

Известия РАН. Серия физическая, 2022, T. 86, № 10, стр. 1458-1472
От нано- к макромеханическим свойствам древесины через иерархию структуры и размерные эффекты (обзор)
Ю. И. Головин 1, 2, *, А. А. Гусев 1, 3, С. М. Матвеев 3
1 Федеральное государственное бюджетное образовательное учреждение высшего образования
“Тамбовский государственный университет имени Г.Р. Державина”
Тамбов, Россия
2 Федеральное государственное бюджетное образовательное учреждение высшего образования
“Московский государственный университет имени М.В. Ломоносова”
Москва, Россия
3 Федеральное государственное бюджетное образовательное учреждение высшего образования
“Воронежский государственный лесотехнический университет имени Г.Ф. Морозова”
Воронеж, Россия
* E-mail: yugolovin@yandex.ru
Поступила в редакцию 01.06.2022
После доработки 15.06.2022
Принята к публикации 22.06.2022
- EDN: UZZOKC
- DOI: 10.31857/S0367676522100076
Аннотация
Описано состояние исследований иерархической структуры древесины – от нано- до макромасштабного уровня современными физическими методами. Рассмотрена связь структуры с механическими свойствами на всех этих уровнях. Показано, что размерные эффекты в прочности древесины следуют соотношению Холла–Петча в интервале около 6 порядков величины характерных размеров структуры.
ВВЕДЕНИЕ
К началу 21-го в. стало совершенно очевидным, что непрерывно растущие объемы добычи минеральных ресурсов (особенно, различных руд и нерудных материалов, угля, нефти и газа) и незамкнутые технологии их переработки приводят к росту объемов промышленных и бытовых отходов. Это создает угрозу биосфере вследствие загрязнения окружающей среды и роста концентрации диоксида углерода в атмосфере. Малая доля материалов, подвергающаяся вторичному использованию, усугубляет положение. К этому добавляется опережающий рост производства упаковочных полимерных материалов, львиная доля которых не биоразлагаема, а значительная часть не участвует в рециклинге. Нарастающее давление на окружающую среду требует всё больших сил и средств для его нейтрализации.
В этой связи одной из перспективных мер представляется замена традиционных материалов целлюлозосодержащими, в первую очередь модифицированной древесиной, растительными отходами сельского хозяйства и в особенности – содержащейся в них наноцеллюлозой. Целлюлоза – наиболее распространенный в биосфере природный линейный полимер – полисахарид (C6H10O5)n. Материалы на его основе обладают многими достоинствами:
1) в отличие от абсолютного большинства добываемых полезных ископаемых целлюлозосодержащие материалы имеют возобновляемые, стабильные источники сырья – леса, полевые культуры, аква ресурсы;
2) эти материалы многофункциональны и могут применяться в строительстве, промышленном производстве [1–6], медицине [7–9] и быту [1, 2];
3) они технологичны и не токсичны;
4) одной из привлекательных особенностей биокомпозитов, роль которых с каждым годом нарастает, является их биоразлагаемость, “самоуничтожение” после окончания срока службы без образования токсичных продуктов [1–6];
5) с их помощью легко организовать замкнутый углеродный цикл, не повышающий содержания углекислого газа в атмосфере, который создает парниковый эффект и рост среднегодовой температуры на Земле;
6) нано- и микроструктурные компоненты в структуре древесины (нанокристаллы, нанофибриллы, микроволокна целлюлозы) обладают механическими характеристиками, сопоставимыми или превышающими таковые у высокопрочных конструкционных материалов (сталей, титановых и алюминиевых сплавов). Удельная прочность нано-/микро- целлюлозы может на порядок и более превышать таковую в сталях (с учетом пятикратно более низкой плотности ρ ~ 1.5 г/см3 против ~8 г/см3 у стали).
Разумеется, древесине и другим целлюлозосодержащим материалам присущ и ряд недостатков. Без дополнительной обработки они горючи, гигроскопичны, при увеличении влажности теряют часть прочностных свойств, подвержены гниению и биоповреждениям. Кроме того, древесина любых пород обладает существенной зависимостью механических свойств от условий роста, эксплуатации и испытаний: влажности, структуры клеточных стенок и годовых колец роста, доли ранней и поздней древесины, напряженного состояния, размеров образца или нагруженной области, направления, скорости и продолжительности приложения нагрузки. Это затрудняет выявление наиболее общих закономерностей формирования механических свойств древесины различных пород. Тем не менее, литературные данные и накопленный опыт позволяют сделать некоторые обобщения, изложенные ниже.
В обзоре рассмотрены методы и результаты мультимасштабного изучения структуры и механических свойств древесины различных пород с учетом нано-, микро- и мезомасштабного структурного уровня ее строения. В заключение проведенного анализа литературных данных показано, что в громадном диапазоне характерных размеров структуры (около 6 порядков величины) механические свойства древесины в целом следуют соотношению Холла–Петча, хорошо известному специалистам-металловедам.
ИЕРАРХИЧЕСКАЯ СТРУКТУРА ДРЕВЕСИНЫ
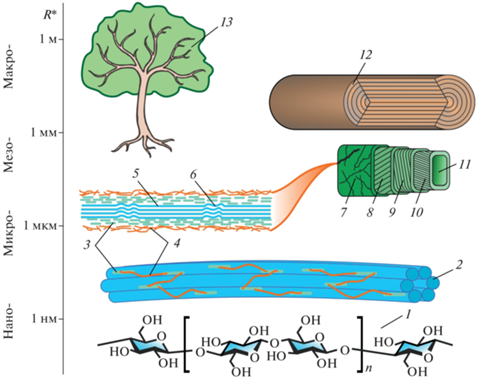
С точки зрения физического материаловедения древесина – это иерархически сложно структурированный природный композитный материал с ярко выраженной гетерогенностью и анизотропией всех свойств, а также способностью к регенерации [10–13]. В структуре древесины можно выделить (весьма условно) несколько масштабно-иерархических уровней (рис. 1) – атомно-молекулярный, нано- (нанокристаллы, нанофибриллы), микро- (микроволокна, клеточные стенки) мезо- (клетки, крупные сосуды) и макро- (годовые кольца, макродефекты строения, трещины и т.п.). Каждый из них вносит свой вклад в формирование всего комплекса физико-химических и механических свойств. Большой диапазон характерных размеров структурных компонентов древесины (6–8 порядков величины) и широкий спектр задач и вопросов, возникающих при ее изучении, требует большого арсенала исследовательски методов и средств для их реализации. Кратко они будут проанализированы в следующем разделе.
Рис. 1.
Масштабно-иерархические уровни структуры древесины. 1 – Полимерная молекула целлюлозы, 2 – элементарные нанофибриллы, 3 – гемицеллюлоза, 4 – лигнин, 5 – нанокристаллическая зона, 6 – аморфная зона, 7 – первичная стенка клетки, 8, 9 и 10 – слои S1, S2 и S3 вторичной стенки клетки, 11 – трахеида (капилляр), 12 – годовые кольца, 13 – дерево.
Важнейший аспект древесиноведения – закономерности формирования макросвойств древесины в функции нано-, микро-, мезо- и макроструктуры. Существует множество причин интереса к связям макромеханических свойств древесины с ее нано- и микроструктурой. Назовем наиболее важные из них. Во-первых, соответствующие закономерности проясняют природу и механизмы формирования наиболее значимых для практики физико-механических свойств древесины в макрошкале. Во-вторых, наномеханические прочностные характеристики, будучи значительно более высокими, чем таковые на микро- и макроуровне, указывают на потенциальные возможности упрочнения, близкие к теоретическому пределу прочности нанокристаллической целлюлозы (~10 ГПа). В-третьих, расширяющееся применение композитных материалов в различных областях техники, строительства, медицины позволяет вытеснять традиционные металлы и сплавы более легкими и более экологичными композитами. Так, фюзеляжи последних моделей Боингов и Эйрбасов уже более, чем на половину по массе состоят из армированных композитов. Они приобретают все большую популярность и в других областях – автомобилестрении, судострении, производстве спортивного инвентаря и др. Однако применяемые для армирования стекло или базальтовое волокно (не говоря о микро- и нано- углеродном) весьма дороги и пока не могут завоевать широкий рынок потребительских товаров. Целлюлозное волокно примерно на порядок дешевле стекловолокна и не сильно уступает ему по механическим характеристикам. Поэтому задачи углубления понимания природы прочности и механизмов разрушения, поиска методов упрочнения микроцеллюлозного волокна, тканей и нетканых материалов из него весьма актуальны. Наконец, в-четвертых, многие породы деревьев, имея период вегетации в течение нескольких сотен, а иногда и тысяч лет (например, секвойя), накапливают в своей нано-, микро-, мезо- и макроструктуре большой массив информации о климатических условиях роста и экологических катастрофах, Этот природный архив может служить источником информации для климатологии и датировки различных событий в истории Земли (дендрохронология).
ФИЗИЧЕСКИЕ МЕТОДЫ ИССЛЕДОВАНИЯ СТРУКТУРЫ И СВОЙСТВ ДРЕВЕСИНЫ НА РАЗЛИЧНЫХ ИЕРАРХИЧЕСКИХ УРОВНЯХ
Классическое материаловедение древесины фокусируется на выявлении и описании зависимости макромеханических, физико-химических и служебных свойств от внутренних структурных характеристик, а также внешних термодинамических факторов [14–21]. Начиная с конца прошлого века, все больше внимания стало уделяться тонкой структуре древесины на наноуровне. Этому способствовало как развитие нанотехнологии и нанометрологии, так и осознание того, что неиспользованные резервы кроются именно на наноуровне.
В последние 1.5–2 десятилетия к исследованиям микроструктуры и свойств древесины стали привлекать все в большем объеме современные методы и средства, традиционные в физике твердого тела и физическом материаловедении [2, 3, 22–24]. Микроструктуру изучают с помощью просвечивающей и сканирующей электронной микроскопии, сканирующей зондовой (преимущественно атомно-силовой), конфокальной лазерной и оптической микроскопии в различных модах [2, 3, 25–27]. Для определения состава, атомной и микроструктуры используют большой набор рентгеновских методов. Характер и степень упорядоченности молекул целлюлозы в нановолокнах, угол между микроволокнами и длинной осью клеток определяют методом рентгеновской дифрактометрии [2, 3, 28–30] и микротомографии [31, 32], синхротронного рентгеновского излучения [33], малоуглового (SAXS) и широкоуглового (WAXS) рентгеновского рассеяния [2, 3]. Элементный и молекулярный состав выявляют различными спектроскопическими методами. Используют рентгеновскую флуоресценцию [34], различные виды спектроскопии – инфракрасную (ИК) и ИК с фурье-преобразованием (FTIR) [35‒39], рамановскую [39–42], бриллюэновскую [43], ядерную магниторезонансную (ЯМР) [44] и другие аналитические методы. Совместно они перекрывают громадный пространственно-временной диапазон структур и событий в них (рис. 2а) [45]. Сравнительный анализ возможностей наиболее распространенных физических методов исследования молекулярной, субклеточной и клеточной структуры древесины можно найти в недавних обзорах [14, 45].
Рис. 2.
Карта диапазонов пространственно-временного разрешения наиболее популярных физических (а) [45] и нано-/микромеханических методов исследования структуры древесины (б). 1 – Спектроскопия (ЯМР, ИК, FTIR, Raman), 2 – рентгеновская дифрактометрия (XRD), 3 – рассеяние рентгеновских лучей (SAXS, WAXS), 4 – Атомно-силовая микроскопия (АСМ), 5 – рентгеновская компьютерная томография (ХСТ), 6 – электронная микроскопия (TEM, SEM), 7 – конфокальная лазерная микроскопия, 8 – оптическая микроскопия, 9 – оптический пинцет (OT), 10 – магнитный пинцет (MT), 11 – наноиндентирование (НИ), 12 – тестирование механических свойств на миниатюрных образцах SSMT).

Механические свойства в нано- и микрошкале исследуют методами нано-/микро-механического тестинга (SSMT – Small Scale Mechanical Testing) [46–49], чаще всего с помощью атомно-силовых микроскопов (АСМ) [50–53] и наноиндентометров (НИ) [54–62] (Рис. 2б). Их структурная схема и возможности схожи [55–57]. В обоих случаях зонд с радиусом закругления от единиц (в АСМ) до десятков (в НИ) нанометров подводится к тестируемой поверхности прецизионными приводами и начинает взаимодействовать с ней. При этом непрерывно измеряют силу Р и глубину внедрения h зонда и регистрируют кинетику их изменения в течение всего цикла испытания. Затем на основании этих данных строят P–h диаграмму (аналог диаграммы σ–ε при макроиспытаниях) и с помощью стандартизованных алгоритмов извлекают около десятка различных механических характеристик материала (модуль Юнга, твердость, вязкость разрушения, скорость ползучести и др.) в нано-/микро- масштабной шкале. В НИ используют трехгранную алмазную пирамиду Берковича, гораздо лучше аттестованную с точки зрения реальной геометрии вершины, чем в АСМ, что обеспечивает более точные и достоверные данные.
Методами SSMT исследовали механические свойства отдельных нанофибрилл и микроволокон целлюлозы [63–67], клеточных стенок [45, 68–74], годовых колец древесины различных пород и получили много интересных данных. Однако они относились к определенным локациям в структуре и не предполагали сканирования или картирования механических свойств области, охватывающей несколько годовых колец роста. Это не позволяет проследить связь свойств отдельных нано- и микроструктурных элементов древесины с ее макромеханическими свойствами.
В противоположность микроструктурным и физико-химическим исследованиям древесины, основным инструментом изучения и анализа годовых колец роста в интересах дендрохронологии, дендроклиматологии, прогнозирования прочности и других служебных свойств являются простые оптические методы, в которых первичная информация основана на различии отражательных способностей ранней и поздней древесины (EW и LW соответственно) [75]. С помощью этих методов извлекают чисто геометрические и морфологические характеристики объекта изучения (толщину годовых колец роста, долю EW и LW в них, их вариации от кольца к кольцу и т.д.). В [76] сравнивают и анализируют данные, полученные разными методами. Предпринимались многочисленные попытки усовершенствования традиционных методов дендрохронологии, главным образом, путем модификации методов пробоподготовки, использования не белого, а синего света, применения технологий технического зрения и математической обработки данных (см., например, [27, 77–85]). Однако, несмотря на это, возможности подхода, в основе которого лежит фоторегистрация и анализ изображения поперечного среза, остаются сильно ограниченными, поскольку отражательные оптические свойства древесины весьма изменчивы и неоднозначно или очень слабо связаны с другими ее физическими характеристиками, в частности, с механическими.
В [31–33, 86, 87] описаны попытки применения современных физических методов и в дендрохронологии, где необходимо получение информации не в одной точке, а как минимум, вдоль радиальной координаты. Применяли также двумерное картирование свойств на поперечном срезе ствола дерева, 3D рентгеновскую [31, 32] и ЯМР томографию [43]. Их краткое описание и сравнение дано в обзоре [58]. Однако эти методы сложны в реализации, трудоемки и требуют дорогостоящего или уникального оборудования, поэтому они применяются эпизодически. Более простым является метод рентгеновской денситометрии [88], но он требует двустороннего доступа к плоскому образцу, вырезанному строго перпендикулярно длинным осям клеток.
Отметим, что механические свойства целлюлозосодержащих материалов обладают существенной зависимостью от скорости монотонного нагружения, частоты осциллирующей нагрузки, длительности приложения сил. Они могут варьироваться от образца к образцу и меняться в процессе эксплуатации в гораздо большем диапазоне, чем в близких по архитектуре техногенных композитах (например, в армированных стекло- и углепластиках) [6, 7, 89]. Такая вариабельность свойств еще больше осложняет выявление общих закономерностей их формирования.
К настоящему времени сквозные связи между свойствами на всех масштабно-иерархических уровнях – от нанокристаллов целлюлозы до макрообразцов – изучены недостаточно полно. Однако информации на каждом из этих уровней в отдельности достаточно много. Ниже она приведена в порядке нарастания характерного масштаба структуры.
НАНОЦЕЛЛЮЛОЗА И ЭЛЕМЕНТАРНЫЕ НАНОФИБРИЛЛЫ
Основной “строительный” материал древесины, стеблей злаковых, тростниковых растений, бамбука и т.п., который несет силовую нагрузку, – это целлюлоза – наиболее распространенный природный полимер. Целлюлоза – это высокомолекулярный полисахарид (C6H10O5)n, в котором молекулы глюкозы связаны β-(1 → 4) связями в линейные неразветвленные цепи длиной от нескольких сотен до многих тысяч n [23] (рис. 1). По происхождению целлюлоза может быть двух видов – растительная или бактериальная [7, 9, 23]. Молекулы целлюлозы легко образуют нанокристаллы с поперечником 3–10 нм и типичной длиной 100–300 нм (рис. 3а), из которых формируются нанофибриллы диаметром 5–20 нм и длиной до многих сотен и тысяч нанометров (рис. 3б). Целлюлоза в нанофибрилле находится в аморфно-кристаллическом состоянии в виде чередующихся доменов. Присутствие аморфной фазы несколько ослабляет нанофибриллу, но делает ее более гибкой и пластичной. Типичные морфологические характеристики наноцеллюлозных образований приведены в табл. 1.

Таблица 1.
Структурные характеристики нанокристаллов и элементарных нанофибрилл целлюлозы [67]
| Характеристики | Нанокристаллы | Нановолокна |
|---|---|---|
| Длина наночастиц, нм | 100–500 | ≥103 |
| Поперечный размер наночастиц, нм | 5–30 | 10–40 |
| Аспектное отношение наночстиц | 10–50 | 60–100 |
| Длина нанокристаллитов, нм | 70–200 | 60–150 |
| Поперечный размер нанокристаллитов, нм | 5–10 | 3–7 |
| Степень кристалличности, % | 72–80 | 50–65 |
| Доля аморфной фазы, % | 20–28 | 35–50 |
| Плотность, г · см–3 | 1.57–1.59 | 1.54–1.56 |
| Пористость, см3 · г–1 | 0.01–0.05 | 0.1–0.2 |
Механические свойства нанокристаллической целлюлозы (табл. 2) определяли различными способами, включая расчеты прочности связей в макромолекуле и между ними, компьютерное моделирование, обработку данных ИК и рамановской спектроскопии, АСМ, WAXS и др. Разброс данных обусловлен как особенностями расчетных схем, моделей и алгоритмов обработки первичных данных, так и трудностями проведения прямых измерений механических свойств в наношкале. Во втором случае также сказываются различия в возрасте, структуре и происхождении древесины.
Таблица 2.
Механические характеристики нанокристаллов и элементарных нанофибрилл целлюлозы [67]
| Характеристики | Нанокристаллы | Нановолокна |
|---|---|---|
| Модуль Юнга продольный, ГПа | 140–160 | 30–40 |
| Модуль Юнга поперечный, ГПа | 15–30 | 10–15 |
| Прочность на растяжение вдоль волокон, ГПа | 8–10 | 0.8–1 |
| Прочность на растяжение поперек волокон, ГПа | Около 1 | Около 0.1 |
НАНО- И МИКРОВОЛОКНА ЦЕЛЛЮЛОЗЫ
Типичная иерархия структуры древесины более высокого уровня включает нано- и микроволокна. Они формируются из элементарных нанофибрилл вследствие их взаимодействия (главным образом, через водородные связи). Нанофибриллы образуют пучки, погруженные в матрицу, которая состоит преимущественно из лигнина (полимер ароматической природы – полифенол), гемицеллюлозы (низкомолекулярный разветвленный полисахарид), пектина (желеобразующий полисахарид) и воды [23]. Содержание целлюлозы в волокнах варьируется в разных породах древесины от 40 до 60% (в хлопковых волокнах может превышать 96%) [7, 9, 23].
Нано-/микроструктура целлюлозных материалов, как и их свойства, сильно зависят от характера взаимодействия нанокристаллов и элементарных нанофибрил, а также степени их упорядоченности в нано- и микроволокнах. Механические, в частности, прочностные характеристики целлюлозных нано-/микроструктур являются структурно-чувствительными, как и в большинстве других органических и неорганических материалов. В свою очередь, морфология и внутренняя структура определяются видовым происхождением сырья, технологией выделения целлюлозы и степенью упорядоченности структуры [23, 90–93]. Решающую роль в формировании свойств волокон играет степень кристалличности и доля целлюлозы в нановолокне, характер их связанности в материале. Большую роль в формировании механических свойств играет угол μ между осью нанофибрилл и осью волокна или клетки.
В обзоре [90] приводятся разнообразные сведения о морфологии, микроструктуре, механических свойствах наноцеллюлозных волокон различного происхождения (рис. 4а), а также примеры их использования в качестве армирующих компонентов для упрочнения полимерных композитов. Наиболее прочные из них имеют модуль Юнга Е = 75–85 ГПа и прочность на растяжение σb = 1.6–1.7 ГПа (отношение Е/σb ≈ 50) [66]. Такие высокие механические характеристики достигались за счет упорядочения нанофибрилл и их поперечной сшивки (рис. 4б). Согласно фундаментальным представлениям, теоретическая прочность любого бездефектного материала должна составлять около 0.1Е, в то время как самые прочные микроцеллюлозные волокна, из описанных выше, демонстрировали прочность на уровне 0.015–0.020Е. Отсюда следует, что даже самые прочные из испытанных микроволокон целлюлозы имеют потенциал, как минимум, 3–5 кратного увеличения прочности.
Рис. 4.
Прочность σb в функции модуля Юнга Е (a) различных природных целлюлозосодержащих микроволокон [90] и диаграммы нагружения σ–ε (б) высокоориентированных целлюлозных микроволокон в несшитом (1) и сшитом поперечными связями (2) состоянии [66]. (а) 1 – Капок (хлопковое дерево), 2 – хлопок, 3 – кенаф (гибискус коноплевый), 4 – конопля (пенька), 5 – древесина, 6 – Рами (китайская крапива), 7 – бамбук, 8 – лен, 9 – ковыль тянущийся. (б) 1 – до и 2 – после сшивания целлюлозных нанофибрилл молекулами лигнина. На врезках показаны структурные молекулярные схемы: 3 – целлюлозы, 4 – лигнина, 5 – поперечной сшивки.

Следует также отметить, что данные по механическим свойствам целлюлозных микроволокон, полученные разными методами, существенно различаются (табл. 3) [90]. Результаты, полученные с помощью АСМ и НИ, согласуются между собой в пределах точности измерений несмотря на то, что они определены разными инструментами и зондами; следовательно, им можно доверять так же, как и полученным бесспорным методом одноосного растяжения. Обычно наблюдается двух- трехкратное превышение прочности на растяжение над прочностью на сжатие или твердостью [93–96], в то время как в сплошных материалах имеет место обратное их соотношение (например, в металлах действует соотношение Тейбора – твердость превышает предел текучести примерно в 3 раза). По нашему мнению, это различие обусловлено спецификой поведения нанофибрилл при испытании на растяжение и сжатие или при измерении твердости. В первом случае молекулярные цепи вытягиваются и частично ориентируются вдоль оси волокна, что приводит к их упрочнению. При индентировании (как с помощью НИ, так и АСМ) превалируют напряжения сжатия, под действием которых микрофибриллы изгибаются и теряют устойчивость значительно раньше, чем разрушаются при одноосном растяжении.
Таблица 3.
Значения величины модуля Юнга Е в разных микроволокнах, полученные различными методами на трех материалах [94]
| Материал | Растяжение, ГПа | Наноиндентирование, ГПа | Картирование, контактная мода AFM, ГПа |
|---|---|---|---|
| Лен | 68.9 ± 24.6 | 20.4 ± 1.1 | 21.3 ± 2.2 |
| Бамбук | 43.6 ± 0.6 | 21.3 ± 1.7 | 21.3 ± 2.9 |
| Дерево | 18–40 | 14–20 | 11 |
КЛЕТКИ И КЛЕТОЧНЫЕ СТЕНКИ
Микроволокна целлюлозы в результате взаимодействия с другими компонентами (лигнин, гемицеллюлоза, пектин, вода и др.) формируют клеточные стенки (рис. 1). Различают первичную стенку и многослойную вторичную, последняя вносит основной вклад в жесткость и прочность клетки. По мере перехода от EW к LW размеры клеток несколько уменьшаются, толщина клеточных стенок нарастает, а внутреннее сечение капилляров падает.
Результаты, полученные методом НИ, проиллюстрируем на нескольких типичных примерах. Так, в [73] авторы анализировали зависимости продольного модуля Юнга Е и нанотвердости Н клеточных стенок от положения клетки в двух годовых кольцах сосны обыкновенной (Pinus sylvestris L.), соответствующих возрасту 7 лет и 74 года. Из (рис. 5а) следует, что величина Е при переходе из EW в LW увеличивается примерно на 50%, а величина Н – всего на 6–7% (рис. 5б). В ряде других работ приводятся схожие данные, свидетельствующие о том, что в разных слоях и годовых кольцах и даже в различных породах дерева нанотвердость клеточных стенок отличается не очень сильно. Так, в [97] в сосне (Pinus massoniana Lamb.) нанотвердость клеточной стенки варьировалась в интервале 0.35–0.42 ГПа. Авторами [98] найдено, что твердость клеточной стенки в древесине сосны Masson pine составляла 0.41–0.53 ГПа и в пределах рассеяния результатов не отличалась для EW и LW, а авторами [99] в сосне ладанной (Pinus taeda) Н = 0.34–0.54 ГПa (без дифференцирования на EW и LW).
Рис. 5.
Зависимости модуля упругости Еn (а) и нанотвердости Нn (б) клеточной стенки от номера клетки в годовом слое для двух годовых колец, соответствующих возрасту 7 лет (синие значки) и 74 года (красные значки) [73].

В [100, 101] максимальная нагрузка Pmax, приложенная к индентору, была выбрана равной 2 мН, что обеспечивало образование отпечатков глубиной h, в несколько раз превышающей параметр шероховатости Ra ~ 200 нм поверхности образца, подготовленной механической шлифовкой, и латеральный размер зоны деформации порядка поперечника клетки. Измеренные таким образом значения Ee и He можно рассматривать как эффективные для данного слоя древесины (как и определяемые в любых макроиспытаниях пористых тел, в частности, древесины) ввиду отмеченного выше соотношения размеров отпечатка и поперечного размера клеток. Основные экспериментальные результаты по определению радиальной зависимости Ee и He в дубе черешчатом (Quercus robur) представлены на рис. 6а, а в липе мелколистной (Tilia cordata) – на рис. 6б. Каждая точка на графиках есть результат усреднения от 10 до 20 индивидуальных измерений. Переход от структуры EW к LW может происходить скачкообразно, как в дубе (рис. 6а), или плавно, как в липе (рис. 6б). Разница между механическими свойствами в конце стадии роста и в начале роста в следующем году составляла несколько сотен процентов. Это позволяет весьма точно определить толщину колец и механические свойства в каждом из них, а затем связать с условиями роста не только по годам, но и внутри сезона роста, что недоступно традиционным оптическим методам исследования годовых колец.
Рис. 6.
Зависимости Нe и Еe при наноиндентировании дуба (а) и липы (б) от расстояния r поперек годовых колец (для 4 последовательных годовых колец). Максимальная нагрузка, приложенная к индентору Берковича, Рmax = 2 мН. Границы годовых колец показаны пунктирными линиями.

Важно отметить, что предложенная методика сканирования без предварительного отбора точек индентирования и прицеливания в клеточные стенки, выбранная величина Pmax = 2 мН и упрощенная процедура подготовки поверхности образца к измерениям, оставляющая часть сошлифованных микроволокон в капиллярах, не только не мешают определению эффективных значений Ee и He, но и имеют ряд преимуществ перед описанными ранее методами измерения E и H в отдельных клеточных стенках при меньших Pmax (см., например, [16, 73], где НИ осуществлялось при Pmax = 0.2–0.8 мН): 1) методика позволяет высокопроизводительно инспектировать большие площади (~104 мм2 и более) вместо единиц мм2 при подготовке их с помощью микротома; 2) соотношение Ee и He в LW и EW намного выше, чем соотношение величин E и H в клеточных стенках; 3) наконец, определение эффективных значений Ee и He с учетом пористости p сближает их с макро характеристиками древесины и дает возможность предсказания последних (при необходимости) без независимого определения величины p. Отмеченное выше означает, что измерение эффективных значений Ee и He может быть гораздо более чувствительным методом дендрохронологии и дендроклиматологии, чем определение по локальным значениям E и H в клеточных стенках и измерению вариаций ширины колец роста.
Таким образом, приведенные данные о величине Е и Н в наношкале свидетельствуют о близости структуры клеточных стенок в различных слоях и породах древесины. Их значения выше измеренных в [100, 101] эффективных микроскопических Еe и Нe, и тем более табличных макроскопические характеристик [102–104]. Формально это можно расценивать как проявление размерного эффекта (РЭ) в механических свойствах древесины. Однако вопрос, какую часть этого РЭ можно отнести на счет сплошного материала древесины, а какую на счет пор, требует отдельного исследования.
РАЗМЕРНЫЕ ЭФФЕКТЫ В ДРЕВЕСИНЕ
В отношении РЭ в древесине на разных масштабно-иерархических уровнях имеется не так много информации. Приведем наиболее интересные и типичные данные. Расчетная и экспериментально определенная прочность элементарных целлюлозных нанофибрилл диаметром 3–15 нм составляет 4.9–10 ГПа и превышает прочность микроволокон диаметром 8–12 мкм от 5 до 10 раз. Как следует из рис. 6а и 6б эффективные значения Н, как в слоях EW, так и LW, были в несколько раз ниже (от 2 до 4) нанотвердости клеточных стенок с типичной толщиной 2–5 мкм [59, 65] и в 5–10 раз меньше, чем прочность микроволокон целлюлозы (0.8–1.57 ГПа) с поперечными размерами 8–20 мкм. Вместе с тем эффективная величина Нe была в несколько раз выше макротвердости НВ по Бринеллю и прочности на одноосное растяжение, полученных в макроиспытаниях [102–104].
Ясно, что помимо внутренних причин, определяемых молекулярной и надмолекулярной структурой, на падение эффективного модуля Юнга и твердости на мезо- и макроуровнях большое влияние оказывает еще и нано-/микро- пористость древесины, обусловленная наличием в структуре любой древесины пор, капилляров и более крупных трахеид с большим аспектным соотношением. Их присутствие приводит к нескольким существенным отличиям в механическом поведении древесины от сплошных макротел. Во-первых, практически никогда не выполняется правило Тейбора, согласно которому твердость мягких материалов должна превышать предел текучести или прочности примерно в 3 раза. Напротив, в большинстве пород древесины макротвердость, как правило, в несколько раз ниже пределов текучести и прочности на растяжение. По-видимому, во всех подобных явлениях причиной является потеря устойчивости ячеистой структуры древесины при индентировании и одноосном сжатии, которая наступает гораздо раньше, чем неупругая деформация и разрушение при растяжении. Ввиду анизотропии механических свойств древесины эти явления сильно зависят от направления приложения нагрузки по отношению к длинной оси клеток. В продольном направлении они примерно на порядок выше, чем в поперечном. Как бы то ни было, из приведенных выше данных следует, что в древесине наблюдаются яркие РЭ, вследствие чего прочность/твердость падает от ~10 ГПа в нанокристаллической целлюлозе до ~0.1 ГПа в макрообъемах древесины. Это означает, что любые целлюлозосодержащие материалы имеют большой потенциал упрочнения, который можно реализовать с помощью оптимально организованной нано- и микро- структуры и соответствующих технологий.
Для многих областей применения – авиация, космонавтика, автомобилестроение, спортивный инвентарь и др. решающее значение имеет не абсолютные, а удельные механические характеристики, отнормированные на плотность материала ρ. Бездефектная наноцеллюлоза может более чем на порядок величины превышать характеристики алюминиевых сплавов и конструкционных сталей по σbρ–1 (рис. 7), уступая им только по деформации до разрушения и трещиностойкости. Конкурентами наноцеллюлозы по удельным прочностным характеристикам являются лишь некоторые полимерные микроволокна (углеродные, пара-арамидные – Кевлар). Одностенные углеродные нанотрубки (SWCNT) и графены многократно превосходят любые известные материалы по удельным механическим показателям.
Рис. 7.
Механические характеристики наноцеллюлозы и микроволокон целлюлозы в сравнении с характеристиками наиболее распространенных существующих и перспективных конструкционных материалов. Заштрихованные поля – абсолютные характеристики (модуль Юнга Е и прочность на растяжение σb), незаштрихованные – удельные характеристики прочности материалов, нормированные на их плотность ρ. 1 – Пластик, 2 – древесина, 3 – композиты, 4 – металлы и сплавы, 5 – керамика, 6 – кевлар, 7 – микроцеллюлоза, 8 – наноцеллюлоза, 9 – одностенные углеродные нанотрубки.

Представление прочностных характеристик в функции характерных размеров структуры R* в двойных логарифмических координатах дает характерную диаграмму в виде хоккейной клюшки (рис. 8). Нисходящая часть этой зависимости имеет наклон, близкий к 0.5, что роднит ее с известным в физическом материаловедении соотношением Холла–Петча: σy = σ0 + A(R*)–0.5, где σy – предел текучести, σ0 и А – константы материала. Разумеется, причины падения прочности с ростом R* в разных группах материалов могут быть различными, тем не менее, имеются и общие черты. В макрошкале тоже может наблюдаться размерная зависимость, но гораздо более слабая, чем в нано- и микромасштабной области. Она объясняется ростом вероятности появления крупных дефектов с увеличением объема объекта и описывается статистической теорией Вейбула, но ее рассмотрение лежит за пределами тематики настоящего обзора. Механизмы разупрочнения с ростом R* требуют дополнительного изучения связей нано-/микроструктуры со свойствами.

КОМПОЗИТЫ С ЦЕЛЛЮЛОЗНЫМИ АРМИРУЮЩИМИ ВОЛОКНАМИ
На основе древесины создают и более сложно устроенные древесно-полимерные композиты, в частности, армированные хаотически расположенными или ориентированными волокнами, со слоистой или сотовой структурой и архитектурой. Набирают популярность и материалы, состоящие целиком из модифицированных растительных компонентов. В последние годы по совокупности причин, описанных во введении, интерес к композитам с природными наполнителями и армирующими элементами сильно возрос. Свидетельством тому – большое количество появившихся новых материалов и обзоров, опубликованных в последние год–два [105–114]. Основные проблемы, которые приходится решать при создании таких композитов, заключаются в выборе сырья, технологии его обработки, выделения и упорядочении волокон, обеспечении хорошей адгезии к матрице и создании запланированной архитектуры при соблюдении требований к экологии и безопасности производства, последующего использования и утилизации. Всё это должно находиться в балансе с механическими и служебными свойствами, а также со стоимостью. Биоразлагаемые полимерные матрицы, к сожалению, сами по себе имеют невысокую прочность, и их не удается сильно упрочнить даже высокопрочными волокнами.
ЗАКЛЮЧЕНИЕ
Несмотря на то, что древесина различных пород и ее важнейший компонент – целлюлоза служат человечеству уже многие тысячи лет, природа их свойств до конца не раскрыта, и потенциал этого уникального природного материала используется далеко не в полной мере. Для прогнозирования характеристик древесины, целенаправленного их изменения в процессе выращивания или модификации с помощью различного вида обработок, а также сохранения в процессе эксплуатации необходимо понимание того, какие элементы структуры древесины, их состояние, ориентация, взаимодействие и эволюция, а также дефекты и пороки нано-, микро-, мезо- и макроструктуры ответственны за те или иные макроскопические свойства. В первую очередь это требует знания таких свойств у наиболее важных и общих, ключевых элементов микроструктуры древесины – нанокристаллических и аморфных фаз целлюлозы, микрофибрилл, клеточных стенок, годовых колец роста.
Разнообразные современные методы исследования нано- и микроструктуры, а также сканирующее индентирование с разной максимальной нагрузкой позволяет получать мультимасштабные данные о механических свойствах древесины на различных структурных уровнях – от нано- до макро-. Сопоставление детальной информации о распределении механических характеристик внутри годового кольца, а затем в соседних кольцах роста с макрохарактеристиками позволяет лучше понять природу и механизмы формирования последних. В свою очередь это может дать новые подходы к оптимизации условий выращивания древесины с заранее заданными механическими свойствами, например, с высокой прочностью и упругостью, необходимыми акустическими характеристиками или низкой скоростью ползучести, а также физически обоснованных методов упрочнения древесины.
Мультимасштабное изучение связи структуры и свойств целлюлозосодержащих материалов создает предпосылки для поиска новых подходов к дизайну композиционных материалов следующего поколения, выявления внутренних связей различных свойств (например, механических и тепловых). Высокая удельная прочность наноцеллюлозы, превышающая таковую практически у всех современных конструкционных материалов, кроме наноуглеродных, ставит задачу использования этого потенциала в создании новых высокопрочных экологически чистых материалов. Результаты сканирования нано- и микромеханических свойств древесины, охватывающего последовательный ряд годовых колец роста, создают основу для разработки инновационных методов и средств дендрохронологии и дендроклиматологии, дополняющих традиционные. Поскольку средний размер клеток в поперечном срезе древесины составляет 30–50 мкм, а средняя толщина годового кольца 1–3 мм, в нем укладывается 50–100 клеток. В принципе, наноиндентирование позволяет измерять механические характеристики каждой клетки. Следовательно, предел временного разрешения дендохронологического применения метода НИ может составлять около 1 недели.
Работа частично выполнена в Центре коллективного пользования ТГУ им. Г.Р. Державина при финансовой поддержке Российского научного фонда (проект № 21-14-00233) и Министерства науки и высшего образования Российской Федерации в рамках проекта по соглашению № 075-15-2021-709 (уникальный идентификатор проекта RF-2296.61321X0037).
Список литературы
Wang J., Wang L., Gardner D.J. et al. // Cellulose. 2021. V. 28. P. 4511.
Wood handbook—wood as an engineering material. General Technical Report FPL-GTR-282. Madison: U.S. Department of Agriculture, Forest Service, Forest Products Laboratory, 2021. 543 p.
Plocher J., Mencattelli L., Narducci F., Pinho S. // Compos. Sci. Technol. 2021. V. 208. Art. No. 108669.
Zhu J.Y., Agarwal U.P., Ciesielski P.N. et al. // Biotechnol. Biofuels. 2021. V. 14. P. 114.
Sandberg D., Kutnar A., Karlsson O., Jones D. Wood modification technologies. Principles, sustainability, and the need for innovation. CRC Press. Taylor & Francis Group, 2021. 431 p.
Pandey K.K., Ramakantha V., Chauhan S.S., Kumar A.N.A. Wood is good. Current trends and future prospects in wood utilization. Springer Nature Singapore Pte Ltd, 2017. 480 p.
Swingler S., Gupta A., Gibson H. et al. // Polymers. 2021. V. 13. P. 412.
Raut H.K., Das R., Ziqian L. et al. // Biotechnol. J. 2020. V. 15. No. 12. Art. No. e2000160.
Mali P., Sherje A.P. // Carbohydr. Polym. 2022. V. 275. Art. No. 118668.
Fratzl P., Weinkamer R. // Prog. Mater. Sci. 2007. V. 52. P. 1263.
Gibson L.J. // J. Royal Soc. Interface. 2012. V. 9. P. 2749.
Song J., Chen C., Zhu S. et al. // Nature. 2018. V. 554(7691). P. 224.
Chen C., Kuang Y., Zhu S. et al. // Nature. Rev. Mater. 2020. V. 5. No. 9. P. 1.
Donaldson L.A. // IAWA J. 2019. V. 40. No. 4. P. 645.
Johansson C.J. // In: Timber engineering. London: Wiley, 2003. P. 23.
Porteous J., Kermani A. Structural timber design to Eurocode 5. Malden: Blackwell Publishing Ltd, 2007. 555 p.
Kretschmann D.E. // In: Wood handbook – Wood as an engineering material. General Technical Report FPL–GTR–190. Madison: Forest Products Laboratory. United States Department of Agriculture Forest Service, 2010. P. 5-1.
Olorunnisola A.O. Design of structural elements with tropical hardwoods. Springer Int. Publishing AG, 2018. 288 p.
Jahan Z., Niazi M.B.K., Gregersen O.W. // J. Ind. Engin. Chem. 2018. V. 57. P. 113.
Glukhikh V. // Architect. Engin. 2021. V. 2. No. 1. P. 24.
Burawska-Kupniewska I., Krzosek S., Mankowski P. et al. // BioResources. 2019. V. 4. No. 14. P. 9287.
Moon R.J., Martini A., Nairn J. et al. // Chem. Soc. Rev. 2011. V. 40. No. 7. P. 3941.
Kargarzadeh H., Ahmad I., Thomas S., Dufresne A. Handbook of nanocellulose and cellulose nanocomposites. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA, 2017. 849 p.
Baghaei B., Skrifvars M. // Molecules. 2020. V. 25. Art. No. 2836.
Reza M., Kontturi E., Jääskeläinen A.-S., Vuorinen T. // Bioresources. 2015. V. 10. No. 3. P. 6230.
Cascione M., De Matteis V., Rinaldi R., Leporatti S. // Microsc. Res. Technol. 2017. V. 80. P. 109.
Balzano A., Novak K., Humar M., Čufar K. // Les/Wood. 2019. V. 68. No. 2. P. 5.
Broda M., Popescu C.-M. // Spectrochim. Acta. A. 2019. V. 209. P. 280.
Xia Y., Ma C., Wang H. et al. // Polymers. 2020. V. 12. P. 237.
Papadopoulos A.N., Bikiaris D.N., Mitropoulos A.C., Kyzas G.Z. // Nanomaterials. 2019. V. 9. P. 607.
Moghaddam M.S., den Bulcke J.V., Wålinder M.E.P. // Holzforschung. 2017. V. 71. No. 2. P. 119.
Thomas J., Collings D.A. // In: Wood is good. Current trends and future prospects in wood utilization. Springer Nature Singapore Pte Ltd, 2017. P. 29.
Alves E.E.N., Rodriguez D.R.O., Rocha P.A. et al. // Results Chem. 2021. V. 3. Art. No. 100121.
Scharnweber T., Hevia A., Buras A. et al. // Sci. Total Environ. 2016. V. 566-567. P. 1245.
Guo F., Altaner C.M. // Carbohydr. Polym. 2018. V. 197. P. 1.
Tintner J., Spangl B., Reiter F. et al. // Wood Sci. Technol. 2020. V. 54. P. 313.
Özgenç Ö., Durmaz S., Boyaci I.H., Eksi-Kocak H. // Spectrochim. Acta A. 2017. V. 171. P. 395.
Popescu C.M., Jones D., Krzisnik D., Humar M. // J. Mol. Struct. 2020. V. 1200. Art. No. 127133.
Gierlinger N. // Appl. Spectrosc. Rev. 2018. V. 53. No. 7. P. 517.
Kanbayashia T., Kataoka Y., Ishikawa A. et al. // J. Photochem. Photobiol. B. 2018. V. 187. P. 136.
Saletnik A., Saletnik B., Puchalski C. // Molecules. 2021. V. 26. Art. No. 1537.
Szymanska-Chargot M., Chylinska M., Pieczywek P.M. et al. // Planta. 2016. V. 243. P. 935.
Kang X., Kirui A., Widanage M.C.D. et al. // Nature Commun. 2019. V. 10. Art. No. 347.
Elsayad K., Urstoger G., Czibula C. et al. // Cellulose. 2020. V. 27. P. 4209.
Toumpanaki E., Shah D.U., Eichhorn S.J. // Adv. Mater. 2020. Art. No. 2001613.
Bhushan B. Nanotribology and nanomechanics. An introduction. Berlin–Heidelberg–N.Y.: Springer, 2008. 1516 p.
Tiwari A. Nanomechanical analysis of high performance materials. Dordrech–Heidelberg–N. Y.–London: Springer Science + Business Media, 2014. 348 p.
Ranganathan N.M. Materials characterization: modern methods and applications. Boca Raton: CRC Press, 2015. 334 p.
Головин Ю.И. // ФТТ. 2021. Т. 63. № 1. С. 3; Golovin Yu.I. // Phys. Solid State. 2021. V. 63. P. 1.
Kargarzadeh H., Ahmad I., Thomas S., Dufresne A. Handbook of nanocellulose and cellulose nanocomposites. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA, 2017. 849 p.
Hsueh C.-H., Schmauder S., Chen C.-S., Chawla K.K. Handbook of mechanics of materials. Springer Nature. Singapore Pte. Ltd, 2019. 2431 p.
Khalili A.A., Ahmad M.R. // Int. J. Mol. Sci. 2015. V. 16. Art. No. 18149.
Cascione M., De Matteis V., Rinaldi R., Leporatti S. // Microsc. Res. Technol. 2017. V. 80. P. 109.
Головин Ю.И., Иволгин В.И., Коренков В.В. и др. // Конд. среды и межфаз. границы. 2001. Т. 3. № 2. С. 122.
Головин Ю.И. // ФТТ. 2008. Т. 50. № 12. С. 2113; Golovin Yu. I. // Phys. Solid State. V. 50. No. 12. P. 2205.
Головин Ю.И. // Завод. лаб. 2009. Т. 75. № 1. С. 45.
Головин Ю.И. Наноиндентирование и его возможности. М: Машиностроение, 2009. 312 с.
Fischer-Cripps A.C. Nanoindentation. N.Y.: Springer, 2011. 282 p.
Oyen M.L. Handbook of nanoindentation with biological applications. Pan Stanford Publishing Pte. Ltd, 2011. 359 p.
Nemecek J. Nanoindentation in materials science. London: InTech, 2012. 318 p.
Tiwari A. Nanomechanical analysis of high performance materials. Dordrech–Heidelberg–N.Y.–London: Springer Science+Business Media, 2014. 348 p.
Tiwari A., Natarajan S. Applied nanoindentation in advanced materials. N.Y.: John Wiley & Sons, 2017. 689 p.
Lee H.-R., Kim K.H., Mun S.C. et al. // Carbohydr. Polym. 2018. P. 276.
Shamsuddin S.-R., Lee K.-Y., Bismarck A. // Composites A. 2016. V. 90. P. 633.
Eichhorn S.J., Dufresne A., Aranguren M. et al. // J. Mater. Sci. 2010. V. 45. No. 1. P. 1.
Mittal N., Ansari F., Gowda K. et al. // ACS Nano. 2018. V. 12. No. 7. P. 6378.
Ioelovich M. // In: Handbook of nanocellulose and cellulose nanocomposites Weinheim: Wiley-VCH Verlag GmbH & Co, 2017. P. 51.
Burgert I., Keplinger T. // J. Exper. Botany. 2013. V. 64. No. 15. P. 4635.
Eder M., Arnould O., Dunlop J.W.C. // Wood Sci. Technol. 2013. V. 47. P. 163.
Rongpipi S., Ye D., Gomez E. D., Gomez E.W. // Front. Plant Sci. 2019. V. 9. Art. No 1894.
Nelson N., Stubbs C.J., Larson R., Cook D.D. // J. Exper. Botany. 2019. V. 70. No. 4. P. 3649.
Perepelkin N.V., Borodich F.M., Kovalev A.E., Gorb S.N. // Biolog. Biomimet. Mater. Nanomat. 2020. V. 10. Art. No. 15.
Mania P., Nowicki M. // Bull. Polish Acad. Sci. Tech. Sci. 2020. V. 68. No. 5. P. 1237.
Normand A.C., Charrier A.M., Arnould O., Lereu A.L. // Sci. Rep. 2021. V. 11. Art. No. 5739.
Vaganov E.A., Hughes M.K., Shashkin A.V. Growth dynamics of conifer tree rings. images of past and future environments. Berlin-Heidelberg: Springer, 2006. 354 p.
Samusevich A., Lexa M., Vejpustková M. et al. // Dendrochronologia. 2020. V. 60. Art. No 125686.
Rubino D.L., McCarthy B.C. // Dendrochronologia. 2004. V. 21. P. 97.
Heinrich I. Dendrogeomorphology. The encyclopedia of quaternary science. Amsterdam: Elsevier, 2013.
Helama S., Lindholm M., Timonen M., Eronen M. // Theor. Appl. Climatol. 2004. V. 79. P. 239.
Meko D.M., Friedman J.M., Touchan R. // Holocene. 2015. V. 25. P. 1093.
Gärtner H., Cherubini P., Fonti P. et al. // J. Vis. Exp. 2015. V. 97. Art. No. e52337.
Zhang X., Li J., Liu X., Chen Z. // J. Forest Res. 2019. V. 31. No. 2. Art. No. 1002.
Arbellay E., Jarvis I., Chavardès R.D. et al. // Tree Physiology. 2018. V. 38. P. 1237.
Zhang W., Zhao T., Su X. et al. // Forests. 2021. V. 12. P. 464.
Bjorklund J., von Arx G., Fonti P. et al. // Dendrochronologia. 2021. V. 69. Art. No. 125880.
Mayer K., Grabner M., Rosner S. et al. // Dendrochronologia. 2020. V. 64. Art. No. 125781.
Biondi F. // Forests. 2020. V. 11. Art. No. 146.
Jacquin P., Longuetaud F., Leban J.-M., Mothe F. // Dendrochronologia. 2017. V. 42. P. 42.
https://forestrypedia.com/physical-and-mechanicalproperties-of-wood.
Bourmaud A., Deaugrand J., Shah D.U. et al. // Progr. Mater. Sci. 2018. V. 97. P. 347.
Lee H.-R., Kim K.H., Mun S.C. et al. // Carbohydr. Polym. 2018. V. 180. P. 276.
Chen C., Kuang Y., Zhu S. et al. // Nature Rev. Mater. 2020. V. 5. No. 9. P. 642.
Zarna C., Opedal M. T., Echtermeyer A.T., Chinga-Carrasco G. // Composites C. 2021. V. 6. Art. No. 100171.
Tanguy M., Bourmaud A., Baley C. // Mater. Lett. 2016. V. 167. P. 161.
Tayeb A.H., Amini E., Ghasemi S., Tajvidi M. // Molecules. 2018 V. 23. Art. No. 2684.
Vincent M., Tong Q., Terziev N. et al. // Wood Sci. Technol. 2014. V. 48. No. 1. P. 7.
Wu Y., Wu X., Yang F. et al. // Forests. 2020. V. 11. Art. No. 1247.
Huang Y.H., Fei B.H., Yu Y. et al. // Bioresources. 2012. V. 7. No. 3. P. 3028.
Tze W.T.Y., Wang S., Rials T.G. et al. // Composites A. 2007. V. 38. P. 945.
Головин Ю.И., Тюрин А.И., Головин Д.Ю. и др. // Изв. вузов. Физ. 2020. Т. 63. № 11(755). С. 187; Golovin Y.I., Tyurin A.I., Golovin D.Y. et al. // Russ. Phys. J. 2021. V. 63. № 11. P. 2041.
Головин Ю.И., Тюрин А.И., Гусев А.А. и др. // Письма в ЖТФ. 2022. Т. 48 № 4. С. 36.
https://lesoteka.com/.
https://extxe.com/.
https://les.novosibdom/.
Chen C., Kuang Y., Zhu S. et al. // Nature Rev. Mater. 2020. V. 5. No. 9. P. 1.
Heise K., Kontturi E., Allahverdiyeva Y. et al. // Adv. Mater. 2021. V. 33. No. 3. Art. No. e2004349.
Isogai A. // Adv. Mater. 2021. V. 33. No. 28. Art. No. e2000630.
Thomas P., Duolikun T., Rumjit N. P. et al. // J. Mech. Behav. Biomed. Mater. 2020. V. 110. Art. No. 103884.
Yang X., Biswas S.K., Han J. et al. // Adv. Mater. 2021. V. 33. No. 28. Art. No. e2002264.
Niinivaara E., Cranston E.D. // Carbohydr. Polym. 2020. V. 247. Art. No. 116664.
Li C., Wu J., Shi H. et al. // Adv. Mater. 2021. Art. No. 2105196.
De France K., Zeng Z., Wu T., Nyström G. // Adv. Mater. 2021. V. 33. Art. No. 2000657.
Trache D., Tarchoun A. F., Derradji M. et al. // Front. Chem. 2020. V. 8. P. 392.
Mahmud S., Hasan K. M. F., Jahid A. et al. // J. Mater. Sci. 2021. V. 56. P. 7231.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия физическая