

Лесоведение, 2019, № 1, стр. 7-18
Формирование пространственной структуры сосновых древостоев при рубках ухода
А. Н. Борисов a, *, В. В. Иванов a, А. Е. Петренко a
a Институт леса им. В.Н. Сукачева СО РАН – обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28,
Россия
* E-mail: alnik_borisov@mail.ru
Поступила в редакцию 11.01.2018
После доработки 02.04.2018
Принята к публикации 08.10.2018
Аннотация
Рассматривается пространственная структура древостоя и распределение доступного ресурса между деревьями с точки зрения получения максимальной продуктивности древостоя и обеспечения характеристик, соответствующих его целевому назначению. Анализ пространственного распределения ресурса между конкурирующими за него деревьями основан на использовании областей доминирования (ОД). Объектом исследований служили молодняки, средневозрастные и спелые, чистые по составу сосновые насаждения экспериментального хозяйства Института леса СО РАН “Погорельский бор”, территориально входящего в Красноярскую островную лесостепь. Проведенный анализ позволяет сделать вывод, что оптимальной с точки зрения прироста стволовой древесины можно считать площадь ОД, составляющую 30–40% от площади свободного роста. Деревья, у которых площадь ОД близка к указанной величине, эффективно используют доступный ресурс, имеют хорошо развитую крону и показывают стабильный рост. Показано, что для всех возрастных этапов развития древостоя увеличение густоты сверх рассчитанной оптимальной приводит к снижению прироста по объему ствола дерева. Проведенные исследования позволяют определить параметры целевых насаждений при интенсивном лесовыращивании и формировании структуры древостоев, обеспечивающей выполнение как хозяйственных, так и природоохранных функций лесов.
В процессе роста древостоя происходит его изреживание. Об интенсивности этого процесса говорит тот факт, что к возрасту рубки в древостое остается 3–5% от начальной численности деревьев. В связи с этим встает вопрос о том, как посредством лесохозяйственных мероприятий за счет уменьшения конкуренции со стороны ослабленных и неперспективных особей и перераспределения доступного ресурса получить деревья более высокого качества и сократить сроки лесовыращивания. Практические рекомендации, направленные на интенсификацию лесовыращивания, носят слишком общий характер и требуют уточнения с учетом лесорастительных условий и ценотической структуры насаждений. Поэтому необходима разработка научно обоснованных лесоводственных приемов и нормативов, позволяющих формировать высокопродуктивные насаждения разного целевого назначения в соответствующих лесорастительных условиях.
Изучение естественных процессов роста и анализ реакции деревьев и древостоев на разреживание и другие воздействия является важной задачей. Многие лесоводы считают, что максимальный текущий прирост древостоев наблюдается при полноте ниже 1.0 и для любого насаждения существуют оптимальные полнота и густота (Assmann, 1961; Поляков, 1973; Малышев и др., 2010; Антанайтис и др., 1986; Кузьмичев, 2013). Вопросы оптимизации структуры древостоев должны решаться с учетом лесорастительного районирования и типологической дифференциации, но основным критерием является их целевое назначение. В работе (Вайс, 2005) под оптимальной структурой древостоя понимается размещение деревьев, которое обеспечивает их максимальный прирост по объему и устойчивость в целом для насаждения. В лесах эксплуатационного назначения одним из ведущих принципов назначения рубок ухода считается максимальное приближение полноты и запаса древостоя к параметрам нормального леса к возрасту главной рубки (Нагимов, 2000). В зависимости от выбранного критерия оптимальности структура древостоя на каждом возрастном этапе формирования может существенно меняться. В исследованиях по оптимизации густоты применяются разные методы анализа данных, тем не менее при использовании текущего прироста деревьев по объему и площади сечения обеспечиваются близкие результаты (Чернов и др., 2012). При изучении структуры древостоев часто используется моделирование процессов роста древостоев (Разин, 1981; Антанайтис и др., 1986; Grabarnik, Sarkka, 2009; Кузьмичев, 2013). В полученных моделях учитываются закономерности изменения прироста по диаметру, высоте, запасу в зависимости от условий местопроизрастания, возраста и густоты древостоев. При разработке таблиц оптимальной густоты древостоев нередко использовались те или иные оценки площадей роста (Поляков, 1973; Разин, 1981; Вайс, 2005; Нагимов, 2000). Рост отдельных деревьев и древостоя в целом зависит от того, насколько эффективно используется ресурс, доступный на данном участке. Хорошо известна тесная зависимость характеристик дерева от площади роста, которую рассматривают как оценку доступного для данного дерева ресурса (Поляков, 1973; Тябера, 1982; Вайс, 2005; Чернов и др., 2012; Борисов и др., 2014а; Рогозин, Разин, 2015).
В связи с этим особую важность приобретает вопрос формирования пространственной структуры древостоев, обеспечивающей эффективное распределение ресурса между деревьями и получение максимальной продуктивности. Рассмотрению именно этих аспектов посвящена настоящая работа.
ОБЪЕКТЫ И МЕТОДИКА
При изучении роста деревьев в древостое важно знать, как происходит распределение доступного ресурса между конкурирующими за него деревьями. Для этой цели обычно используют площадь роста. Однако, в разных работах в это понятие вкладывается различный смысл, и способы оценки этой характеристики значительно разнятся. Наиболее широко применяется построение для каждого дерева многоугольников, образуемых прямыми, которые проводятся перпендикулярно линиям, соединяющим это дерево с его ближайшими соседями. В случае, когда эти перпендикуляры проводятся по середине расстояния между соседями, получают полигоны, носящие имя Вороного. Полигоны Вороного позволяют провести анализ горизонтальной структуры древостоя, сделать выводы о том или ином виде размещения деревьев на занимаемой ими территории (Brown, 1965). Однако подобный подход не отражает напряженности конкурентных отношений и распределения ресурса между особями, поскольку не учитывает размерных характеристик деревьев. В методе Штера (Нагимов, 2000) стороны полигонов смещаются от центрального дерева в направлении произрастания более тонкого соседа пропорционально диаметрам их стволов. В этом случае на полигоны разбивается весь рассматриваемый участок, даже низкополнотные насаждения и участки с прогалинами. Считать такие полигоны площадью, на которой растет дерево, и применять в этом случае термин “площадь роста”, на наш взгляд, некорректно. Некоторые исследователи считают площадью роста размер проекции кроны дерева (Изюмский, 1968), не учитывая, что корневая система, обеспечивающая доступность почвенных ресурсов, по площади существенно превышает проекцию кроны. Ситуацию не меняет и использование комбинированных методов разбиения на полигоны с учетом как диаметров стволов, так и размеров крон (Кузьмичев, 2013). Использование термина “площадь питания”, по крайней мере, имеет понятный биологический смысл, если иметь в виду пространственные характеристики корневой системы, обеспечивающей дерево влагой и минеральными веществами. Но и в этом случае разбиение территории на площади питания для каждого дерева наталкивается на ряд сложностей. Прежде всего, определение архитектоники корневой системы дерева очень трудоемко и на практике производится для единичных деревьев. Следует учитывать, что протяженность корней, например, в сосновых древостоях в возрасте 30 лет достигает 8–10 м, и коэффициент взаимоперекрытия проекций корневых систем деревьев в зависимости от густоты древостоя составляет 6 и более (Рахтеенко, 1952). Поэтому проблема оценки вклада пространственно перекрывающихся корневых систем в использование ресурса в области их перекрытия остается нерешенной. Суммарная же площадь питания будет многократно превышать размер участка, занимаемого деревьями, и использование площади питания в качестве оценки распределения между особями ресурса, который имеется на данном участке, будет некорректным.
В данной работе применен анализ пространственного распределения ресурса между конкурирующими за него деревьями, основанный на использовании областей доминирования (Борисов, 2013). Этот метод позволяет получить экологически обоснованные оценки распределения ресурса между конкурирующими за него особями. Под ресурсами здесь мы понимаем те факторы, образующие экологическую нишу вида, которые обеспечивают его жизнедеятельность. В предложенном подходе областью доминирования (ОД) является область, в которой особь оказывает доминирующее влияние в пространственном освоении ресурса в предположении, что это влияние прямо пропорционально размеру особи и обратно пропорционально квадрату расстояния от нее. Степень этого влияния оценивается как:
(1)
$\left\{ \begin{gathered} E = \frac{k}{{P(x,y)}}\frac{M}{{({{{(x - {{x}_{0}})}}^{2}} + {{{(y - {{y}_{0}})}}^{2}})}},\,\,\,\,E \geqslant {{E}_{0}} \hfill \\ E = 0,\,\,\,\,E < {{E}_{0}}, \hfill \\ \end{gathered} \right.$Объектом исследований служили молодняки, средневозрастные и спелые, чистые по составу сосновые насаждения экспериментального хозяйства Института леса СО РАН “Погорельский бор”, территориально входящего в Красноярскую островную лесостепь. Закладка пробных площадей была выполнена с учетом теоретических положений таксации согласно ОСТ 56-69-83 “Пробные площади лесоустроительные” (1983). Таксационные характеристики пробных площадей приведены в табл. 1.
Таблица 1.
Основные таксационные показатели древостоев на пробных площадях
| № пр. пл. | Возраст, лет | Состояние | Густота, шт. га–1 | Средний диаметр, см | Средняя высота, м | Сумма площадей сечения, м2 га–1 | Полнота | Запас, м3 га–1 |
Интенсивность рубки по запасу, % |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 8 | После рубки | 2900 | 3.5 | 2.4 | 2.8 | – | 5.1 | 78 |
| 13 | 2900 | 8.0 | 6.2 | 14.7 | – | 51.6 | |||
| 2 | 8 | 9500 | 2.9 | 2.5 | 6.3 | – | 12.2 | 46 | |
| 13 | 9450 | 5.4 | 5.4 | 21.7 | – | 73.4 | |||
| 3 | 8 | 17 480 | 2.5 | 2.3 | 8.2 | – | 15.8 | 29 | |
| 13 | 16 830 | 4.2 | 5.6 | 22.9 | – | 83.2 | |||
| 4 | 8 | Контроль | 40 667 | 1.9 | 2.1 | 11.2 | – | 22.4 | 0 |
| 13 | 34 200 | 3.3 | 5.5 | 28.9 | – | 109.3 | |||
| 5 | 37 | До рубки | 5460 | 10.5 | 15.7 | 47.5 | 1.5 | 355 | 13 |
| После рубки | 2727 | 13.2 | 17.6 | 37.5 | 1.1 | 308 | |||
| 47 | – | 2420 | 15.8 | 19.0 | 47.4 | 1.4 | 415 | ||
| 51 | После рубки | 1280 | 18.0 | 19.7 | 32.5 | 1.0 | 293 | 29 | |
| 6 | 37 | До рубки | 4020 | 12.4 | 17.1 | 48.2 | 1.5 | 388 | 17 |
| После рубки | 2280 | 14.9 | 18.5 | 39.8 | 1.2 | 321 | |||
| 47 | – | 2073 | 17.1 | 19.6 | 50.5 | 1.5 | 456 | ||
| 7 | 37 | До рубки | 5593 | 10.8 | 15.9 | 50.9 | 1.6 | 383 | 16 |
| После рубки | 2487 | 14.0 | 18.1 | 38.5 | 1.2 | 323 | |||
| 47 | – | 2367 | 16.2 | 19.2 | 49.0 | 1.4 | 431 | ||
| 8 | 47 | Контроль | 2640 | 15.4 | 18.4 | 48.9 | 1.3 | 433 | 0 |
| 9 | 105 | До рубки | 474 | 33.0 | 23.6 | 40.5 | 1.1 | 424 | 30 |
| После рубки | 303 | 34.7 | 23.4 | 28.6 | 0.8 | 298 | |||
| 110 | – | 289 | 36.2 | 23.7 | 29.7 | 0.8 | 313 | ||
| 10 | 80 | После рубки | 462 | 26.8 | 22.4 | 26.0 | 0.7 | 261 | 35 |
| 110 | – | 444 | 35.4 | 23.8 | 43.7 | 1.2 | 462 | ||
| 11 | 107 | Контроль | 863 | 29.2 | 23.7 | 57.6 | 1.5 | 576 | 0 |
| 112 | 767 | 31.0 | 23.8 | 57.9 | 1.6 | 614 |
Молодняки произрастают на бывших сельскохозяйственных угодьях в Красноярской лесостепи. Благодаря хорошему осеменению эти участки достаточно быстро в течение 2-х лет были заселены сосной, и сформировались одновозрастные загущенные молодняки. В самих насаждениях, пройденных рубками ухода разной интенсивности с выборкой деревьев по числу стволов от 93 до 57%, заложены пробные площади размером 20 × × 20 м (Иванов и др., 2017). На контрольном участке размер пробной площади составил 20 × 15 м.
Средневозрастное сосновое насаждение, чистое по составу, сформировано на вырубке 1965 г. В лесотаксационном выделе сосняка зеленомошного I класса бонитета заложено три пробные площади размером 0.15 га с числом деревьев на каждой из них не менее 300, что обеспечило репрезентативность для исследуемого лесного участка. Все пробные площади отделялись друг от друга буферной зоной шириной 5 м. На пробных площадях выполнено картирование деревьев с разбивкой площади на квадраты размером 5 × 5 м, проведены сплошной перечет и нумерация деревьев. В качестве контрольной использовалась смежная территория в том же выделе. В 37 лет насаждения на пробных площадях обладали высокой полнотой и густотой и характеризовались значительной дифференциацией деревьев по диаметру. Наибольшее (до 57.3%) число деревьев было представлено диаметрами стволов на высоте 1.3 м до 10 см, с явными признаками угнетения, что свидетельствует о высокой напряженности конкурентных отношений.
Спелые сосновые насаждения, в которых проводились исследования, относятся к разнотравно-зеленомошной группе типов леса. Пробные площади закладывались на репрезентативных участках с разными вариантами выборочных рубок и в насаждении без рубки (контроль). Возраст древостоев на пробных площадях до рубки составлял 105–112 лет при относительной полноте 0.8–1.6 и стволовом запасе древесины 424–614 м3. Количество сухостоя не превышало 8–12% от общего числа стволов.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Величина доступного ресурса во многом определяет рост дерева в высоту, формирование кроны и прирост по диаметру ствола. Использование областей доминирования позволяет оценить, какую величину ресурса в том или ином возрасте должны иметь деревья для наиболее полной реализации потенциала роста. Анализ радиальных приростов и приростов по запасу в древостоях различной густоты направлен на определение площади ОД (SОД), позволяющей обеспечить высокие темпы роста деревьев на разных возрастных этапах. В первую очередь анализируются условия роста деревьев 2–3-го классов Крафта как наиболее эффективно использующих свои площади питания для увеличения прироста (Нагимов, 2000). Это согласуется также с “Правилами ухода за лесами” (2017), где к категории лучших относятся в первую очередь деревья 2, а также 1 и 3 классов Крафта.
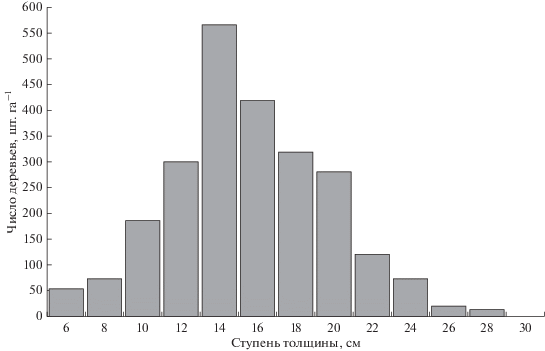
В средневозрастных насаждениях в возрасте 37 лет в пределах таксационного выдела проведено разреживание древостоев с интенсивностью по числу деревьев 42–56% и 13–17% по запасу. Сплошной перечет, проведенный на заложенных в этом выделе постоянных пробных площадях, показал, что через 10 лет имеется явно выраженная дифференциация со значительной долей отставших в росте и угнетенных деревьев, что говорит о недостаточной интенсивности проведенного разреживания. Сухостой составил около 5% от общего числа деревьев и представлен диаметрами от 4.6 до 12.6 см при среднем значении 6.9 см. Более 58% деревьев в возрасте 47 лет имели диаметр ствола менее величины среднего диаметра древостоя, который был равен 16 см (рис. 1). Вследствие высокой полноты древостоя формировались кроны с небольшой протяженностью. У деревьев среднего диаметра соотношение протяженности кроны по отношению к высоте дерева составило около 30%.
На основе анализа радиального прироста по кернам выделено 5 групп деревьев, имеющих разную динамику рангового положения. В группу “А” вошли деревья со стабильно низким рангом, в группу “Б” – уменьшающие ранг в процессе роста; группу “В” составляют деревья со стабильным средним ранговым положением; в группу “Г” вошли деревья, увеличивающие ранг, а в группу “Д” – со стабильно высоким рангом (Борисов и др., 2014б). Деревья групп “Г” и “Д” имеют хорошо развитую крону и показывают стабильный рост в высоту и по объему. Именно такие деревья, испытывающие некоторый дефицит ресурсов, но обладающие устойчивыми темпами роста, Г.Л. Кравченко (1972) относит к II–III типам онтогенеза. По классификации Крафта они относятся ко 2–3-му классам. Радиальный прирост в этой группе сопоставим и в некоторых случаях превышает прирост у деревьев высших рангов. Диаметр ствола отражает кумулятивный эффект роста дерева в зависимости от доступного для его роста ресурса. До 37-летнего возраста на пробных площадях 5–8 не проводилось рубок, и хотя в результате естественного отпада происходило изменение горизонтальной структуры древостоя и некоторое перераспределение ресурса, проведенный статистический анализ показал для насаждений в 37-летнем возрасте тесную зависимость диаметра ствола деревьев на высоте 1.3 м от SОД:
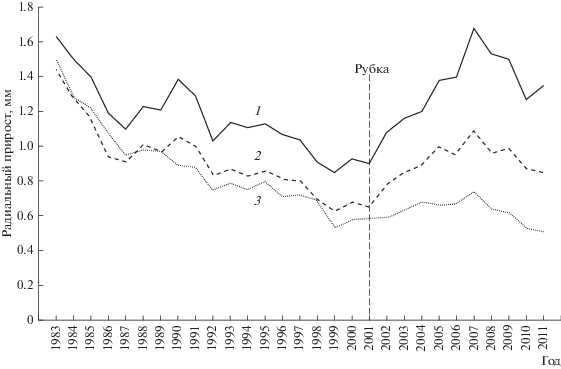
Анализ динамики радиального прироста выявил, что деревья групп “В” и “Г” после рубки показали наиболее выраженную реакцию на проведенную рубку 2001 г, и их годичный радиальный прирост увеличился до двух раз (рис. 2). Обеспеченность ресурсами в указанных группах по величине SОД составила 5 м2 и более. Для деревьев с 5 < SОД < 2 м2 прирост возрос в 1.7 раза, а в группе, где SОД < 2 м2 – не более чем в 1.3 раза.
Рис. 2.
Динамика годичного радиального прироста деревьев в зависимости от SОД. 1 – SОД > 5 м2, 2 – 5 м2 > SОД > 2 м2, 3 – SОД < 2 м2.
В средневозрастных и спелых древостоях выполнялось картирование и изучалось размещение деревьев на пробных площадях. Изменения в горизонтальной структуре и величинах ОД в результате рубок в возрасте 37 и 51 года показаны на рис. 3, a–б и 3, в–г, соответственно. В возрасте 51 года в рубку назначались деревья, имеющие низкие значения площади ОД. Интенсивность этой рубки по числу стволов составила 47%, а по запасу 29%. В табл. 2 для пробной площади 5 приведены значения средней площади ОД (Sср), стандартного отклонения для Sср (σ) и относительного отклонения σотн = σ/Sср. Величина Sср к 51-летнему возрасту после рубки выросла с 1.65 до 6.74 м2, а σотн уменьшилась с 0.96 до 0.28. Это говорит о том, что рубки привели к перераспределению ресурсов между оставшимися деревьями, размещение на исследуемом участке стало более равномерным. В период с 37 до 47-летнего возраста несколько выросли значения Sср и σотн, что объясняется увеличением дифференциации деревьев по диаметру, вызываемой значительной густотой и, как следствие, высокой напряженностью конкурентных отношений.
Рис. 3.
Размещение деревьев и их области доминирования: а – 37 лет, до рубки; б – 37 лет, после рубки; в – 51 год, до рубки; г – 51 год, после рубки.
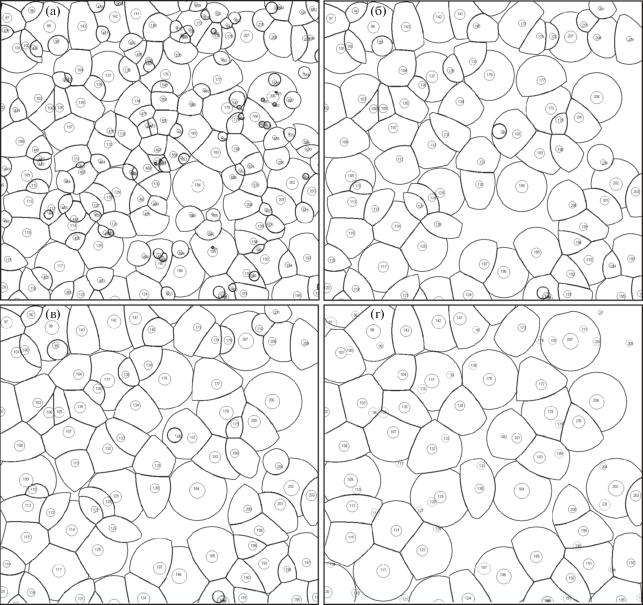
Таблица 2.
Характеристики площади ОД для пробной площади 5
| Показатель | 37 лет, до рубки | 37 лет, после рубки | 47 лет | 51 год, после рубки |
|---|---|---|---|---|
| Густота, шт га–1 | 5460 | 2727 | 2420 | 1280 |
| Средняя площадь ОД, м2 | 1.65 | 3.34 | 3.82 | 6.74 |
| Стандартное отклонение ОД, м2 | 1.59 | 1.87 | 2.21 | 1.92 |
| Относительное отклонение | 0.96 | 0.56 | 0.58 | 0.28 |
В спелых сосновых насаждениях изучены реакции деревьев и древостоя после разреживания. В пределах одного таксационного выдела в 1986 г проведена проходная рубка ухода за лесом интенсивностью 35% и в 2011 г – несплошная рубка интенсивностью 30% по запасу (табл. 1).
Оптимальная сумма площадей сечения, а, следовательно, и полнота древостоев зависят от климатических и почвенно-гидрологических условий, возраста насаждений. Текущий прирост по запасу и общая производительность древостоев максимальных значений достигают при их оптимальной сумме площадей сечений (Assmann, 1961). Анализ материалов, полученных при изучении сосновых молодняков, средневозрастных и спелых насаждений, позволяет сделать вывод, что оптимальной с точки зрения прироста стволовой древесины можно считать площадь, составляющую ОД 30–40% от площади свободного роста. Такой оптимум обеспечивает компромисс между увеличением прироста по объему ствола за счет роста доступного ресурса и увеличением запаса древостоя вследствие большей густоты. Деревья, у которых SОД близка к указанной величине, эффективно используют доступный ресурс, имеют хорошо развитую крону и показывают стабильный рост в высоту и по объему. Оптимальные значения площади ОД (Sопт) рассчитывались в зависимости от значений диаметра ствола на высоте 1.3 м и с учетом условий местообитания в соответствии с методикой расчета площади свободного роста, описанной в работе А.Н. Борисова с соавторами (2014а).
Используя значение Sопт можно рассчитать оптимальную густоту одновозрастного древостоя как Nопт = 10 000/Sопт шт. га–1. Предлагаемый подход позволяет определить желательную структуру сосняков на разных возрастных этапах их формирования с учетом лесорастительных условий. Величины, оптимальные для одновозрастных сосновых насаждений I класс бонитета, приведены в табл. 3. Диаметры ствола, использованные в табл. 3, соответствуют таблице хода роста для полных сосновых древостоев (Таблицы …, 2008). В насаждениях с аналогичными значениями среднего диаметра ствола при снижении густоты до рассчитанного Nопт будет обеспечен эффективный рост деревьев.
Таблица 3.
Характеристика сосновых насаждений I класс бонитета при оптимальной густоте
| Нормальный древостой | Оптимальные показатели | ||||
|---|---|---|---|---|---|
| возраст, лет | диаметр ствола, см | густота, шт. га–1 | Nопт, шт. га–1 | сумма площадей сечений, м2 га–1 | полнота |
| 20 | 6.3 | 6158 | 6061 | 18.9 | 0.99 |
| 30 | 10.6 | 3016 | 2778 | 24.5 | 0.92 |
| 40 | 14.9 | 1840 | 1600 | 27.9 | 0.87 |
| 50 | 19.0 | 1271 | 1081 | 30.6 | 0.85 |
| 60 | 22.7 | 953 | 784 | 31.7 | 0.82 |
| 70 | 26.1 | 757 | 604 | 32.3 | 0.79 |
| 80 | 29.1 | 628 | 490 | 32.6 | 0.78 |
| 90 | 31.7 | 538 | 413 | 32.6 | 0.77 |
| 100 | 34.0 | 474 | 358 | 32.5 | 0.75 |
| 110 | 36.0 | 426 | 317 | 32.3 | 0.74 |
| 120 | 37.7 | 390 | 287 | 32.0 | 0.73 |
Как видно из табл. 3, относительная полнота в средневозрастных сосняках I класса бонитета будет составлять 0.82–0.85, в спелых – 0.73–0.75, а сумма поперечных сечений стволов – 30.6–31.7 и 32.0–32.5 м2 га–1 соответственно. При снижении полноты средневозрастных и спелых насаждений соответственно до 0.71–0.75 и 0.65–0.67 будет обеспечиваться максимальное накопление фитомассы крон (Нагимов, 2000) С другой стороны, по данным Н.П. Гординой (1991) при снижении полноты до 0.58–0.61 в средневозрастных сосняках наблюдается максимальное накопление фитомассы крон, а в приспевающих, спелых и перестойных это происходит при полноте 0.60–0.63. С увеличением SОД прирост деревьев по объему вначале возрастает и достигает максимума при ее определенной величине. При дальнейшем возрастании SОД прирост по объему ствола остается на одном и том же уровне или даже снижается за счет разрастания крон. Такие тенденции при увеличении площадей роста отмечены и в работе В.В. Кузьмичева (2013).
Известно, что текущий прирост по запасу с возрастом закономерно уменьшается. Возраст дерева, на который приходится максимум прироста по диаметру, зависит от величины доступного ресурса на всем периоде его роста. Для деревьев II типа онтогенеза, которые при некотором дефиците ресурса характеризуются устойчивыми темпами роста, в возрасте 45 лет прирост по диаметру равен 0.39 см, а к 95 г. это значение составляет 0.22 см (Кравченко, 1972). Максимальная продуктивность древостоя в целом не означает максимальную продуктивность каждого дерева. В.В. Кузьмичев (2013) отмечал, что высокая продуктивность древостоя обеспечивается при пониженной продуктивности составляющих его особей, но более высокой численности. При повышенной густоте не только происходит ослабление древостоя в целом вследствие напряженных конкурентных отношений, но и не обеспечивается эффективный рост составляющих его деревьев, формируются разреженные кроны с малой их протяженностью, прирост стволовой древесины также характеризуется низкими значениями. В дальнейшем это приводит к интенсивному отпаду, снижению густоты и запаса.
В молодняках прирост по запасу на контрольной пробной площади выше всего. Однако это происходит только за счет того, что густота на этом участке в разы выше, чем на пробных площадях 1–3. А по величине среднего радиального прироста и среднего диаметра ствола на контрольном участке оказываются худшие показатели (табл. 1). За последние 5 лет густота на пробной площади 4 уже сократилась с 40 667 до 34 200 шт. га–1. В дальнейшем продуктивность деревьев в насаждении будет снижаться, поскольку из-за недостатка ресурсов и вследствие большой напряженности конкурентных отношений усилится их дифференциация. К 20-летнему возрасту по данным А.И. Бузыкина с соавторами (2008) в таких насаждениях густота уменьшается в 2 раза.
Анализ степени соответствия густоты оптимальным значениям был проведен по данным, полученным на экспериментальных участках для сосновых древостоев (табл. 1). В качестве меры отклонения от оптимума вычислялась величина относительного отклонения,
(3)
$dN = \frac{{({{N}_{{{\text{ф а к т }}}}} - {{N}_{{{\text{о п т }}}}})}}{{{{N}_{{{\text{о п т }}}}}}} \times 100\% ,$Рис. 4.
Средний прирост деревьев по объему ствола и отклонения густоты от оптимальной для одновозрастных сосновых древостоев. 1 – dM, 2 – dN. Индекс “к” для возраста древостоя на пробной площади обозначает контрольные участки.
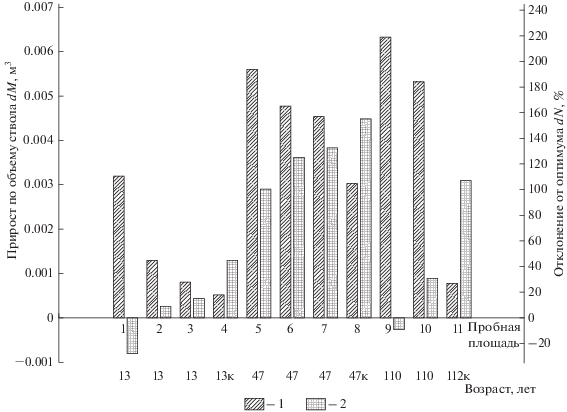
В то время, как в молодняках на пробных площадях 1–4 с уменьшением густоты прирост по запасу сокращается прирост по объему ствола dM растет (рис. 4). Из рис. 4 видно, что для средневозрастных сосняков интенсивность рубки, проведенной в 37-летнем возрасте, была недостаточной и эффект от нее проявился не в полной мере. Это подтверждается сплошным перечетом деревьев на пробных площадях 5–8. через 10 лет после рубки. В результате, как было сказано выше, это привело к явно выраженной дифференциации деревьев и наличию значительной доли отставших в росте и угнетенных особей. Существенный положительный эффект от рубки проявляется и в спелых насаждениях.
Заключение. В сосновых древостоях с высокой густотой при жесткой внутривидовой конкуренции велика вероятность потери общей производительности из-за несоответствия хода роста древостоев густотно-возрастным оптимумам продуктивности. Метод ОД позволяет определить, как распределяется в древостое доступный ресурс между конкурирующими за него деревьями. Показана тесная связь диаметров стволов деревьев на высоте 1.3 м с полученными с помощью этого метода значениями Sод. Проведенный анализ показал, что при Sод, составляющей 30–40% от площади свободного роста, деревья эффективно используют доступный ресурс, имеют хорошо развитую крону и устойчивый рост по диаметру и объему ствола. Такие значения Sод можно считать оптимумом, обеспечивающим компромисс между увеличением прироста по объему ствола за счет доступного ресурса и ростом запаса древостоя за счет большей густоты. Для рассмотренных в работе древостоев рассчитаны оптимальные величины густоты, которые могут служить ориентиром при проведении рубок ухода за лесом.
В случае свободного роста существенная часть биомассы приходится на крону, протяженность которой достигает 80–90% от высоты, повышается сучковатость ствола и снижаются темпы роста в высоту и по запасу древесины. При уменьшении площади ОД ниже Sопт формируется древостой, который может обладать большей густотой и высокой полнотой, но характеризующийся значительной дифференциацией по диаметрам и меньшим значением среднего диаметра. Оптимальные условия для роста необходимо обеспечивать не просто на момент рубки, а на весь период до следующего ее приема. Интенсивность рубки тогда следует делать соответственно большей. Необходимые оценки можно получить, применяя соответствующие модели роста древостоя.
Проведенные исследования позволяют в рамках предложенного подхода реализовать интенсивное лесовыращивание и формирование структуры древостоев, обеспечивающей выполнение как хозяйственных, так и природоохранных функций лесов. Эти данные могут быть использованы при разработке рекомендаций по проведению рубок ухода в сосновых насаждениях.
Список литературы
Антанайтис В.В., Тябера А.П., Шяпятене Я.А. Законы, закономерности роста и строения древостоев. Каунас: Литовская сельскохозяйственная академия, 1986. 157 с.
Борисов А.Н. Метод оценки распределения ресурса между деревьями в древостое // Состояние лесов и актуальные проблемы лесоуправления. // Матер. Всеросс. конф. с междунар. участием. (г. Хабаровск, 10–11 сентября 2013 г.). Хабаровск: Изд-во ФБУ “ДальНИИЛХ” С. 293–296.
Борисов А.Н., Иванов В.В., Екимов Е.В. Метод оценки пространственного распределения ресурса в экологической нише // Сибирский лесной журнал. 2014а. № 5. С. 113–121.
Борисов А.Н., Иванов В.В., Петренко А.Е. Оценка реакции соснового древостоя Красноярской лесостепи на рубку ухода // Лесоведение. 2014б. № 4. С. 22–27.
Бузыкин А.И., Пшеничникова Л.С., Собачкин Д.С., Собачкин Р.С. Естественное изреживание разногустотных молодняков в экспериментальных посадках сосны // Хвойные бореальной зоны. 2008. Т. 25. № 3–4. С. 244–249.
Вайс А.А. Оптимизация горизонтальной структуры смешанных разновозрастных пихтовых древостоев // Лесная таксация и лесоустройство. 2005. № 2. С. 17–21.
Гордина Н.П. Оптимизация структуры защитных лесов на основе биометрических показателей // Лесная таксация и лесоустройство: Межвуз. сб. науч. тр. Красноярск: СТИ, 1991. С. 52–59.
Иванов В.В., Борисов А.Н., Петренко А.Е., Семенякин Д.А., Собачкин Д.C., Собачкин Р.C. Густота сосновых древостоев при интенсивном лесовыращивании // Сибирский лесной журнал. 2017. № 6. С. 102–109.
Изюмский П.П. О методе рубок ухода за лесом // Лесное хозяйство. 1968. № 1. С. 23–26.
Кравченко Г.Л. Закономерности роста сосны. М.: Лесная пром-ть, 1972. 168 с.
Кузьмичев В.В. Закономерности динамики древостоев: принципы и модели. Новосибирск: Наука, 2013. 207 с.
Малышев В.В. Метод моделирования динамики хода роста и оптимизация густоты древостоев сосны // Генетика, селекция, семеноводство и воспроизведение древесных пород // Матер. Всеросс. конф. с междунар. участием. (г. Воронеж, 2010 г.). Воронеж: Воронежская государственная лесотехническая академия, 2010. С. 186–189.
Нагимов З.Я. Закономерности роста и формирования надземной фитомассы сосновых древостоев: Автореф. дис. … д-ра с.-х. наук 06.03.03. Екатеринбург: Уральская государственная лесотехническая. академия, 2000. 40 с.
ОСТ 56-69-83 Площади пробные лесоустроительные. Метод закладки. М.: ЦБНТИ Гослесхоза СССР, 1984. 60 с.
Полетаев И.А. О математических моделях элементарных процессов в биогеоценозах. Проблемы кибернетики. 1966. Вып. 16. С. 171–190.
Поляков А.К. Определение оптимальной густоты сосны в свежей субори // Лесное хозяйство. 1973. № 12. С. 14–18.
Правила ухода за лесами. Утверждены приказом Минприроды России от 22.11.2017 г. № 626 (Зарегистрирован в Минюсте РФ 22.12.2017 N 49 381).
Разин Г.С. Способ определения оптимальной текущей густоты древостоев при их целевом выращивании // Известия высших учебных заведений. Лесной журнал. 1981. № 3. С. 35–38.
Рахтеенко И.Н. Корневые системы древесных и кустарниковых пород. М.; Л.: Гослестехиздат, 1952. 107 с.
Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы. Raleigh, North Carolina, USA: Lulu Press, Inc, 2015. 200 с.
Швиденко А.З., Щепащенко Д.Г., Нильссон С., Булуй Ю.И. Таблицы и модели хода роста и продуктивности насаждений основных лесообразующих пород Северной Евразии (нормативно-справочные материалы). М.: Изд-во Рослесхоз, IIASA, 2008. 886 с.
Тябера А.П. Влияние площади роста на таксационные показатели и качество древесины сосняков Литовской ССР // Лесоведение. 1982. № 2. С. 78–84.
Чернов Н.Н., Соловьев В.М., Нагимов З.Я. Методические основы лесокультурных исследований. Екатеринбург: Уральская государственная лесотехническая академия, 2012. 421 с.
Assmann E. Waldertragskunde. Organische Produktion, Struktur, Zuwachs und Ertrag von Waldbeständen. München–Bonn–Wien: BLV Verlagsgessellschaft: 1961. 490 s.
Brown G. S. Point density in stems per acre. New Zealand Forestry Research Notes. 1965. № 38. P. 1–11.
Grabarnik P., Särkkä A. Modelling the spatial structure of forest stands by multivariate point prosesses with hierarchical interactions // Ecological Modelling, 2009, V. 220. № 9. P. 1232–1240.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение