

Известия РАН. Серия биологическая, 2023, № 6, стр. 706-715
Экология юрков Fringilla montifringilla L. На севере Западной Сибири
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 марта, 202, Россия
* E-mail: ryzhanovskiy@ya.ru
** E-mail: ryzhanovsky@ipae.uran.ru
Поступила в редакцию 17.03.2022
После доработки 18.05.2023
Принята к публикации 19.05.2023
- EDN: UDDKRG
- DOI: 10.31857/S1026347022100262
Аннотация
Рассмотрены особенности экологии юрков на северном пределе ареала в Приобской лесотундре. Плотность гнездования остается высокой, локально 50–100 пар/км2, вплоть до границы распространения прибрежных лесов и пойменных редколесий вдоль 67-й параллели. Местообитания, пригодные для гнездования юрков, имеются и севернее, но птицы малочисленны. Биотопические условия продвижение птиц на север существенно не ограничивают. Велико значение фенологических условий – летняя часть годового цикла вида должна вписаться в определенный лимит температур и сезонных явлений природы. У юрков нет адаптаций к условиям Субарктики. Это средне- и северо-таежный вид осваивающий лесотундру по мере ее потепления и загущения редколесий.
Юрок – обычная гнездящаяся перелетная птица лесных и лесотундровых ландшафтов Северной Евразии, поэтому многие авифаунистические сводки по регионам России содержат очерки экологии вида. Общим свойством очерков является фрагментарность материала, на основании которого они написаны. Как правило, нет обсуждения адаптивных черт экологии, позволяющих популяциям юрков существовать в той или иной части ареала. Материалы, имеющиеся в моем распоряжении, собраны на северной границе ареала вида, в Приобской лесотундре. Сочетание разных методов сбора: отлов сетями и ловушками с прижизненной обработкой, мечение и повторные отловы; содержание юрков в неволе при разных фотопериодических условиях, учеты на пробных площадках, наблюдения и длительный период исследований позволяют рассмотреть особенности жизни юрков на северном пределе ареала детальнее, чем делалось ранее, в том числе и в нашей (Данилов и др., 1984) сводке. Поскольку ареал юрков заходит в Субарктику, несомненный интерес представляют адаптации к освоению Заполярья, что также является задачей данной работы.
МАТЕРИАЛЫ И МЕТОДЫ
Основной район исследований – долина реки Обь в окрестностях г. Лабытнанги (стационар Октябрьский, 66°40′ с.ш. 66°40′ в.д.) и среднее течение р. Собь на Полярном Урале (ж д. ст. Красный Камень, 66°40′ с.ш. 66°30′ в.д.), где в 1976–1989, 2002–2004, 2016–2018 гг. отыскивали и контролировали гнезда, отлавливали молодых и взрослых юрков, в том числе птиц, ранее окольцованных в гнездовое время. Там же набирали птенцов для выкармливания и дальнейшего содержания при разных фотопериодических условиях. Имеется небольшое количество данных по гнездованию (68 гнездовых карточек), данные по отловам птиц паутинными сетями и большой конусной ловушкой, результаты ежедневных учетов птиц на участке площадью 24 га в 1978–1983 гг. в период занятия территорий, на территории 36 га в 2002–2004 гг.; результаты маршрутных учетов птиц в долине Соби в 1977, 2002–2004 гг., результаты вскрытия птиц, погибших в сетях; материалы экспериментов по передержке взрослых и молодых юрков в неволе при разных фотопериодических условиях. При определении дат начала яйцекладки использовали сведения по гнездам, найденным в период откладывания яиц и по срокам вылупления первого птенца. Продолжительность насиживания определяли по гнездам, найденным с неполной кладкой, а продолжительность выкармливания по датам вылупления первого птенца и ухода из гнезда последнего слетка.
Описание состояния оперения проводили по методике Носкова и Рымкевич (1977), где приведена полная схема расположения птерилий и их отделов. Анализ весьма растянутого процесса замены оперения требует его деления на ряд этапов – стадий (Блюменталь, Дольник, 1966). У юрка, при полной послебрачной линьке выделяются 11 стадий, при частичной постювенальной линьке – 7 стадий. Среднесезонные даты начала, окончания, длительности линьки в природе определяли по уравнениям регрессии (Pimm, 1976) программы Microsoft® Excel 2002.
Часть содержавшихся в летний период юрков (5 первогодков и 6 самцов старше года) осенью перевезли в лабораторию, где содержали птиц до следующего лета. У всех ежедекадно определяли вес, жирность; 4 первогодка содержались в клетках с импульсными счетчиками прыжков и 2 – с регистраторами ночного беспокойства. Помещение лаборатории имело небольшие окна, поэтому в течение 10 ч в лаборатории горели электрические лампы. Когда в октябре длина дня за окном сократилась до 10С : 14Т и продолжала сокращаться дальше, фоторежим в клетках оставался без изменений до середины марта, т.е. до увеличения длины дня свыше 10 ч света.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Пространственное и биотопическое распределение. В Западной Сибири юрок гнездится от южной границы таежной зоны до северной оконечности островных и пойменных лесов зоны лесотундры на 67°–68.5° с.ш. На полуострове Ямал наиболее северные точки гнездования – верховья рек Ядаяходаяха, Хадытаяха и левых притоков Щучьей (Данилов и др., 1984), по склону Полярного Урала вид проникает до верховьев р. Байдарата (68° с.ш.). Здесь юрки населяют высокорослые ивняки и куртины ольхи. При этом они предпочитают те заросли, где растут отдельные деревья, а лесные участки на горных склонах были предпочтительней прибрежных лесов (Головатин, Пасхальный, 2005). Южнее юрки являются обычными обитателями лесов речных долин, где выбирают смешанные негустые древостои из ольхи, березы, ели. Лиственничные леса пользуются меньшим успехом, лиственничных редколесий юрки, видимо, избегают, т.к. в лиственничниках лесотундрового стационара Харп (окрестности г. Лабытнанги) в течение многих лет эпизодически гнездилось не более одной пары (Данилов и др., 1984).
Плотность гнездования зависит от биотопа и в некоторых может быть высокой. Значительное количество сведений по плотности юрков в разных типах местообитаний Полярного Урала приводят Головатин и Пасхальный (2005): от 1.4 ± 0.4 пары/км2 в зарослях высокорослых кустарников до 35.4 ± 4.2 пары/км2 в лесах паркового типа, 41.6 ± ± 5.5 пары/км2 в лесах горных склонов, локально – 52 ± 14.4. В долине р. Собь на участке смешанного леса “137 км” в 1977 г. юрки гнездились с плотностью 51 пары/км2, у станции Красный камень (141 км) в 2002 и 2003 гг. было 16.3–33.3 пары/км2 (Рыжановский, Пасхальный, 2007).
На Южном Ямале, в пойменном лесу р. Хадытаяха, в разные годы (n = 8) юрки гнездились с плотностью 28.6–35.7, в среднем 33.0 ± 1.3 пары/км2 (Рябицев, 1993). В долине среднего течения р. Щучья в 1973 г. плотность гнездования была 10 и 20 пары/км2 (Кучерук и др., 1975). На стационаре Октябрьский в 1978–1983 гг. регистрировали от 36.4 до 81.8 пары/км2, в 2002–2004 гг. 42.1–73.6 пары/км2, средняя плотность за 8-летний период – 62.8 ± 5.7 пары/км2. На границе лесотундры и северной тайги, на территории стационара “Войкар”, средняя за 15 лет наблюдений плотность была 22.6 ± 3.0 пары/км2, локально в пойменном лесу было до 96.8 ± 14.9 пары/км2 (Головатин, Пасхальный, 2005). В северной и средней тайге Обь-Енисейского междуречья плотность гнездования юрков обычно не выше, чем в лесотундре: 46–56 особей/км2 (Вартапетов, 1998), и ниже, чем в северной тайге западного склона Приполярного Урала – 60 ± 5.8 пары/км2 (Шутов, 1990).
Весенняя миграция. На широте Полярного круга прилет всегда начинался в мае, между 3.05–29.05. Средняя дата прилета первых юрков в течение 38 лет – 19.05. Начинается прилет при продолжительности светового дня не менее 17 ч 40 мин (начало мая), но чаще при 20 и более часов света. По сведениям Пасхального (2002), юрок относится к видам, которые в 1986–2002 гг. стали появляться раньше, чем в предшествующие 15 лет (1970–1985 гг.). Сдвиг средней даты прибытия в район г. Лабытнанги составил 5 дней: 21.05 и 16.05 соответственно. Но в последние 5 лет наблюдений (2016–2021 гг.) прилет на территорию сада Арктического стационара УрО РАН начинался между 8.05–29.05, в среднем 18.05 (Штро, личные сообщения). Таким образом, устойчивого смещения начала прилета на более ранние даты в связи с глобальным потеплением нет, тренд смещения с учетом последних дат не достоверен (рис. 1).
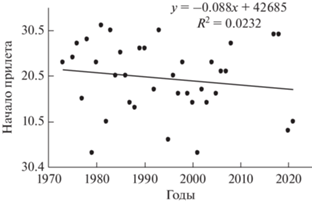
Появление первых юрков наблюдали за 1–3 дня до основного майского потепления, во время которого начинают прилет большинство видов воробьиных лесотундры. Среднесуточная температура воздуха в день встречи первой в сезон птицы колебалась от –3.2° до 10.3° С, в среднем – 2.4°C (n = 34). В период анализируемого временного ряда (1971–2020) наблюдается не достоверное повышение температуры воздуха в первый день прилета юрков).
Период пролета юрков на север по результатам отловов сетями (1977–1983 гг.) и ловушкой (1987–1989 гг.) продолжался 11–36, в среднем 21.6 сут. Поскольку птицы мигрировали лесной полосой, в нижней части которой в 1978–1982 гг. на постоянной позиции стояла линия сетей длиной 50–70 м, было рассчитано общее число юрков, которые должны быть пойманы 100-метровой линией за период прилета. Максимальное их число, 275 юрков, должны быть пойманы в 1978 г., в последующие 1979–1982 гг. должны попасть в сети 73, 50, 65, 67 птиц соответственно. Следует отметить, что весна 1978 г. отличалась поздним, дружным и массовым прилетом многих лесных и кустарниковых воробьиных: в пересчете на 100 м сетей весной поймано 4107 воробьиных птиц против 512–715 птиц в последующие годы. Юрки в 1978 г., возможно, были увлечены на север волной массовой миграции чечеток, на долю которых пришлось 69% численности потока. В последующие годы чечетки были малочисленны, количество пролетающих юрков снизилось и держалось на одном уровне.
Существенных отличий в сроках прилета самцов и самок нет. За 10-летний период отлова на стационаре Октябрьский самцы появлялись на 1–3 сут раньше самок в течение 4 лет, в 1981 г. первой поймана самка, в остальные годы первые самцы и самки пойманы одновременно. Только в 1983 г. в первой и второй декадах с начала прилета соотношение полов было практически равным, в другие годы доминировали самцы (табл. 1). Суммарно, за все годы в две первые декады прилета пойманы 615 самцов и 384 самки. Доля самцов по пятидневкам: 61.2, 63.5, 61.2, 56.6%, доля самцов по годам: 54.5–71.4, в среднем 64.2% (n = 10).
Таблица 1.
Соотношение полов в сетях и ловушке в период прилета в разные годы
| Пол | Год | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1978 | 1979 | 1980 | 1981 | 1982 | 1983 | 1986 | 1987 | 1988 | 1989 | |
| Самец | 90 | 125 | 30 | 35 | 83 | 24 | 127 | 38 | 36 | 38 |
| Самка | 65 | 55 | 15 | 17 | 67 | 21 | 81 | 24 | 20 | 21 |
У самцов юрков по контрасту в окраске больших верхних кроющих второстепенных маховых весной и летом можно отделить первогодков от птиц старше года. Из 304 осмотренных за все годы периода прилета самцов с зарегистрированным возрастом, первогодков было 179 (58.9%). На небольшое преобладание первогодков в период весеннего пролета юрков через Куршскую косу указывает Паевский (2008). Он же приводит сведения о доминировании юрков старших возрастных групп в первые пятидневки миграции. В Нижнем Приобье в первые 10 дней со дня начала прилета из 113 птиц было 57 самцов старше годовалого возраста (51.4%); во вторую декаду из 70 птиц старше года было 28 (40%), что совпадает с данными Паевского (2008).
Среди отловленных весной юрков периодически встречались самки с наседными пятнами, причем регистрировались они уже в первые дни прилета, в конце мая – начале июня, когда самцы только занимали гнездовые участки (Рыжановский, 2008). Это были птицы на второй (фаза яйцекладки) и на четвертой (фаза кормления птенцов) стадиях. Доля таких птиц могла быть значительной. В частности в 1988 г. из 18 самок юрка наседные пятна имели 8 птиц, в 1989 – 6 из 15, в 1981 г. – 4 из 12, в 1983 г. – 2 из 28, в 1986 г. – 2 из 83. В 1978, 1980, 1982 и 1987 таких птиц мы не ловили. Как правило, птицы с наседными пятнами попадали в сети и ловушку в составе стаи, где соотношение полов было почти равным. Например, 10 июня 1981 г. сетью поймана стайка из 8 юрков: 4 самца и 4 самки с пятнами 4-й стадии. В 1989 г. на третий день с начала прилета поймана стайка юрков, включавшая 14 самцов 10 самок, из которых 6 самок также имели наседные пятна 4-й стадии.
Отмечено совпадение сезонов наличия и отсутствия наседных пятен у чечеток (Acanthis flammea) и юрков, что более реально при климатических воздействиях, охватывающих большие площади (Рыжановский, Рябицев, 2021). Можно, также предположить, что птицы на первой-второй стадиях предпринимали попытку гнездования в северной тайге и в южной лесотундре, а на 4-й стадии – в средней тайге. Возможно, у юрков при неудачной попытке гнездования пары не распадаются, но мигрируют на север дальше.
Первая волна прилетающих птиц, судя по упитанности, включает особей с развитым миграционным состоянием. В первую декаду прилета упитанность 21 птицы (8.7%) оценена баллом “много”; среднежирных было 33.4%; остальные отнесены к тощим и маложирным. Во второй декаде доля хорошо упитанных юрков снизилась до 2.7%; доля среднежирных не изменилась (35.2%), а в последующие дни лета очень жирных птиц мы больше не ловили; доля среднежирных птиц в разные декады лета колебалась между 0 и 10% (Рыжановский, 2004).
Занятие территорий и образование пар. Самцы занимали участки, начиная с первой волны прилета. В 1980 г. местное население участка сформировалось за 21 день, в 1981 г. – за 10 дней, в 1982 г. – за 20 дней. Во всех случаях рассматриваются самцы, занявшие участки и отмеченные на них неоднократно. Предполагается, что это одни и те же птицы. В отдельные годы, как показали опыты Рябицева (1993) на северный предел ареала, в пойму Хадытаяхи, юрки прилетали в избытке: после отстрела пары освободившаяся территория занималась повторно, иногда через несколько минут, но в другие годы освободившиеся территории не занимались. Территорию могут занимать пары птиц, сформированные на пролете, (Рябицев, 1993) или при первой попытке гнездования, как предполагалось выше.
Юрок относится к видам со слабой территориальной привязанностью. От 17 гнездившихся в долине Хадытаяхи птиц Рябицев (1993) не получил возвратов. На Приполярный Урал вернулись 5 взрослых юрков из 97 (Шутов, 1989). В Шведской Лапландии вернулся 1 взрослый из 262; из 434 птенцов не вернулся ни один (Lindström, 1987). В долину Соби не вернулись ни один из 149 взрослых и 34 молодых юрков, на стационаре Октябрьский в последующие, после мечения годы, не пойманы ни один из 1175 взрослых, 52 молодых, 35 меченых в гнездах.
Гнездование. Юрки прилетают с достаточно развитыми гонадами, у самцов хорошо развит клоакальный выступ. В первую пятидневку с начала прилета у самцов, отстрелянных или погибших в сетях (n = 7), гонады имели вес 315–429 мг, в среднем 382 ± 16.5 мг; во вторую пятидневку 270–460, в среднем 351 ± 26.0 (n = 8), в четвертую пятидневку семенник одного самца весил 470 мг. Клоакальные выступы максимальных размеров наблюдались у птиц, пойманных из стай в четверную пятидневку (10 из 23), т.е. самцы, прилетающие в хвосте миграционного потока, к гнездованию готовы в максимальной степени. Юрок относится к видам, которые по прилете в Субарктику не нуждаются фотостимуляции полярным днем. Самцы из первой волны прилета, помещенные в условия короткого дня (14С : 10Т), начали линьку в те же сроки, что и самцы, жившие при полярном дне в вольере, т.е. дополнительное пребывание в гнездовом районе для полового созревания этим птицам не нужно (Рыжановский, 2001).
Основной период начала яйцекладки в контрольных гнездах приходился на вторую половину июня и продолжался 8–20 сут, в среднем – 10.6 ± 1.5 (n = 8), а сезон яйцекладки, от первого до последнего яйца, длился 15–25 сут, в среднем 17.6 ± 1.2 (n = 8). В 1978–1982 гг. яйцекладка начиналась через 9–26 сут, в среднем, через 19.1 ± 2.4 сут (n = 6) после устойчивого перехода среднесуточной температуры через 0°, при 4.5–22.8°C, в среднем 10.9 ± ± 1.1°C (n = 6). От занятия самцом гнездового участка до появления в гнезде на участке первого яйца прошло 14 и 19 сут. Из 68 гнезд с полными кладками в 1 было 2 яйца, в 1 – 3 яйца, в 6 по 4 яйца, в 27 – по 5, в 28 – по 6, в 5 – по 7 яиц. Средняя величина кладки – 5.39 ± 0.12. Не достоверно больше кладка юрков в Карелии – 5.59 ± 0.19 (Зимин, 1988).
Насиживает только самка. Судя по растянутости вылупления в некоторых гнездах, устойчивое насиживание может начаться после откладывания второго–третьего яйца при кладке из 6 яиц, чаще – с предпоследнего яйца, но иногда (Рябицев, Шубенкин, 1980) с первого. От первого яйца до вылупления последнего птенца прошло 17–21 сут, в среднем 18.5 ± 0.4 (n = 6), от последнего яйца до первого птенца – 8–14 сут, в среднем 11.0 ± 0.7 (n = 9). Последний период является периодом собственно насиживания, фактически это период насиживания последнего яйца в условиях сформировавшегося ритма. Самцы кормят насиживающих самок, но последние способны высидеть и выкормить птенцов в одиночку. Самка, утратившая самца, насиживала кладку в течение 16 сут от последнего яйца, затем одна выкормила птенцов (Рябицев, Шубенкин, 1980; Рябицев и др.1980). Кладку с погибшими эмбрионами самка насиживала 20 сут, до прекращения наблюдений (Данилов и др., 1984).
Обычные сроки вылупления птенцов: первая–начало второй декады июля. Вылупление чаще растянуто на сутки (n = 9), в одном гнезде оно растянулось на 3 сут, в другом – на 4 сут. Потревоженные птенцы выскакивали из гнезда на 9-й день, не умея летать, не потревоженные покидали гнезда через 10–12 сут, в среднем через 11. 5 ± 0.4 (n = 12) после вылупления. К 12-суточному возрасту, слетки были в состоянии отлететь от гнезда, в зависимости от высоты его расположения, на 10–20 и более метров. Способность к активному полету приобретают к 18–20-суточному возрасту, выводки распадаются по достижению 25–30-суточного возраста слетков.
В гнездах, находившихся под наблюдением от начала яйцекладки до вылета птенцов, общая длительность гнездового периода пары была: 27, 27, 29 сут. От откладки первого, в сезон яйца до вылета последнего в сезон птенца прошло 39–61 сут, в среднем 47.4 ± 4.2 (5 сезонов наблюдений).
Кочевки выводков. Из 35 окольцованных в гнездах слетков юрков повторно, недалеко от гнезда, пойман один в возрасте 24 сут. В начале августа одной сетью были пойманы три птицы в возрасте 20–23 сут, явно из одного выводка. Через 17 сут две из них попались в сети недалеко от места первого отлова. Поскольку у мелких воробьиных к 30-суточному возрасту выводки обычно распадаются и покидают гнездовой район в процессе дисперсионного разлета, можно предположить, что у юрков часть особей в дисперсии не участвует. Но доля таких птиц незначительна, т.к. юрок – единственный вид из певчих нашего района, для которого в течение всех лет отлова было характерно преобладание во второй половине лета взрослых особей (рис. 2). В долине Соби среди отловленных птиц доля взрослых составила 86%, на стационаре Октябрьский – 81.1%. Несомненно, большинство молодых юрков после распадения выводков отлетают из лесотундры. Одновременно возрастает численность юрков в северной тайге Западной Сибири: от 53 особей/км2 в первой половине лета (среднее по ландшафту) до 687 особей/км2 во второй половине (Равкин, 1978). Поскольку местная популяция такой прирост обеспечить не может, есть основание считать, что здесь преобладают юрки более северного происхождения (Рогачева и др., 1983); в енисейской лесотундре также отмечали быструю откочевку выводков в южном направлении.
Рис. 2.
Количество самцов (1), самок (2), молодых (3) юрков, пойманных сетями и ловушками на стационарах Собь и Октябрьский за 5-дневку в конце лета–осенью в 1976–1982 гг.
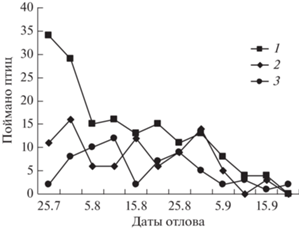
Но небольшая часть молодых юрков остается в зоне лесотундры, а возможно, и в районе вылупления, почти до начала миграции. За 5 лет отлова (1977–1981 гг.) в августе были окольцованы 52 особи, из которых 11 (21.1%) поймано повторно: семь птиц находились в контрольном районе от 1 до 6 сут и 4 птицы – 17–22 сут. Поведение этих юрков (не откочевавших из гнездового района) было типичным для северных воробьиных: сначала кратковременный дисперсионный разлет, затем длительная остановка на постювенальную линьку (Рыжановский, 1997).
Взрослые птицы, судя по повторным отловам, после распадения выводков остаются в районе гнездовой территории до начала отлета. В частности, на стационаре Октябрьский в 1978 г. в августе–сентябре пойманы 10 птиц (7 самцов, 3 самки) из 108 (71 самец, 37 самок), окольцованных на площадке отлова месяцем раньше, в гнездовое время.
Линька. В годовом цикле юрков одна линька, протекающая в гнездовом районе: постювенальная у молодых и послебрачная у взрослых. Постювенальная линька частичная, охватывает контурное оперение, большую часть кроющих крыла, пуховые перья на аптериях, делится на 7 этапов-стадий. У всех пойманных на средних стадиях линьки юрков (n = 20) и выкормленных в неволе при естественном освещении (n = 4) на крыле зарегистрирована линька части (проксимальных) больших, всех средних и малых верхних кроющих второстепенных маховых, верхних и нижних кроющих пропатагиальной складки, кисти, средние нижних кроющих второстепенных маховых; большинство птиц заменяли средние верхние кроющие первостепенных маховых и нижние кроющие третьестепенных маховых (табл. 2). Для юрков Кольского полуострова Гагинская (1990) указывает линьку средних верхних кроющих первостепенных маховых, не обнаруженную нами у птиц, пойманных в природе и у вольерных птиц. Возможно, полнота постювенальной линьки юрков в западной части ареала больше, чем в восточной.
Таблица 2.
Схема линек юрка
| Птерилии, отделы птерилий |
Постювенальная линька стадии |
Послебрачная линька стадии |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Головная | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||
| Брюшная | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||
| Спинная | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||
| Плечевая | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | ||||
| Бедренная | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | ||||
| Голенная | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||
| Анальная | $\square $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||||
| Рулевые | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | ||||||||||
| В.К.Х. | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | ||||||
| Н.К.Х | $\square $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||
| 1-ст. Маховые | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||||
| 2-ст. Маховые | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | ||||||||||||
| 3-ст. Маховые | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | ||||||||||||
| Б.В.К.П.М. | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||||
| С.В.К.П.М. | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||||||||||
| Б.В.К.В.М. | $\square $ | $\blacksquare $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||||||
| С.В.К.В.М. | $\square $ | $\blacksquare $ | $\square $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\square $ | ||||||||
| М.В.К.В.М. | $\square $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | |||||||||
| В.к. проп. | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | ||||||||||
| К. К. | $\square $ | $\blacksquare $ | ||||||||||||||||
| М. Ал. | $\square $ | $\blacksquare $ | $\blacksquare $ | |||||||||||||||
| К. Ал. | $\square $ | $\square $ | $\square $ | $\square $ | $\square $ | $\blacksquare $ | ||||||||||||
| В.К.К. | $\square $ | $\square $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||||||||
| Н.К.К. | $\square $ | $\square $ | $\square $ | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | ||||||||||
| Б.Н.К.П.М. | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||||||||||||
| С.Н.К.П.М. | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||||||||||||
| Б.Н.К.В.М. | $\blacksquare $ | $\blacksquare $ | ||||||||||||||||
| С.Н.К.В.М. | $\square $ | $\blacksquare $ | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | ||||||||||||
| Н.К.Т.М. | $\square $ | $\square $ | $\blacksquare $ | $\blacksquare $ | ||||||||||||||
| Аптерии | $\square $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | $\blacksquare $ | |||||||||||
Обозначения: $\blacksquare $ – перья отдела линяют у всех осмотренных птиц, $\square $ – перья отдела линяют у части осмотренных птиц. Полные названия отделов птерилий: В.К.Х. – верхние кроющие хвоста; Н.К.Х. – нижние кроющие хвоста; Б.В.К.П.М. – большие верхние кроющие первостепенных маховых; С.В.К.П.М. – средние верхние кроющие первостепенных маховых; Б.В.К.В.М. – большие верхние кроющие второстепенных маховых; С.В.К.В.М. – средние верхние кроющие второстепенных маховых; М.В.К.В.М. – малые верхние кроющие второстепенных маховых; В. К. проп. – верхние кроющие пропатагиальной складки; К.К. – карпальное кроющее; М. Ал. – маховые крылышка; К. Ал. – кроющие крылышка; В.К.К. – верхние кроющие кисти; Н.К.К. – нижние кроющие кисти; Б.Н.К.П.М. – большие нижние кроющие первостепенных маховых; С.Н.К.П.М. – средние нижние кроющие первостепенных маховых; Б.Н.К.В.М. – большие нижние кроющие второстепенных маховых; С.Н.К.В.М. – средние нижние кроющие второстепенных маховых; Н.К.Т.М. – нижние кроющие третьестепенных маховых.
Полнота линьки юрков (n = 7), выкормленных и передержанных до начала сентября в лаборатории стационара Октябрьский при фотопериоде 24С : 0Т была не больше, чем в природе. Юрки (n = 7), выкормленные при 16С : 8Т и передержанные при сокращающемся дне (на 30 мин за 5 дней) полноту линьки несколько сократили: заменили только 17–20-е большие верхние кроющие второстепенных маховых против 12–20-го при длинном дне.
В Ленинградской области юрки из ранних выводков начинали линьку в возрасте 30–35 сут, на Кольском полуострове – в 20–25 сут (Гагинская, 1990). Юрки из Нижнего Приобья, выкормленные в разных фотопериодических условиях, начинали ее в разном возрасте (табл. 3). Отличия достоверны между птицами групп короткодневного и длиннодневного фотопериодов, т.е. возраст начала линьки у юрков зависит от длины дня (контролируется фотопериодом). Но возрастной интервал, в пределах которого начинается линька (14 дней), невелик по сравнению, например, с зябликом (Fringilla coelebs) (30 дней) или чечеткой (42 дня) (Носков, 1975, Носков, Смирнов, 1986; Рыжановский, Рябицев, 2021).
Таблица 3.
Возраст начала и длительность постювенальной линьки при разных фотопериодических условиях
| Фотопериод | Возраст начала линьки, сут | Длительность линьки, сут | ||
|---|---|---|---|---|
| N | $\frac{{{\text{Lim}}}}{{M \pm m}}$ | N | $\frac{{{\text{Lim}}}}{{M \pm m}}$ | |
| КФП 16С : 8Т сокращающийся на 6 мин/сутки до 12С:12Т | 7 | $\frac{{20{\kern 1pt} - {\kern 1pt} 26}}{{23.0 \pm 0.9}}$ | 7 | $\frac{{40{\kern 1pt} - {\kern 1pt} 48}}{{43.2 \pm 1.9}}$ |
| ЕФП 24С : 0Т до середины июля сокращающийся позднее | 4 | $\frac{{22{\kern 1pt} - {\kern 1pt} 27}}{{22.6 \pm 1.3}}$ | 4 | $\frac{{44{\kern 1pt} - {\kern 1pt} 60}}{{53.0 \pm 3.9}}$ |
| ДФП 24С : 0Т до конца августа, естественный позднее | 7 | $\frac{{25{\kern 1pt} - {\kern 1pt} 34}}{{28.6 \pm 1.3}}$ | 7 | $\frac{{59{\kern 1pt} - {\kern 1pt} 70}}{{65.8 \pm 1.8}}$ |
В связи со значительной полнотой линьки юрки в неволе линяли достаточно долго, не менее 40 сут при коротком дне, до 70 сут при длинном дне (табл. 3). При фотопериоде 24С : 0Т (до конца августа, естественный позднее) линька длилась достоверно дольше чем при 16С : 8Т (сокращающийся).
В 1977–1982 гг. не начавших линьку молодых птиц отлавливали между 26 июля–22 августа (n = 5), в 2018 г. – 26–28 июля (n = 4). В 1977–1982 гг. мы поймали 20 линяющих юрков между 1 августа–7 сентября на 1–4-й стадиях линьки, одна особь поймана 20 сентября на 5-й стадии. Юрков, закачивающих и тем более закончивших линьку мы не ловили. Из лесотундры молодые юрки отлетают в южном направлении до окончания линьки. Уравнение регрессии (Pimm, 1976), составленное по средним датам линьки, дает следующие результаты: начало – 9 августа, конец – 28 сентября, длительность – 51 сут, что совпадает с ходом линьки клеточных юрков при естественном дне. Сезон постювенальной линьки (сроки встречи молодых линяющих птиц) у юрков Нижнего Приобья продолжается около полутора месяцев, период линьки популяции северных птиц растянут на 2.5–3 мес. и совмещается с периодом миграции.
Послебрачная линька полная. Общая последовательность смены оперения приведена в табл. 2 и не отличается от таковой птиц Ленинградской области (Гагинская, 1990). Часть юрков, возможно значительная, совмещает линьку с размножением. От гнезд с 10–12-дневными птенцами пойманы два самца и три самки. Не приступил к смене оперения один самец, остальные птицы находились на 2-й стадии линьки. Начинающие линьку юрки, на 1–2-й стадиях отлавливались редко. Основная масса пойманных линяющих юрков находилась на 4–7-й и на 10–11-й стадиях. Способность к полету птицы никогда не теряют, но на 8–9-й стадиях, когда растут маховые вершины крыла и большинство второстепенных маховых, летные качества, несомненно, снижаются, поэтому птицы реже ловятся.
Сезон послебрачной линьки начинается в первой декаде июля, в ранневесенние годы, возможно, даже в конце июня (Рымкевич, Рыжановский, 1987) и заканчивается во второй декаде сентября. Юрков в старом оперении отлавливали до 28 июля 1977–4 августа 2018 гг. Первые линяющие птицы пойманы 17–28 июля (n = 4), но все они имели продвинутую линьку (4–6 стадии), т.е. началась она на 5–15 дней раньше. Средняя вычисленная дата (Pimm, 1976) окончания линьки самцов – 7 сентября, самок – 11 сентября, средняя длительность линьки самцов – 63 сут, самок – 58 сут. Судя по этим данным, самки начинают линьку все же несколько позднее самцов, что не выявилось при отловах у гнезд; несколько позднее ее заканчивают, идет она более высокими темпами. Последнее связано с реакцией на сокращающийся день августа. Гагинская (1990) показала, что на Кольском полуострове юрки, начинающие линьку рано, заменяли оперение за 70–75 сут; поздно линяющие – за 55–60 сут.
Период вступления в послебрачную линьку на северном пределе ареала, на широте Полярного круга, растянут почти на месяц. На средних стадиях линьки птицы ловились такое же время. В частности, в 1978 г. на 7-й стадии самцы ловились в течении 25 дней, самки – в течении 31 дня. Самцы, содержавшиеся с весны в вольере при естественном фотопериоде (n = 7), т.е. не участвующие в гнездовании, начали линьку в период с 25 июня по 20 июля, в среднем 10 июля; закончили линьку в первой половине сентября через 60–66 сут, в среднем через 63.4 ± 0.8 сут. В данном случае сроки почти полностью совпали со сроками и темпами линьки самцов, участвующих в размножении.
Осенняя миграция. Отлет начинался в середине августа, продолжался свыше месяца, до третьей декады сентября. Даты последних отловов и встреч юрков на маршрутах на Полярном Урале и в пойме Нижней Оби – 16–21 сентября. Взрослые птицы, которых можно считать “местными”, (окольцованы в июне–июле) перестали отлавливаться повторно, т.е. включились в миграцию, между 16 августа–17 сентября. В этот же период на участке возрастало число не меченных юрков, заканчивающих линьку и в новом наряде. Предположительно, они гнездились севернее нашего района, т.е. являлись пролетными. Возможно, первые этапы миграции совершаются бросками с продолжительными остановками. В частности, в 1978 г. на участке в период с 29 августа по 2 сентября окольцованы 8 птиц на 10–11-й стадиях линьки, все они пойманы повторно через 2–16 дней, некоторые в новом оперении. Связанных с полом различий в сроках отлета, вероятно, нет, самки ловились до конца миграции. Молодые юрки, как уже говорилось выше, из лесотундры в массе уходят в первой половине августа. Оставшиеся отлетают одновременно с взрослыми птицами. В отличие от весенней миграции, когда юрки летят стаями, одновидовыми или совместно с чечетками, осенью стаи юрков очень редки, преобладают небольшие группы.
Осеннее миграционное состояние размножавшихся птиц начинает формироваться поздно. Для определения сроков появления южного направления активности в конусные клетки (Emlen, Emlen, 1966) помещали взрослых птиц, пойманных в третьей декаде августа (n = 12) и в первой декаде сентября (n = 10). В августе отмечено небольшое преобладание беспокойства в восточном и западном направлениях, в сентябре юрки начинают выбирать южное направление активности (Рыжановский, 2010). Жировые накопления у взрослых птиц также образуются весьма поздно. Из 28 юрков, пойманных в третьей декаде августа запасы жира, оцениваемые как “средние”, имела одна особь; в первой декаде сентября все 24 птицы были тощими или маложирными, во второй декаде сентября средние запасы жира имели 3 птицы (16.6%) из 18 осмотренных.
У молодых юрков, вероятно, существует два периода проявления южного направления активности: 1 – кратковременный у большинства птиц, на начальных этапах послегнездовых кочевок, во время которых основная часть лесотундровых юрков переселяется в тайгу, 2 – длительный у всех, с началом формирования миграционного состояния. Интересно также, что во второй декаде августа, в период откочевки в тайгу, среди 12 пойманных юрков 3 имели средние запасы жира, т.е. сформировался еще один компонент миграционного состояния, но, вероятно, также кратковременный, до развития интенсивной линьки.
Юрки из нашего района летят на юг Западной Европы, о чем свидетельствуют данные кольцевания. В 1979 г. в середине июля на стационаре Октябрьский был пойман самец с югославским кольцом. Сведения о конкретном месте мечения не получены. Самка юрка, окольцованная 12.8.1980 на стационаре Октябрьский поймана 5.11.1980 в Италии: 45°45′ N, 12°35′ E. Поскольку раньше начала сентября из нашего района она отлететь не должна, т.к. в день мечения находилась на 4-й стадии линьки, на весь миграционный путь птица потратила менее 2-х мес.
ЗАКЛЮЧЕНИЕ
В Западной Сибири северная граница ареала юрка совпадает с северной границей пойменных лесов; лиственничных, ивовых, ольховых лесков в котловинах озер и горных долинах. Плотность гнездования остается высокой, локально 50–100 пар/км2, вплоть до границы почти сплошного распространения прибрежных лесов и участков пойменных редколесий вдоль 67-й параллели. Местообитания, пригодные для гнездования юрков, имеются и севернее, например лесной остров значительных размеров в долине Ядаяходаяхи, но в 80-х годах эти птицы там гнездились нерегулярно и с низкой плотностью. Вероятно, непосредственно биотопические условия продвижение вида на север не ограничивают.
Данилов (1966) при анализе факторов, определяющих северные границы распространения птиц в Субарктике, пришел к выводу о решающем значении фенологических условий, т.е. летняя часть годового цикла вида должна вписаться в определенный лимит температур и сезонных явлений природы. Юрки прилетают в северную часть ареала при дневных положительных температурах (близких к 0°C), с началом активного таяния снега в лесу, и отлетают с началом фенологической осени при положительных, иногда весьма высоких, температурах. Отлет последних особей заканчивается после опадения листвы, но до первого снегопада. Общая продолжительность пребывания юрков в Нижнем Приобье, от регистрации первой весенней птицы до регистрации последней птицы в 80-е годы: 102–137 сут, в среднем 112.2 (n = 4) сут, а средняя продолжительность безморозного периода на широте г. Салехарда – 94 сут. Юрки при сопоставлении средних дат начала массового прилета (третья декада мая) и завершения массового отлета (первая декада сентября) в целом вписываются в безморозный период лесотундры. С учетом длительности безморозного периода на широте пос. Тамбей (Северный Ямал, 71° с.ш.) – 51 сут (Орлова, 1962), его сокращение составляет 7.8 сут/1° географической широты и в долине р. Ядаяходаяхи этот период длится около 85 сут. Отличия от широты Полярного круга кажутся незначительными, но в безморозный период вид уже не вписывается. Потепление последних десятилетий должно привести к более раннему началу прилета юрков в лесотундру, что и наблюдается в некоторые годы, но средние за 5 лет даты не изменились.
С другой стороны, прилетая в лесотундру с созревшими гонадами, птицы утрачивают стимулы к миграции и не должны стремиться расширить ареал за счет локальных редколесий в тундровой зоне. Фотопериодические условия южной тундры (полярный день на протяжении 1.5–2-х мес.) дальнейшее продвижение на север не стимулируют. Для начала гнездостроения и откладывания яиц юркам достаточно светового режима таежной зоны.
По моему мнению, у юрков нет адаптаций к условиям Субарктики. Это средне- и северо-таежный вид осваивающий лесотундру по мере ее потепления и загущения редколесий.
Список литературы
Блюменталь Т.И., Дольник В.Р. Географические и внутрипопуляционные различия в сроках размножения, линьки и миграции у некоторых перелетных воробьиных птиц // Тр. Всесоюз. совещ. по внутривидовой изменчивости наземных позвоночных и микроэволюции. Свердловск: 1966. С. 319–332.
Вартапетов Л.Г. Птицы северной тайги Западно-Сибирской равнины. Новосибирск: Наука, 1998. 327 с.
Гагинская А.Р. Юрок – Fringilla montifringilla L. // Линька воробьиных птиц Северо-Запада СССР. Л.: ЛГУ, 1990. С. 208–212.
Головатин М.Г., Пасхальный С.П. Птицы Полярного Урала. Екатеринбург: Изд-во Уральск. ун-та, 2005. 564 с.
Данилов Н.Н. Пути приспособлений наземных позвоночных животных к условиям существования в Субарктике. Т. 2. Птицы. Свердловск: 1966. 140 с.
Данилов Н.Н., Рыжановский В.Н., Рябицев В.К. Птицы Ямала. М.: Наука, 1984. 332 с.
Зимин В.Б. Экология воробьиных птиц Северо-Запада СССР. Л.: Наука, 1988. 184 с.
Кучерук В.В., Ковалевский Ю.В., Сурбанос А.Г. Изменения фауны птиц Южного Ямала за последние 100 лет // Бюл. Моск. о-ва испытателей природы. Отд. биологии. 1975. Т. 80. № 1. С. 52–64.
Носков Г.А. Линька зяблика (Fringilla coelebs) // Зоол. журн. 1975. Т. 54. № 3. С. 413–425.
Носков Г.А., Рымкевич Т.А. Методика изучения внутривидовой изменчивости у птиц // Методика исследования продуктивности и структуры видов в пределах их ареалов. Вильнюс: Моклас, 1977. Т. 1. С. 37–48.
Носков Г.А., Смирнов Е.Н. Некоторые особенности фотопериодического контроля сроков и полноты постювенальной линьки обыкновенной чечетки // Актуальные проблемы орнитологии. М.: 1986. С. 69–75.
Орлова В.В. Климат СССР. Вып. 4. Западная Сибирь // Л., Гидрометеоиздат, 1962. 359 с.
Паевский В.А. Демографическая структура и популяционная динамика певчих птиц. СПб.: М.: КМК, 2008. 235 с.
Пасхальный С.П. Сроки прилета некоторых видов птиц в низовья Оби в 1970–2002 гг. // Многолетняя динамика численности птиц и млекопитающих в связи с глобальным изменением климата. Материалы международного симпозиума. Казань: “Новое знание” 2002. С. 151–156.
Равкин Ю.С. Птицы лесной зоны Приобья. Новосибирск: Наука, 1978. 287 с.
Рогачева Э.В., Равкин Е.С., Сыроечковский Е.Е., Кузнецов Е.А. Фауна и население птиц енисейской лесотундры // Животный мир енисейской лесотундры и природная зональность. М.: Наука, 1983. С. 14–47.
Рыжановский В.Н. Экология послегнездового периода жизни воробьиных птиц Субарктики. Екатеринбург: Изд-во Урал ун-та, 1997. 288 с.
Рыжановский В.Н. Гнездовой сезон как часть годового цикла воробьиных Субарктики // Гнездовая жизнь птиц. Пермь: Пермский гос. пед. ин-т, 2001. С. 3–22.
Рыжановский В.Н. Масса тела и жировые резервы воробьиных птиц Нижнего Приобья // Рус. орнитол. журн. 2004. № 13. (271). С. 799–812.
Рыжановский В.Н. О попытках гнездования некоторых видов вьюрковых на путях весенней миграции // Рус. орнитол. журн. 2008. № 17 (418). С. 731–733.
Рыжановский В.Н. Формирование направления осенней миграции у некоторых воробьиных Нижнего Приобья // Рус. орнитол. журн. 2010. № 19 (564). С. 671–680.
Рыжановский В.Н., Пасхальный С.П. Динамика населения птиц Нижнего Приобья в связи с потеплением климата // Научный вестник Ямало-Ненецкого автономного округа. Салехард: 2007. № 6 (50). Ч. 2. С. 58–74.
Рыжановский В.Н., Рябицев В.К. Биология и экология чечетки (Acanthis flammea sensu lato, Passeriformes, Fringillidae) на Ямале и в Приобской лесотундре // Зоол. журн. 2021. Т. 100. № 2. С 203–219.
Рымкевич Т.А., Рыжановский В.Н. Линька птиц на Полярном Урале // Орнитология. М.: Изд-во Моск. ун-та, 1987. Т. 22. С. 84–95.
Рябицев В.К. Территориальные отношения и динамика сообществ птиц в Субарктике. Екатеринбург: Наука, 1993. 296 с.
Рябицев В.К., Шубенкин В.П. О длительности насиживания у юрка // Инф. мат-лы ИЭРиЖ. Свердловск. 1980. С. 80–81.
Рябицев В.К., Головатин М.Г., Якимкенко В.В. Территориальность воробьиных в условиях весеннего половодья и экспериментального изъятия самцов // Экологические аспекты поведения животных. Свердловск. 1980. С. 49–60.
Шутов С.В. Территориальный консерватизм, дисперсия массовых видов воробьиных на Приполярном Урале и некоторые закономерности их широтных изменений // Экология. 1989. № 4. С. 69–74.
Шутов С.В. Плотность гнездования массовых видов воробьиных (Passeriformes) на Приполярном Урале и некоторые закономерности ее пространственного изменения в поймах Южной Субарктики // Зоол. журн. 1990. Т. 69. № 5. С. 93–98.
Emlen S.T., Emlen J.T. A technique for recording migratory orientation of captive birds // Auk. 1966. V. 83. P. 361–367.
Lindström A. Breeding nomadism and site tenacity in the bramling, Fringilla montifringilla // Ornis. Fenn. 1987. V. 64. № 2. P. 50–56.
Pimm S.L. Estimation of the duration of bird moult // Condor. 1976. V. 78. № 4. P. 550.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая