокомплексов в этом ряду связано с породным составом лесных растительных формаций. На приречных участках он характеризуется участием березы, ели, лиственницы, тополя и высоких ивовых кустарников. На склонах долин и их надпойменных террасах преобладают лиственничные и сосновые древостои, а на водоразделах – лиственничные леса с участием ели. Кроме того, здесь представлены классы орнитокомплексов гарей, которые разделены по возрасту (класс 1.3.1 и 1.3.2).
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ СТРУКТУРА ЛЕТНЕГО НАСЕЛЕНИЯ ПТИЦ АЛДАНСКОГО НАГОРЬЯ
Пространственно-типологическая структура орнитокомплексов выявлена на уровне подтипов населения птиц при пороге значимости связей в 10%. На схеме вертикальный ряд отражает изменения в облесенности и высотной поясности, направленный снизу вверх от лесных местообитаний через редколесья к горным тундрам (рис. 1). В приречных лесах (подтип 1.1), отмечены наибольшие видовое богатство и плотность населения птиц, которые последовательно уменьшаются с увеличением абсолютных высот. В пределах орнитокомплесов лесов и редколесий (подтипы 1.2 и 2.2) это уменьшение относительно невелико, но становится наиболее заметным при переходе к сообществам стлаников и кустарников (подтип 2.4) и особенно – горных тундр (тип 3). В лесных сообществах лидируют – пятнистый конек, буроголовая гаичка, корольковая пеночка и юрок, в редколесьях и кустарниках – пеночки: бурая и таловка, пятнистый конек и соловей-красношейка, а в горных тундрах – альпийская завирушка, зеленоголовая трясогузка, горный конек и каменка. Таким образом, с подъемом вверх лесные виды постепенно сменяются кустарниковыми, а последние – арктоальпийскими. Почти повсеместно сохраняется китайско-сибирский фаунистический состав населения птиц, и только в горных тундрах он становится сибирско-тибетским. Отклонение от вертикального ряда (подтип 1.3) связано с воздействием пожаров. На гарях продолжают доминировать лесные виды (пятнистый конек, юрок, пеночка-зарничка), а видовое богатство и плотность населения птиц лишь немного меньше, чем в приречных лесах.
Рис. 1.
Пространственно-типологическая структура летнего населения птиц Алданского нагорья. Условные обозначения. Орнитокомплексы: 1 – лесов, 2 – редколесий, 3 – горных тундр, 4 – марей и лугов, 5 – поселков, 6 – водоемов и водотоков. Цифрами внутри значков показаны номера типов и подтипов населения птиц, а у связей между значками – межклассовое сходство. Сплошные линии – значимое сверхпороговое сходство, прерывистые – максимальное при отсутствии значимого. Для каждого типа и подтипа населения птиц указаны лидирующие виды (первые три по обилию), среднее суммарное обилие птиц (особей/км2) и (через косую черту) биомасса (кг/км2), далее общее количество и число фоновых видов. Стрелками показаны основные направления пространственной изменчивости сообществ птиц и определяющие их факторы среды.
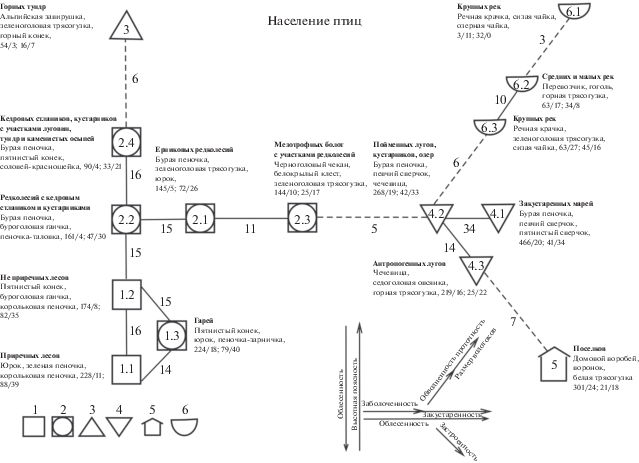
Горизонтальный ряд последовательно образуют сообщества птиц редколесий, мезотрофных болот, пойменных лугов и закустаренных марей (подклассы 2.2–4.1). Этот ряд формируется при воздействии заболоченности (от сообществ редколесий к таковым мезотрофных болот) и далее закустаренности – при переходе от орнитокомплексов мезотрофных болот к таковым в лугах, кустарниках и далее к закустаренным марям. При этом плотность населения птиц резко возрастает при переходе от орнитосообществ редколесий к таковым пойменных лугов и марей за счет обитателей кустарников и высокотравий (бурая пеночка, певчий и пятнистый сверчки, обыкновенная чечевица). Изменяется и фаунистический состав населения птиц: в редколесьях и на мезотрофных болотах он китайско-сибирский, а на лугах и в закустаренных марях – сибирско-китайский, при высоком участии транспалеарктов в лугово-болотных местообитаниях.
От сообществ пойменных лугов, кустарников и озер начинаются два диагональных ряда изменений. В нижнем из них значима связь орнитокомплексов пойменных и антропогенных лугов, которые в свою очередь имеют запороговое сходство с населением поселков. Лидирующие виды в населении антропогенных лугов представлены чечевицей, седоголовой овсянкой и горной трясогузкой, а в поселках преобладают домовый воробей, воронок и белая трясогузка. Верхний ряд образуют сообщества птиц водно-околоводных местообитаний. В этом ряду происходит снижение плотности населения от пойменных лугов и кустарников к озерам и небольшим рекам с минимумом на крупных реках. Это сопровождается сменой лидирующих видов сначала на речную крачку, зеленоголовую трясогузку и сизую чайку по берегам озер и стариц, на средних и малых реках их сменяют перевозчик, гоголь и горная трясогузка, а затем на крупных реках снова в качестве лидеров появляются речная крачка и сизая чайка. Таким образом, основные тренды в населении птиц Алданского нагорья связаны с высотной поясностью, облесенностью, заболоченностью и закустаренностью, а также с застроенностью и обводненностью. Кроме того, прослежено влияние проточности и размера акваторий.
При сопоставлении пространственно-типологической структуры населения птиц Алданского нагорья и Алтая (Цыбулин, 2009) установлено, что большинство перечисленных трендов характерно и для Алтая, и, по-видимому, для остальных гор Северной Евразии. Однако только для Алданского нагорья характерны такие структурообразующие факторы, как заболоченность и закустаренность. Это определяется наличием обширных мезотрофных болот в межгорных котловинах и закустаренных марей в его предгорьях. К общим трендам неоднородности населения птиц изучаемой территории и Центрально-Якутской равнины (Вартапетов, Ларионов, 2014) относятся воздействие облесенности, заболоченности, закустаренности, увлажненности, застроенности и обводненности. При этом только на Центрально-Якутской равнине выделены тренды, связанные с зональностью и с пойменным режимом рек, а только на Алданском нагорье – изменения орнитокомплексов, которые определяются высотной поясностью.
ПРОНСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ НАСЕЛЕНИЯ ПТИЦ АЛДАНСКОГО НАГОРЬЯ
По результатам кластерного анализа на Алданском нагорье выделено 12 значимых факторов среды, определяющих территориальную изменчивость орнитокомплексов. При оценке силы и общности их влияния на формирование орнитокомплексов природные и антропогенные факторы разделены. Основным фактором среды, влияющим на неоднородность летнего населения птиц, следует считать облесенность местообитаний (табл. 1). Менее значимы по доле учтенной дисперсии состав лесообразующих пород, продуктивность биоценозов, рельеф местности и обводненность. Еще меньше связь с абсолютными высотами и высотной поясностью, в связи с взаимопроникновением соседних поясов и широким распространением кедрового стланика от нижнего лесного пояса до верхнего тундрового. Наименьшее значение имеют закустаренность, наличие стлаников и заболоченность. Антропогенные факторы (наличие гарей и застроенность) мало значимы по сравнению с природными. Все перечисленные факторы совместно учитывают 59% дисперсии, что соответствует множественному коэффициенту корреляции, равному 0.77%.
Таблица 1.
Оценка силы и общности связи факторов среды и неоднородности летнего населения птиц Алданского нагорья
| Фактор, режим | Учтенная дисперсия, % |
|---|---|
| Природные факторы | 52 |
| Облесенность | 35 |
| Состав лесообразующих пород | 16 |
| Рельеф | 15 |
| Продуктивность (кормность) | 15 |
| Обводненость | 14 |
| Абсолютные высоты | 9 |
| Высотная поясность | 7 |
| Закустаренность | 5 |
| Наличие стлаников | 4 |
| Заболоченность | 1 |
| Антропогенные факторы: | 5 |
| Застроенность | 3 |
| Наличие гарей | 2 |
| Все факторы | 53 |
| Режимы (классификационные и структурные) | 38 |
| Все факторы и режимы | 59 |
| Множественный коэффициент корреляции | 0.77 |
При сравнении оценки силы и общности связи факторов среды с неоднородностью населения птиц Алданского нагорья и Алтая, выявлено некоторое сходство этих связей, которое сводится к значимому влиянию облесенности, обводненности и относительно слабому антропогенному воздействию. Кроме того, на Алтае, по мнению С.М. Цыбулина (2009), формирование орнитокомплексов существенно зависит от сложных комплексных факторов: тип растительного покрова, гидротермический режим, ландшафтная дифференциация. На Алданском нагорье вместо них мы выделяем более простые и легче объясняемые факторы: рельеф, продуктивность, абсолютные высоты и высотная поясность, но это скорее различия трактовок, а не реальных причин формирования орнитокомплексов. На Центрально-Якутской равнине по сравнению с Алданским нагорьем территориальная неоднородность населения птиц в большей степени зависит от обводненности и, особенно – от антропогенных факторов, связанных с выпасом, сенокошением и пирогенными сукцессиями лесной растительности (Вартапетов, Ларионов, 2014, Вартапетов и др. 2016, 2018).
ЗАКЛЮЧЕНИЕ
Два основных тренда пространственно-типологической структуры сообществ птиц Алданского нагорья определяют высотная поясность и развитие кустарниковой растительности, а также снижение облесенности в обоих направлениях. При подъеме вверх, снижении теплообеспеченности и смене высотных поясов происходит обеднение орнитокомплексов по числу видов и особей. При увеличении закустаренности и заболоченности видовое богатство населения птиц тоже уменьшается, но их суммарное обилие возрастает.
Пространственную неоднородность населения птиц Алданского нагорья определяет преимущественно облесенность местообитаний. Менее значимы другие “равнинные” факторы: состав лесообразующих пород, продуктивность и обводненность. Еще меньше сила и общность связи с “горными” факторами: абсолютные высоты, высотная поясность, наличие зарослей кедрового стланика. Из антропогенных факторов, которые в 10 раз менее значимы, чем перечисленные природные, заметно влияние наличия гарей и застроенности. Региональные особенности пространственной организации населения птиц Алданского нагорья – большая значимость закустаренности и заболоченности в формировании орнитокомплексов по сравнению с Алтаем, а по сравнению с Центрально-Якутской равниной – меньшее значение обводненности и особенно – антропогенного воздействия. Видовое богатство и плотность населения птиц Алданского нагорья меньше таковых на Северном Урале, в средней тайге Западной Сибири, и примерно такое же, что и в среднесибирской средней тайге. Это определяется усилением континентальности климата, снижением продуктивности биоценозов, “выпадением” темнохвойных из состава лесообразующих пород и преобладанием криогенных ландшафтов.
Работа выполнена в рамках государственного задания по проекту № 0376-2019-0004 АААА-А17-117020110058-4 “Структура и динамика популяций и сообществ животных холодного региона Северо-Востока России в современных условиях глобального изменения климата и антропогенной трансформации северных экосистем: факторы, механизмы, адаптации, сохранение” и поддержана проектом РФФИ (17-04-00088а).
Список литературы
Вартапетов Л.Г. Птицы северной тайги Западно-Сибирской равнины. Новосибирск: Наука, 1998. 327 с.
Вартапетов Л.Г., Егоров Н.Н., Дегтярев В.Г., Исаев А.П. Летнее население птиц долины нижнего течения р. Мая // Сиб. экол. журн. 2008. № 1. С. 161–170.
Вартапетов Л.Г., Исаев А.П., Ларионов А.Г., Егоров Н.Н. Классификация и структура населения птиц Алданского нагорья // Поволж. экол. журн. 2012. № 2. С. 157–164.
Вартапетов Л.Г., Ларионов А.Г. Классификация и пространственно-типологическая структура населения птиц Центрально-Якутской равнины // Успехи современ. биологии. Т. 134. № 5. 2014. С. 519–528.
Вартапетов Л.Г., Ларионов А.Г., Егоров Н.Н. Пространственное разнообразие населения птиц средней тайги Среднесибирского плоскогорья // Сиб. экол. журн. 2016. № 1. С. 13–23.
Вартапетов Л.Г., Железнова Т.К., Юдкин В.А. Пространственная организация населения птиц средней тайги Западно-Сибирской равнины // Известия РАН. Сер. биол. 2018. № 1. С. 71–78.
Гармаев Е.Ж., Намжилова Л.Г., Ананин А.А., Бешенцев А.Н. Разработка программы мониторинга биоразнообразия на особо охраняемых природных территориях в бассейне озера Байкал // География и природные ресурсы. 2016. № 5. С. 247–254.
Егоров Н.Н., Исаев А.П., Находкин Н.А. Орнитофауна среднего течения р. Алгама // Наземные позвоночные Якутии: экология, распространение, численность. Якутск: ЯФ Изд-во СО РАН. 2002. С. 42–50.
Исаев А.П., Егоров Н.Н., Находкин Н.А. Население птиц в районе промышленного освоения Эльгинского каменноугольного месторождения (ср. течение р. Алгама, Южная Якутия) // Естественнознание и гуманизм. Сб. научных работ. Т. 3. № 2. Томск: Изд-во ТГУ. 2006. С. 42–43.
Коблик Е.А., Редькин Я.А., Архипов В.Ю. Список птиц Российской Федерации. М.: Товарищество научных изданий КМК. 2006. 256 с.
Кузякин А.П. Зоогеография СССР. Ученые записки Моск. Обл. пед. Ин-та им. Н.К. Крупской. 1962. 109 с.
Наумов Р.Л. Птицы в очагах клещевого энцефалита: Автореф. дис. канд. биол. наук. М., 1964. 19 с.
Птицы Советского Союза. М.: Наука, 1951–1954. Т. 1–6.
Равкин Ю.С., Куперштрох В.Л., Трофимов В.А. Птицы лесной зоны Приобья. Новосибирск: Наука. Сиб. отд., 1978. 288 с.
Равкин Ю.С., Ливанов С.Г. Факторная зоогеография: принципы, методы и теоретические представления. Новосибирск: Наука. 2008. 205 с.
Степанян Л.С. Конспект орнитологической фауны России и сопредельных территорий. М.: Академкнига, 2003. 808 с.
Трофимов В.А., Равкин Ю.С. Экспресс-метод оценки связи пространственной неоднородности животного населения и факторов среды // Количественные методы в экологии животных. Л. 1980. С. 135–138.
Харвей Д. Научное объяснение в географии. М.: Изд-во Прогресс, 1974. 502 с.
Цыбулин С.М. Птицы Алтая: пространственно-временная дифференциация, структура и организация населения. Новосибирск: Наука, 2009. 234 с.
Шемякин Е.В., Вартапетов Л.Г., Борисов Б.З., Борисов З.З., Исаев А.П. Летнее население птиц верхних высотно-растительных поясов Алданского нагорья (на примере горы Эвота) // Вестник СВФУ. Т. 11. № 3. 2014. С. 53–59.
Шемякин Е.В., Ларионов А.Г., Егоров Н.Н., Секов А.Н., Исаев А.П. Характеристика летнего населения и вертикальное распределение птиц в горных районах юга Среднетаежной зоны // Научная жизнь. № 2. 2016. С. 220–227.
Штегман Б.К. Основы орнитогеографического деления Палеарктики. Фауна СССР. Птицы. Т. 1. М. Л. 1938. Вып. 2. 156 с.
Gregory R.D. Development of breeding bird monitoring in the United Kingdom and adopting its principles elsewhere // Ring. 2000. V. 22. P. 35–44.
Gregory, R.D. Vorisek P., Strien A.Van, Meyling W.G., Jiguet F., Fornasari L., Reif J., Chylarecki P., Burfield I.J. Population trends of widespread woodland birds in Europe // Biol. Conservation. 2007. V. 149. P. 78–97.
Hengeveld R. Dynamic biogeography. Cambridge, University press, 1990. 249 p.
Jaccard P. Lois de distribution florale dans la zone alpine // Bulletin de la Société vaudoise des sciences naturelles. 1902. V. 38. P. 69–130.
Sanderson F.J., Donald P.F., Paln D.J., Burfield I.J., Bommel F.P. van. Long term population declines in Afro-Palearctic migrant birds // Biol. Conservation. 2006. Vol. 131. P. 93–105.
Svensson S.E. European bird monitoring: geographical scales and sampling strategies // Ring. 2000. V. 22. P. 3–23.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая