

Химическая физика, 2021, T. 40, № 11, стр. 40-47
Деструкция оболочек вирусов гриппа А гетерополикислотами Кеггина
С. А. Ковалевский 1, О. А. Лопатина 2, Е. А. Гущина 2, Е. И. Исаева 2, И. Т. Федякина 2, О. В. Бакланова 2, М. В. Мезенцева 2, Е. М. Балашов 1, *, Н. М. Ивашкевич 3, А. И. Кулак 3, Ф. И. Далидчик 1
1 Федеральный исследовательский центр химической физики им. Н.Н. Семёнова,
Российской академии наук
Москва, Россия
2 Национальный исследовательский центр эпидемиологии и микробиологии им. Н.Ф. Гамалеи
Москва, Россия
3 Институт общей и неорганической химии Национальной Академии наук Беларуси
Минск, Беларусь
* E-mail: embalashov@yandex.ru
Поступила в редакцию 03.12.2020
После доработки 09.03.2021
Принята к публикации 22.03.2021
Аннотация
Приведены результаты электронно-микроскопического исследования действия гетерополикислот (ГПК) со структурой Кеггина на частицы вирусов гриппа А/Aichi/1/68 (Н3N2) и A/California/07/09 (Н1N1) pdm09. Показано, что действие ГПК на вирусные частицы приводит к полному (или частичному) удалению трансмембранных гликопротеинов и разрушению матриксного белка М1, которое проявляется в деформациях и разрушениях вирусных оболочек. На примере вирусов A/California/07/09 (Н1N1) pdm09 показано, что эффективность разрушения оболочки вирусов гетерополикислотами зависит от среды культивирования вирусов. Приводится механизм деструкции, предполагающий экстракцию холестерина, травление фосфолипидов и образование пор в липидной оболочке в результате действия гетерополианионов. Предполагается, что проникновение протонов через образовавшиеся поры может приводить к разрушению матриксного белка М1.
ВВЕДЕНИЕ
Быстрые мутации вирусов приводят к резистентности известных противовирусных препаратов. Поэтому идут непрерывные поиски новых лекарственных средств. Одними из этих средств могут быть полиоксометаллаты (ПОМ) [1, 2]. Среди неорганических биологически активных соединений ПОМ занимают особое место. Их производство относительно биологических препаратов менее дорого, а возможности варьирования структуры, химического состава и свойств очень велики. Полиоксометаллаты хорошо известны своими противораковым и противовирусными свойствами [3, 4]. Наиболее эффективными для борьбы с вирусами являются ПОМ со структурами Кеггина и Доусона. Синтезированы десятки ПОМ [5–8], активность которых была проверена на различных вирусах. Показано, что ПОМ со структурой Кеггина проявляют высокую антивирусную активность в отношении ряда оболочечных РНК вирусов [3, 9]. Несмотря на большое количество работ по исследованию механизма противовирусного действия, он остается малоизученным. Показано [9], что одни ПОМ могут ингибировать связывание вируса с клеткой, другие ингибируют проникновение вируса сквозь мембрану. Для доказательства ингибирования ПОМ взаимодействия вирусов с липидной оболочкой ранее использовался метод флуоресценции [8]. Этот метод применяется для изучения липидных пленок и в настоящее время [10]. Прямое действие ПОМ на оболочку вирусов гриппа не изучалось. Предположение о том, что ПОМ со структурой Кеггина разрушает оболочку, делалось в работе [11] в отношении вирусов гипатита С. До настоящего времени прямых подтверждений разрушения оболочки вирусов гриппа в результате действия ПОМ нет. Гетерополикислоты являются частным случаем полиоксометаллатов и изучаются как противовирусные и противораковые препараты [3].
Цель настоящей работы состояла в изучении действия гетерополикислот (ГПК) со структурой Кеггина на оболочку вирусов гриппа. Был применен метод электронной микроскопии, с помощью которого определены дефекты оболочки вирусов гриппа А/Aichi/1/68 (Н3N2), вызванные действием следующих ГПК: H3PW12O40, H3PMo12O40, H4SiW12O40, H4SiMo12О40. Ранее было показано, что эти ГПК эффективно подавляют гемагглютинирующую активность вирусов гриппа А/Aichi/1/68 (Н3N2), но они неэффективны в отношении штамма A/California/07/09 (Н1N1) pdm09 [12].
В работах [13–15] была отмечена высокая противовирусная активность гетерополианионов, содержащих атомы ванадия. Они оказывают ингибирующее действие на различные стадии инфицирования клеток вирусами, но являются более токсичными. Их действие на оболочку вирусов не изучалось, но можно предположить, что эти ГПК будут эффективны в отношении вирусов A/California/07/09 (Н1N1) pdm09. Для этого, а также для решения вопроса об устойчивости вирусов к действию ГПК в зависимости от липидного состава и строения оболочки были проведены эксперименты с применением ванадийзамещенных ГПК: H5PMo10V2O40, H4PMo11VO40, H4PW11VO40.
Можно предположить, что эффективность деструкции оболочки вирусов после воздействия ГПК должна зависеть от состава внешней липидной оболочки вирусов, которая отражает состав мембраны клетки хозяина, и от ассоциированного с ней матриксного каркаса, образованного белком М1. Чтобы проверить, как действуют ГПК на вирусы с разным составом липопротеидной оболочки, были выбраны вирусы, культивированные на куриных эмбрионах и на клетках МДСК (клетки почки собаки). Эти вирусы имеют сильно различающийся состав липидных оболочек. Состав оболочки вирусов гриппа, культивированных на куриных эмбрионах, хорошо изучен (см. обзор [16]). В нем преобладают фосфолипиды, а содержание холестерина достигает 35%. Напротив, основным липидом в составе оболочки вирусов, культивированных на культуре клеток МДСК, является холестерин. В работе [17], в которой применялся метод масс-спектрометрии, показано, что содержание холестерина в оболочке вирусов гриппа А может достигать 52%, что существенно больше его содержания в самой клетке (28%) и больше содержания в апикальной части поверхности мембраны (48%). Содержание глицерофосфолипидов в оболочке не превышает 30%, еще меньше содержание сфингофосфолипидов: ~18%, в состав которых входит сфингомиелин. Холестерин и сфингомиелин являются одними из основных липидов мембраны вирусов, образующих рафты – участки оболочки вирусов с упорядоченной структурой, играющих важную роль во многих важных процессах жизненного цикла вирусов. В рафтах закреплены трансмембранные гликопротеины: гемагглютинин (НА) и нейраминидаза (NA) [18]. От содержания холестерина в мембране вириона зависит инфекционная активность вирусов [19–22].
Прочность оболочки вируса зависит не только от содержания холестерина в липидной мембране, но и от строения матриксного белка М1, выстилающего внутреннюю часть двуслойной липидной мембраны вируса [23, 24]. С этим белком связаны цитоплазматические хвосты гликопротеинов, находящихся на поверхности [21]. Вирусы гриппа, культивированные на куриных эмбрионах, часто содержат структурные дефекты. В работе [25] методом криоэлектронной микроскопии было показано, что матрикс М1 может отслаиваться от липидной мембраны. В работах [26–28] обнаружено, что некоторые вирионы (вирусы гриппа X-31 A/Aichi/68 (H3N2)), культивированные на куриных эмбрионах, вообще не содержат белок М1, но сохраняют гликопротеины на поверхности. Количество таких вирионов может доходить до 10%. В случае вирусов, культивированных на клетках МДСК, такие структурные дефекты не наблюдались. Поэтому можно ожидать, что действие ГПК на вирусы, культивированные в разных средах, могут различаться.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Материалы
Вирусы. Вирусы штаммов гриппа А/Aichi/1/68 (Н3N2), гриппа A/California/07/09 (Н1N1) pdm09 и клетки МДСК получены из Государственной Коллекции вирусов и Коллекции клеточных культур при ФГБУ “НИЦЭМ им. Н.Ф. Гамалеи” Минздрава России.
Гетерополикислоты. Для исследования взяты гетерополикислоты Кеггина H3PW12O40, H3PMo12O40, H4SiW12O40, H4SiMo12О40 производства фирмы “Biochem” (Франция) и содержащие ванадий ГПК H5PMo10V2O40, H4PMo11VO40, H4PW11VO40, синтезированные в Институте общей и неорганической химии НАН Беларуси.
Методы
Методом электронной микроскопии изучено деструктивное действие ГПК на оболочку вирусов, культивированных на куриных эмбрионах, и на оболочку вирусов, культивированных на клетках МДСК.
1. Культивирование вирусов А/Aichi/1/68 (Н3N2) проводили в аллантоисной полости 9–10-дневных куриных эмбрионов в течение 48 ч при 36 °С. Исследование влияния ГПК на структуру вирионов гриппа проводили методом негативного контрастирования суспензии вируса. Содержащую вирусы гриппа суспензию в концентрации 5.5 lg TCID 50 в 1 мл смешивали с водным раствором ГПК.. Кислотность среды pН после добавления ГПК составляла 6.5. Смесь инкубировали в течение 1 ч при температуре 25 °С. Суспензию вирусных частиц, обработанную образцами ГПК, наносили на гальваническую сетку с формваровой подложкой, высушивали и контрастировали 1%-ным водным раствором уранилацетата. Образцы изучали в трансмиссионном электронном микроскопе JEOL 100XC. Подготовку образцов, содержащих частицы вирусов A/California/07/09 (Н1N1) pdm09, проводили аналогичным методом.
2. Влияние ГПК на структуру вируса гриппа A/California/07/09 (Н1N1) pdm09 в культуре клеток МДСК, зараженной вирусом гриппа А, исследовали методом ультратонких срезов. Клетки МДСК культивировали в 24-луночных планшетах в питательной среде Игла МЕМ с двойным набором аминокислот с добавлением трипсина в концентрации 2 мкг/мл. Спустя 24 ч вносили 100 мкл среды с ГПК в конечной концентрации 100 мкМ, инкубировали в течение 2 ч при температуре 37 °С и инфицировали вирусом гриппа A/California/ 07/09 (Н1N1) pdm09 в дозе 0.1 ТCID 50 на клетку. После чего в течение 24 ч клетки культивировали в условиях CO2-термостата при 37 °С. Затем суспензию клеток центрифугировали и полученный осадок фиксировали в 2.5%-ном растворе глютаральдегида в 0.15 М фосфатном буфере. Электронно-микроскопическое изучение влияния ГПК на структуру вируса гриппа A/California/ 07/09 (Н1N1) в культуре клеток МДСК проводили методом ультратонких срезов с использованием общепринятой методики фиксирования, обезвоживания, пропитки и заливки материала в эпоновую смесь. Контрастирование ультратонких срезов осуществляли 5%-ным раствором урацилацетата и цитратом свинца. Изображения образцов получали с использованием электронного микроскопа JEOL 100XC фирмы “JEOL” (Япония).
РЕЗУЛЬТАТЫ
Вирусы гриппа А/Aichi/1/68 (Н3N2), культивированные на куриных эмбрионах, подвергались воздействию ГПК со структурой H3PW12O40, H3PMo12O40, H4SiW12O40, H4SiMo12О40. На рис. 1а приведено контрольное изображение этих вирусов. Полноценные вирусы (отмечены стрелкой) имеют сферическую форму. Наиболее крупные вирусные частицы слегка деформированы. Эти результаты соответствуют изображениям вирионов (Н3N2), полученных методом криоэлектронной микроскопии в [25, 28]. Действие ГПК на вирусы А/Aichi/1/68 (Н3N2) приводило к появлению большого числа частиц вирусов с поврежденной оболочкой. В качестве примера приведено изображение вирусов после воздействия гетерополикислот H4SiW12O40 (рис. 1б). Можно выделить две группы вирусных частиц с дефектами в оболочке: частицы, сохранившие сферическую форму, но частично утратившие гликопротеины (рис. 1в), и частицы, имеющие дефекты оболочки, похожие на глубокие вмятины, и остатки гликопротеинов (рис. 1г). Такие же изменения имели оболочки вирусов А/Aichi/1/68 (Н3N2) после действия гетерополикислот H3PW12O40, H3PMo12O40, H4SiMo12О40.
Рис. 1.
а – Контрольное изображение частиц вирусов гриппа A/Aichi/2/68 (H3N2), культивированных на куриных эмбрионах (стрелками указаны полноценные вирионы); б – изображение частиц тех же вирусов после воздействия на них H4SiW12O40 (100 мкМ); в – вирус, частично сохранивший гликопротеины; г – вирус с деформированной оболочкой, сохранивший отдельные гликопротеины.
Рис. 1.
Окончание.
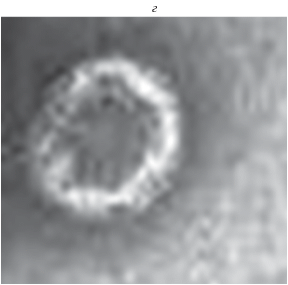
Исследование действия ГПК, содержащих атомы ванадия, проводили на вирусах гриппа А/California/07/09 (Н1N1). Для того чтобы изучить действие ГПК на оболочку вирусов с различающимся липидным составом, были проведены исследования с вирусами A/California/07/09 (Н1N1) pdm09, культивированными на куриных эмбрионах и на клетках МДСК. На рис. 2а приведено контрольное изображение вирусов А/California/07/09 Н1N1, культивированных на куриных эмбрионах. Здесь наблюдаются вирионы, имеющие сферическую форму, вытянутые частицы вирусов и частицы, имеющие дефекты структуры. Оболочки вирусов содержат гликопротеины.
Рис. 2.
а – Контрольное изображение частиц вирусов гриппа A/California/07/09 (Н1N1) pdm09, культивированных на куриных эмбрионах; б – изображение частиц вирусов гриппа A/California/07/09 (Н1N1) pdm09 после воздействия H5PMo10V2O40 (100 мкМ).

Как видно из рис. 2б, действие H5PMo10V2O40 на вирусы гриппа A/California/07/09 (Н1N1) pdm09 привело к появлению большой группы частиц с оболочками, лишенными гликопротеинов, потерявшими сферическую форму и имеющими продольные деформации. Одновременно с этими, структурно-дефектными частицами, на рис. 2б наблюдаются отдельные вирионы, сохранившие свою сферическую форму и гликопротеины на поверхности оболочки (обозначены стрелками). Эти частицы можно считать полноценными. Такое же действие оказывают другие ГПК, содержащие атомы ванадия. Потеря формы и образование продольных деформаций оболочки вирусов, наблюдаемые на рис. 2б, отличаются от дефектов частиц вирусов А/Aichi/1/68, показанных на рис. 1б. Из рис. 2б видно, что действие ГПК, содержащих атомы ванадия, на вирусы гриппа A/California/ 07/09 (Н1N1) pdm09, культивированные на куриных эмбрионах, приводит к деструкции оболочки большей части вирусов. В контрольных экспериментах вирионы, культивированные на куриных эмбрионах, чаще всего имеют сферическую форму, но также наблюдаются вытянутые вирусы (рис. 2а) и вирусы с формой, отличной от сферической [25–28]. После воздействия ГПК сохранившими гликопротеины остаются только частицы вирусов, имеющие сферическую форму. Поскольку на полученных в данной работе изображениях не наблюдаются полноценные частицы, имеющие структурные дефекты или отличающуюся от сферической форму, то можно сделать вывод, что такие частицы вирусов разрушаются в первую очередь.
Результаты электронно-микроскопического исследования действия ГПК, содержащих атомы ванадия (H4PMo11VO40, H5PMo10V2O40 и H4PW11VO40), на вирусы гриппа A/California/07/09 (Н1N1) pdm09, культивированные на клетках МДСК, значительно отличались от результатов, полученных для вирусов, культивированных на куриных эмбрионах. В контрольных экспериментах, в которых инфицированные клетки не подвергались действию ГПК, плазматическая мембрана не имела повреждений. Вирусы наблюдаются как на поверхности мембраны, так и в вакуолях внутри клетки (рис. 3а). В результате воздействия на инфицированные клетки МДСК гетерополикислотами, содержащими атомы ванадия, в концентрации, являющейся цитотоксической концентрацией СС50 для клеток (измеренные методом МТТ значения СС50 для H5PMo10V2O40, H4PMo11VO40 составляли 25 мкМ, а для H4PW11VO40 – 33 мкМ), часть клеток получала повреждения. Возле уцелевших клеток наблюдались полноценные вирусные частицы, не отличающиеся от контрольных. Изображение инфицированной клетки после воздействия на нее H4PMo11VO40 приведено на рис. 3г. Частицы вирусов, обозначенные стрелками, наблюдаются в межклеточном пространстве среди микроворсинок. При увеличении концентрации ГПК до 100 мкм бóльшая часть клеток погибала. Изображение такой разрушенной клетки МДСК приведено на рис. 3б. Из этого рисунка видно, что плазматическая мембрана клетки разрушена, микроворсинки отсутствуют, но заметных морфологических изменений частиц вирусов не наблюдается. Бóльшая часть вирусных частиц имеют сферическую форму и различимую структуру гликопротеинов. Действие ГПК на оболочку проявилось в виде разупорядочивания гликопротеинов на поверхности оболочки небольшого числа вирусов и в появлении участков поверхности, где наблюдается частичное отсутствие гликопротеинов (рис. 3в). Это сильно отличает вирусы, культивированные на клетках МДСК и подвергнутые действию ГПК, от вирусов, культивированных на куриных эмбрионах. Примерно такая же картина наблюдается при воздействии других ГПК: H5PMo10V2O40 и H4PW11VO40, на вирусы гриппа A/California/07/09 (Н1N1) pdm09. Наблюдалась деструкция клеток МДСК, но бóльшая часть частиц вирусов сохраняли сферическую форму.
Рис. 3.
а – Контрольное изображение частиц вирусов гриппа A/California/07/09 (Н1N1) pdm09, культивированных на клетках MДСК. Частицы вирусов на поверхности клетки и внутри вакуоли. б – Изображение клетки с разрушенной мембраной и частиц вирусов гриппа A/California/07/09 (Н1N1) pdm09, подвергнутых воздействию H4PMo11VO40 (100 мкМ). в – Увеличенный фрагмент, на котором наблюдаются вирусные частицы, на поверхности которых частично отсутствуют гликопротеины. г – Изображение вирусов гриппа A/California/07/09 (Н1N1) pdm09, культивированных на культуре клеток МДСК и подвергнутых воздействию H4PMo11VO40 (25 мкМ). Мембрана клетки не разрушена, около поверхности наблюдаются микроворсинки. Частицы вирусов (обозначены стрелками) не имеют заметных дефектов.

Рис. 3.
Окончание.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Из приведенных результатов следует, что действие ГПК на оболочку вирусов, культивированных на куриных эмбрионах, заметно отличается от их действия на вирусы, культивированные на клетках МДСК. Среди вирусов, культивированных на куриных эмбрионах, в первую очередь деструкции подвергаются те вирусы, которые имеют крупные размеры и несферическую форму. Как следует из данных работы [28], крупные и несферические частицы чаще всего имеют внутренние дефекты, такие как отсутствие матрикса М1, или толщину меньше нормальной. Хорошо известно, что к разрушению матрикса вирусов, культивированных на куриных эмбрионах, ведет повышение кислотности среды. Как было отмечено в работах [26, 27], при нормальной кислотности до 10% вирионов X-31 A/Aichi/68 (H3N2), культивированных на куриных эмбрионах, не содержат белок М1 в оболочке. При увеличении кислотности (до pH = 4.9) в течение 5 мин количество вирионов, не содержащих М1, увеличивается до 50%. Авторы объясняют это растворением матрикса М1 при проникновении протонов через канал М2. Это сказывалось на прочности закрепления гликопротеинов в оболочке. Гликопротеины переставали иметь вертикальное положение и могли смещаться. Также частицы вирусов с частично разрушенным матриксом М1 могут иметь участки поверхности без гликопротеинов [27].
Изображение, приведенное на рис. 1б, очень похоже на изображение частиц вирусов, подвергшихся изменениям в результате проникновения внутрь протонов. Но результаты наших экспериментов (кислотность среды после добавления ГПК рН ~ 6.5) трудно объяснить увеличением кислотности среды. Можно предположить другой механизм разрушения оболочки вируса. Известно, что снижение холестерина может приводить к образованию дефектов оболочки вирусов, таких как поры, что было обнаружено в экспериментах по определению действия на нее детергента, такого как метил-β-циклодекстрин. Образование пор приводило к проникновению красителя в частицы вирусов и регистрировалось методами электронной микроскопии. Это наблюдалось в случае различных вирусов, например вируса гриппа (A/WSN/33 H1N1) [29] и вирусов HIV-1 и SIV [30]. Изображения вирусов после воздействия на них метил-β-циклодекстрина, приведенные в этих работах, соответствуют изображению, приведенному на рис. 1г.
В работе [31] методом масс-спектрометрии было обнаружено, что действие гетерополианионов на плазматическую мембрану клеток фибробластов эмбриона человека приводит к снижению содержаний холестерина и сфингомалеина. Возможно, что и на оболочку вирусов ГПК действуют также. Это, как и в случае действия метил-β-циклодекстрина, может приводить к образованию пор. Нужно отметить, что экстракция связующих липидов, таких как холестерин, не является единственным механизмом разрушения оболочки вируса гетерополикислотами. Известен механизм действия гетерополианионов на липидную мембрану, состоящую из фосфолипидов [32–34]. Было обнаружено, что гетерополианионы электростатически связываются с фосфолипидами мембраны, например фосфатидихолином [32], имеющим положительно заряженную холиновую группу, образуя комплексы. Распадаясь, эти комплексы оставляют в мембране поры. При увеличении концентрации ПОМ происходит быстрое разрушение мембраны.
В случае действия токсичных ГПК, содержащих атомы ванадия, вероятно, происходит не только образование пор, но и разрушение матрикса М1. Это видно на примере действия H5PMo10V2O40 на вирусы гриппа A/California/07/09 (Н1N1) pdm09, приводящего к появлению частиц с оболочками, потерявшими сферическую форму и имеющими продольные деформации (рис. 2б). Прочность связи в электростатическом комплексе определяется его зарядом и размерами. Поскольку гетерополианионы, содержащие атомы ванадия, обладают более высоким зарядом по сравнению с гетерополианионами, не имеющими эти атомы, то, возможно, этим объясняется их высокая эффективность при деструкции оболочки вирусов. Формирующиеся в результате действия гетерополианионов поры являются каналами проникновения протонов через липидную оболочку, что должно приводить к разрушению матрикса М1. Вероятно, эти механизмы ответственны за значительные разрушения оболочки вирусов гриппа A/California/07/09 (Н1N1) pdm09 в результате действия гетерополикислоты H5PMo10V2O4 (рис. 2б).
Одним из сложных вопросов, требующих дальнейших исследований, является вопрос об избирательности действия ГПК на различные штаммы вирусов [9, 12, 14]. Известно, что ПОМ могут ингибировать различные стадии инфицирования клеток. Механизм их действия изучен недостаточно. По результатам данной работы можно предположить, что при достаточно больших концентрациях ГПК (100 мкм и выше) будет осуществляться травление оболочек вирусов, скорость которого будет определяться различиями в липидном составе оболочек различных штаммов гриппа, даже культивированных в одних условиях. То, что такие различия могут быть значительными, показано в работе [35], на примере трех различных штаммов вирусов гриппа, культивированных в одинаковых условиях на куриных эмбрионах, но имеющих заметно различающиеся содержания основных фосфолипидов, таких как фосфатидилэтаноламин и фосфатилдихолин. Скорость травления этих оболочек гетерополианионами должна, соответственно, различаться и зависеть также от физико-химических свойств гетерополианионов.
Таким образом, мы полагаем, что при физиологических значениях рН деструктивное действие ГПК на оболочку вирусов осуществляется за счет деструкции липидного слоя в результате воздействия гетерополианионов. Это вызывает образование пор и деструкцию матрикса М1 при проникновении протонов сквозь липидную оболочку. В случае достаточно высокой кислотности среды следует ожидать проникновения протонов внутрь вируса не только сквозь образовавшиеся поры, но и через канал М2.
ЗАКЛЮЧЕНИЕ
Результаты электронно-микроскопических исследований деструкции оболочки вирусов гриппа, приведенные в данной работе, дают основания предположить, что стабильность оболочки к действию гетерополианионов со структурой Кеггина определяется распределением и содержанием холестерина и структурой матриксного белка М1. Предложен механизм деструкции оболочки, основанный на экстракции связующих липидов, таких как холестерин, и травлении липидной оболочки, приводящем к образованию пор. Через эти поры протоны могут проникать в вирусы и растворять матрикс М1.
Работа выполнена в рамках государственного задания по теме “Фундаментальные основы создания наноструктурированных систем нового поколения с уникальными эксплуатационными электрическими и магнитными свойствами” № 0082-2018-0003 (регистрационный номер АААА-А18-118012390045-2) и при финансовой поддержке Российским фондом фундаментальных исследований (грант № 18-54-00004 Бел_а) и Белорусского фонда фундаментальных исследований (договор № Х18Р-110).
Список литературы
Pope M.T. Heteropoly and Isopoly Oxometalates. Berlin: Springer-Verlag, 1983.
Далидчик Ф.И., Балашов Е.М., Буданов Б.А. и др. // Хим. физика. 2010. Т. 29. № 11. С. 21.
Rhule J.T., Hill C.L., Judd D.A. et al. // Chem. Rev. 1998. V. 98. P. 327.
Bijelic A., Aureliano M., Rompel A. // Angew. Chem. Intern. Ed. Engl. 2019. V. 58. № 10. P. 2980.
Shigeta S. // Antivir. Chem. Chemother. 1998. V. 9. № 2. P. 93.
Ikeda S., Neyts J., Yamamoto N. et al. // Antivir. Chem. Chemother. 1993. V. 4. № 5. P. 253.
Shigeta S., Mori S., Watanabe J. et al. // Ibid. 1995. V. 6. № 2. P. 114.
Shigeta S., Mori S., Watanabe J. et al. // Ibid. 1996. V. 7. № 6. P. 346.
Shigeta S., Mori S., Yamase T. et al. // Biomed. Pharmacother. 2006. V. 60. № 5. P. 211.
Васильев Р.Ф., Кънчева В.Д., Наумов В.В. и др. // Хим. физика. 2020. Т. 39. № 6. С. 36.
Qi Y., Xiang Y., Wang J. et al. // Antivir. Res. 2013. V. 100. № 2. P. 392.
Лопатина О.А., Исаева Е.А., Суетина И.А. и др. // Наноматериалы и наноструктуры. 2016. Т. 7. № 1. С. 36.
Лопатина О.А., Суетина И.А., Мезенцева М.В. и др. // Хим. физика. 2020. Т. 39. № 1. С. 52.
Shigeta S., Mori S., Kodama E. et al. // Antivir. Res. 2003. V. 58. P. 265.
Yamase T. // J. Mater. Chem. 2005. V. 15. P. 4773.
Kates M., Allison A.C., Tyrell D.A.J. et al. // Cold Spring Harb Symp. Quant. Biol. 1962. V. 27. P. 293.
Gerl M.-J., Sampaio J. L., Urban S. et al. // J. Cell Biol. 2012. V. 196. № 2. P. 213.
Hu B., Hofer C.T., Thiele C. et al. // J. Virol. 2019. V. 93. № 15. P. e00555-19.
Verma D.K., Gupta D., Kumar L.S. // Viruses. 2018. V. 10. № 11. P. 650.
Barman S., Nayak D.P. // J. Virol. 2007. V. 81. № 22. P. 12169.
Takahashi T., Suzuki T. // Biochem. Res. Intern. 2011. V. 2011. Article ID 245090. P. 1.
Sun X., Whittaker G.R. // J. Virol. 2003. V. 77. № 23. P. 12543.
Li S., Eghiaian F., Sieben C. et al. // Biophys. J. 2011. V. 100. № 3. P. 637.
Sha B., Luo M. // Nat. Struct. Biol. 1997. V. 4. P. 239.
Harris A., Cardone G., Winkler D.C. et al. // Proc. Natl Acad. Sci. USA. 2006. V. 103. № 50. P. 19123.
Fontana J., Cardone G., Heymann J.B. et al. // J. Virol. 2012. V. 86. № 6. P. 2919.
Fontana J., Steven A.C. // J. Virol. 2013. V. 87. № 10. P. 5621.
Chen Q., Huang X., Wei R. et al. // Biochem. Biophys. Res. Commun. 2019. V. 516. № 1. P. 57.
Barman S., Nayak D.P. // J. Virol. 2012. V. 81. № 2. P. 12169.
Graham D.R.M., Chertova E., Hilburn J.M. et al. // J. Virol. 2003. V. 77. № 15. P. 8237.
Ковалевский С.А., Гулин А.А., Лопатина О.А. и др. // Рос. нанотехнологии. 2019. Т. 14. № 9–10. С. 77.
Kobayashi D., Ouchi Y., Sadakane M. et al. // Chem. Lett. 2017. V. 46. P. 533.
Nabika H., Inomata Y., Itoh E. et al. // RSC Adv. 2013. V. 3. P. 21271.
Nabika H., Sakamoto A., Tero R. et al. // J. Phys. Chem. C. 2016. V. 120. P. 15640.
Ivanova1 P.T., Myers D.S., Milne S.B. et al. // ACS Infect. Dis. 2015. V. 11. № 1(9). P. 399.
Дополнительные материалы отсутствуют.
Инструменты
Химическая физика