

Лесоведение, 2019, № 4, стр. 274-285
Закономерности изменения рангового положения деревьев по их размерам в ценопопуляциях сосны обыкновенной
Ю. П. Демаков a, *, Т. В. Нуреева a
a Поволжский государственный технологический университет
424000 г. Йошкар-Ола, пл. Ленина, 3, Россия
* E-mail: DemakovYP@volgatech.net
Поступила в редакцию 25.12.2017
После доработки 16.04.2018
Принята к публикации 04.02.2019
Аннотация
Отражены результаты исследования по выявлению закономерностей изменения рангового положения деревьев по классам Крафта в ценопопуляциях сосны, проведенного на 125 пробных площадях в древостоях различного возраста (от 15 до 110 лет), происхождения, густоты (от 0.5 до 22 тыс. экз./га) и условий произрастания. Показано, что оно не остается стабильным в течение их жизни, изменяясь по мере роста в ту или иную сторону, однако из IV–V классов Крафта в основной полог древостоя со временем переходит более 20% особей, а из высших классов в низшие – всего 6%. Неустойчиво положение только деревьев III класса, половина из которых понижают со временем свой ранг. Одной из причин изменения ранга деревьев является разная скорость исчерпания ими жизненного потенциала, связанного с темпами роста диаметра ствола, которые у деревьев каждого класса Крафта различны. Установлено, что особи IV–V классов Крафта практически полностью исчерпывают свой жизненный потенциал уже к 40–50 гг., почти не увеличивая диаметр ствола, а остальных классов – не ранее 80–90 лет. Приведены математические уравнения, отражающие связь среднего диаметра деревьев каждого класса со средним диаметром древостоя, позволяющие оценить ранговую структуру и жизнеспособность ценопопуляций. Показано, что хорошим диагностическим признаком состояния жизнеспособности дерева и его рангового положения в ценопопуляции является отношение между высотой дерева и диаметром или площадью сечения ствола.
Общеизвестно, что деревья в любом одновозрастном древостое как естественного, так и искусственного происхождения сильно различаются между собой по высоте, диаметру и объему ствола, размерам и форме кроны (Сукачев, 1928; Нестеров, 1961; Белов, 1983). Структурное разнообразие древостоев является одним из наиболее интересных явлений, изучаемых лесными экологами и практиками (Szmyt, Korzeniewicz, 2014). Эта картина является хотя и застывшим, но необычайно ярким кадром динамического процесса дифференциации деревьев, возникающего в результате их борьбы за свое существование и действующего по законам естественного отбора, детерминированного наследственными свойствами особей и усложненного вариациями условий среды. Чем более структурирована ценопопуляция, тем выше возможности управления ее развитием, что особенно важно для искусственно создаваемых в Европе лесов (Pastorella, Paletto, 2013; Szmyt, Korzeniewicz, 2014).
Для оценки рангового положения деревьев в ценопопуляциях были разработаны разнообразные классификационные шкалы, первой и наиболее удачной среди которых являлась шкала немецкого лесовода Эдуарда-Фридриха-Густава Крафта, опубликовавшего свою работу в 1884 г. Хотя шкала, предложенная этим ученым, в определенной мере искусственна, т.к. в действительности между деревьями разных классов наблюдается плавный переход, однако она позволяет на практике все же довольно объективно оценить структуру древостоя и обосновать проведение необходимых лесохозяйственных мероприятий.
Шансы на дальнейшее существование у деревьев разного рангового положения далеко не одинаковы и анализ литературных источников свидетельствует о весьма большом разбросе величины вероятности их выживания, что связано со спецификой исходной структуры древостоев и последующего их развития. Так, многолетние исследования А.Д. Дударева (1956), проведенные им на постоянных пробных площадях в сосняках возрастом от 50 до 100 лет, показали, что за 5–7 лет только небольшая часть деревьев из II и III классов Крафта (по 8% в каждом) перешла в высшие, в основном же они снизили свой ранг. Среди деревьев I класса не изменили положения 67%, II класса – 48%, III – только 23%. Из числа деревьев II класса Крафта усохло 1.5% особей, III – 3.5%, IV – 16%, а V – уже 70%. Отпад среди деревьев I класса отсутствовал. Е.К. Барнишкис (1972), обобщив материалы многолетних наблюдений Н.С. Нестерова, В.Г. Нестерова и П.С. Кондратьева на постоянных пробных площадях в лесной опытной даче ТСХА, показал, что в сосняках с возраста от 45 до 80 лет полностью отпали деревья IV–V классов, убыль из III класса составила от 86.3 до 91.4%, II – 56%, а I – 35%. По данным наших многолетних наблюдений (Демаков, 2000), проведенных на постоянных пробных площадях в 60–80-летних загущенных сосняках лишайниково-мшистых и зеленомошных, за 20 лет отпали практически все деревья V класса Крафта (92–98%) и большая часть деревьев IV класса (52–71%). Из числа деревьев III класса погибло не более 9.5%, а II – 2.3%. Все деревья I класса остались живыми.
Вопрос о закономерностях дифференциации древостоев, несмотря на внешнюю его простоту и давнюю историю изучения, далек от окончательного решения. По-прежнему ощущается острый недостаток экспериментальных данных, полученных путем длительных регулярных наблюдений на постоянных пробных площадях или специальных опытных объектах. Крайне мало исследований влияния на характер рангового распределения деревьев, а также вероятности их выживаемости биоценотических и климатических факторов.
Для практических целей лесоводства важно оценить не только вероятность выживаемости деревьев того или иного класса развития, но также долю их участия в структуре древостоя в зависимости от его возраста, густоты и условий произрастания. Понимание и численное определение структурного разнообразия может помочь в моделировании роста и производительности древостоев, а практикам-лесоводам – в принятии важных решений по управлению лесами. Имеющийся дефицит данных о процессе дифференциации и выживаемости деревьев в лесных биогеоценозах и их противоречивость определил выбор темы нашего исследования.
Цель исследования заключалась в познании закономерностей и причин дифференциации особей в ценопопуляциях сосны, разработке математических моделей хода роста деревьев разных классов Крафта, определении скорости их жизненного потенциала, необходимых для решения практических задач по управлению структурой и производительностью древостоев.
ОБЪЕКТЫ И МЕТОДИКА
Исходным материалом для анализа служили данные, собранные на 125 постоянных и временных пробных площадях, заложенных в чистых сосновых древостоях разного возраста (от 15 до 110 лет), происхождения, густоты (от 0.5 до 22 тыс. экз. га–1) и условий произрастания. Каждая пробная площадь включала не менее 200 живых деревьев, разбитых при учете на ступени толщины и классы развития Крафта. Определение объема ствола и массы различных фракций деревьев проведено расчетным путем по разработанным нами математическим моделям (Демаков и др., 2015), реализованным в соответствующей компьютерной программе (Демаков и др., 2017). На ряде пробных площадей у модельных деревьев, отобранных из разных классов Крафта (по 10–15 деревьев в каждом), оценены величины годичного прироста ствола в высоту и толщину за все время их жизни. Радиальный прирост измеряли с помощью бинокулярного микроскопа на кернах, взятых возрастным буравом Пресслера. На одной из пробных площадей в 55-летних чистых культурах сосны Iа класса бонитета, созданных на светло-серой лесной суглинистой почве по схеме 1.5 × 1.5 м, проведено измерение радиального прироста по пятилетиям у всех оставшихся на ней живых 138 деревьев (текущая густота древостоя составляла 1115 экз. га–1, а сохранность – 25%). С 1981 г. на 10 постоянных пробных площадях с пронумерованными деревьями периодически измеряли длину окружности ствола и оценивали состояние их жизнеспособности. Статистическую обработку материала провели на ПК с использованием стандартных методов и средств.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ собранного материала показал, что дифференциация деревьев по их размерам начинается в ценопопуляциях очень рано и продолжается до глубокой старости. В большинстве древостоев преобладают деревья среднего рангового положения (табл. 1). Доля господствующих и сверхгосподствующих деревьев, которые по своим параметрам значительно превосходят остальные (табл. 2), по числу стволов невелика, однако по площади их сечения и массе хвои, а значит и роли в функционировании биогеоценозов составляет в среднем около 50%, достигая в ряде случаев даже 80%. Доля деревьев IV–V классов Г. Крафта, не имеющих особых перспектив развития, составляет в среднем по числу стволов 31.6%, а по площади их сечения и массе хвои всего лишь 12.3–15.9%.
Таблица 1.
Доля деревьев сосны разных классов Крафта в сложении структуры ценопопуляций, %
| Параметр структуры | I | II | III | IV | V |
|---|---|---|---|---|---|
| Число деревьев | $\frac{{8.4 \pm 0.5}}{{0.5--33.9}}$ | $\frac{{19.9 \pm 0.6}}{{5.7--39.6}}$ | $\frac{{40.2 \pm 0.9}}{{21.2--77.3}}$ | $\frac{{22.5 \pm 0.7}}{{1.1--42.0}}$ | $\frac{{9.1 \pm 0.5}}{{0.0--24.1}}$ |
| Площадь сечения стволов | $\frac{{18.8 \pm 0.9}}{{1.7--47.7}}$ | $\frac{{30.7 \pm 0.6}}{{9.9--47.7}}$ | $\frac{{38.3 \pm 1.0}}{{12.4--76.0}}$ | $\frac{{10.7 \pm 0.5}}{{0.7--33.4}}$ | $\frac{{1.6 \pm 0.1}}{{0.0--6.5}}$ |
| Масса хвои деревьев | $\frac{{16.1 \pm 0.8}}{{1.4--43.9}}$ | $\frac{{28.4 \pm 0.6}}{{9.1--44.2}}$ | $\frac{{39.6 \pm 1.0}}{{14.2--77.6}}$ | $\frac{{13.2 \pm 0.6}}{{0.7--35.1}}$ | $\frac{{2.7 \pm 0.2}}{{0.0--8.2}}$ |
Таблица 2.
Влияние возраста древостоя на величину долевого участия деревьев разных рангов
| Возраст древостоя, лет | Число участков |
Доля деревьев разных классов Г. Крафта по массе их хвои в ценопопуляции, % | ||||
|---|---|---|---|---|---|---|
| I | II | III | IV | V | ||
| 14–15 | 28 | 23.0 (6.8–32.8) | 26.3 (11.9–37.1) | 39.9 (23.8–67.9) | 8.5 (2.9–19.3) | 2.3 (0.2–7.8) |
| 16–18 | 49 | 11.1 (1.4–22.0) | 30.3 (19.3–44.2) | 41.9 (26.4–65.7) | 14.3 (6.0–26.6) | 2.4 (0.4–6.5) |
| 21–23 | 11 | 23.7 (8.9–43.9) | 26.8 (15.2–33.7) | 31.8 (14.2–47.5) | 14.9 (7.4–22.7) | 2.8 (0.9–4.6) |
| 28–31 | 5 | 17.0 (4.1–35.3) | 20.1 (18.8–26.2) | 34.0 (31.8–35.7) | 23.4 (8.7–35.1) | 5.5 (1.7–7.4) |
| 45–55 | 15 | 14.0 (7.1–26.4) | 26.4 (9.1–39.7) | 42.9 (29.3–76.6) | 14.2 (6.2–27.4) | 2.5 (0.0–8.2) |
| 70–100 | 8 | 13.4 (2.8–43.9) | 37.0 (26.9–43.3) | 33.1 (19.5–45.6) | 13.2 (0.7–24.1) | 3.4 (0.0–5.9) |
Долевое участие деревьев разных классов и их относительные размеры, как свидетельствуют приведенные данные, очень сильно варьируют в разрезе изученных ценопопуляций, что связано как с особенностями их генотипической структуры (Романовский, 1994; Санников, Петрова, 2003; Романовский, Щекалев, 2014), так и с субъективными ошибками глазомерной оценки, избежать которых невозможно. Влияние возраста древостоев и их густоты, изменяющихся в очень больших пределах, на величину показателей при этом практически не проявляется (η < 0.25 при р < 0.05), что полностью подтверждает известный закон единства строения древостоев. Только при крайне низкой густоте лесных культур (от 1.0 тыс. шт. га–1 и менее) резко увеличивается доля деревьев III класса Крафта, что указывает на снижение интенсивности процесса дифференциации особей. С возрастом древостоев (А, лет) довольно четко изменяется, как показали расчеты, только величина среднего диаметра деревьев разных классов (dk, см), что с высокой точностью (p < 0.001) описывает функция Митчерлиха dk = К[1 – exp(–a × 10–3A)]b, параметры которой представлены в табл. 3. Установлено, что средний диаметр деревьев каждого класса очень тесно связан со средним диаметром всего древостоя (D, см), что аппроксимирует степенное уравнение dk = aDb, опираясь на которое можно провести оценку ранговой структуры ценопопуляций не только по материалам фактического учета, но также таблицам хода роста насаждений и данным таксационных описаний.
Таблица 3.
Значения параметров функций зависимости среднего диаметра деревьев разных рангов от возраста и среднего диаметра древостоя в сосняках лишайниковых и зеленомошных
| Параметр функции |
Значения параметров функций для деревьев разных классов Г. Крафта | ||||
|---|---|---|---|---|---|
| I | II | III | IV | V | |
| dk = К[1 – exp(–a × 10–3A)]b | |||||
| K | 44.6 | 38.7 | 31.6 | 23.2 | 13.7 |
| a | 21.47 | 18.74 | 16.01 | 13.28 | 10.55 |
| b | 1.350 | 1.292 | 1.234 | 1.176 | 1.118 |
| R2 | 0.973 | 0.960 | 0.956 | 0.967 | 0.976 |
| dk = aDb | |||||
| a | 1.736 | 1.421 | 1.005 | 0.539 | 0.232 |
| b | 0.937 | 0.940 | 0.987 | 1.115 | 1.301 |
| R2 | 0.975 | 0.984 | 0.986 | 0.984 | 0.957 |
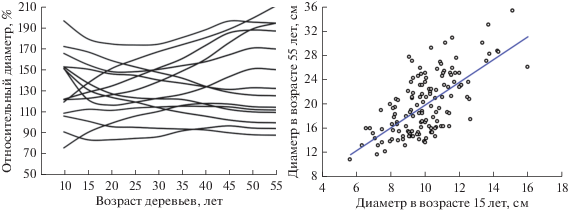
Исследования, проведенные на постоянных пробных площадях, а также данные сплошного отбора и измерения кернов на одной из них, показали определенную ограниченность приложения закона рангового роста деревьев, выдвинутого Е.Л. Маслаковым (1984), поскольку положение их в ценозе не остается постоянным, а со временем существенно изменяется (рис. 1), что приводит к постепенному ослабление тесноты связи текущего диаметра с исходным. Однако, несмотря на это, проводить отбор перспективных экземпляров при рубках ухода можно уже довольно уверенно с 15-летнего возраста. Так, из 18 деревьев I–II классов за 40 лет в IV–V классы перешло только одно (6%), а из 60 деревьев низших классов в основной полог древостоя перешло только 13 (22%). Из числа деревьев V класса только три поднялись на одну ступень. Особенно неустойчиво положение деревьев III класса, половина из которых понизили свое ранговое положение.
Рис. 1.
Динамика рангового положения деревьев в ценозе и характер связи между диаметром деревьев в возрасте 15 и 55 лет.
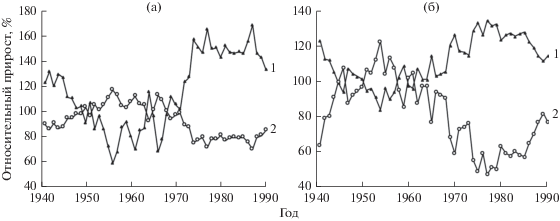
Ход роста каждого дерева, как нами (Демаков, 2000; Демаков, Исаев, Сафин, 2015) было установлено, сугубо индивидуален, что подтверждают в целом выводы других исследователей (Комин, 1970; Полюшкин, 1979, Каплина, Жиренко, Ильюшенко, 2009). Все особи в ценопопуляциях можно объединить в ряд групп, степень представительности которых неодинакова, а картина динамики их рангового положения зависит от выбора методики классификации, которую можно проводить либо по финальной величине диаметра, либо по характеру роста в пределах определенного отрезка времени. Так, если деревья в выборке классифицировать по величине диаметра, достигнутого ими на момент проведения исследования, то разные их группы четко различаются между собой по характеру роста уже с 10–15-летнего возраста (рис. 2). Если же классификацию проводить по динамике относительной величины диаметра в каждый конкретный момент времени, то картина получается совершенной иной: деревья, занимающие в молодом возрасте высокое положение, постепенно снижают его, а некоторые деревья низких рангов, наоборот, повышают его, постепенно догоняя первые. Часть деревьев, занимающих на начальном этапе роста среднее ранговое положение, не изменяют его с возрастом, а часть же снижают, постепенно уступая по величине диаметра деревьям, имеющим в молодости самое низкое положение. Следует отметить, что совокупности деревьев каждого класса не являются однородными и в них происходит постоянная перегруппировка особей, которые в процессе роста меняются между собой местами по величине текущего годичного прироста (рис. 3), регулируя свое ранговое положение в ценозе. Нами также было установлено, что наиболее угнетенные деревья в лесных культурах не образуют в последний этап своей жизни, продолжающийся иногда до 10–15 и более лет, годичных колец, расходуя весь запас питательных веществ на образование новой хвои.
Рис. 2.
Динамика рангового положения деревьев разных кластеров в 55-летних культурах сосны, выделенных по финальному диаметру (a) и характеру роста (б). 1–5 – кластеры деревьев.
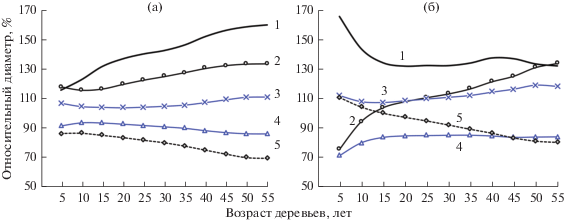
Рис. 3.
Динамика относительной величины радиального прироста деревьев разных типов (1, 2) среди совокупности господствующих (a) и подчиненных (б) особей в культурах сосны 1911 г. 1 – тип 1, 2 – тип 2.
Каковы же причины дифференциации деревьев в ценопопуляциях? В настоящее время среди ученых преобладает концепция, согласно которой полиморфизм древесных растений по их размерам обусловлен наследственными причинами, а конкуренция лишь усиливает их исходное неравенство (Кан-Ихи-Сакай, 1964; Романовский, 1994; Санников, Петрова, 2003; Романовский, Щекалев, 2014, 2016). Подобное объяснение этого процесса не подкреплено, однако, материалами генетических исследований и не вскрывает сути происходящих изменений. Некоторые исследователи (Keddy, 1990; Wedin, Tilmam, 1993) считают, что разные темпы роста деревьев могут определяться неодинаковой способностью особей к резервированию в своих органах ресурсов питательных веществ и их распределению между ними. Быстро растущие особи основную часть аккумулированной солнечной энергии вкладывают в прирост древесной массы и не создают, якобы, долговременных запасов питательных веществ, а медленно растущие, наоборот, оставляют их в резерве. Это заключение также не подтверждено данными о наличии запасов питательных веществ в тех или иных органах растений и характере их транспортирования от кроны к корням. Деревья отличаются друг от друга, по мнению этих исследователей, по длине окружности ствола, приходящейся на единицу листовой массы дерева, величина которой возрастает по мере снижения их рангового положения в ценозе. Наши исследования полностью подтвердили это положение, которое является, однако, следствием, а не причиной дифференциации деревьев. Изменение соотношения между массой хвои и размером кольца флоэмы, по которому происходит отток ассимилянтов, происходит, как это было установлено исследователями (Вомперский, Иванов, 1984; Кайбияйнен, Хари, 1985; Кайбияйнен и др., 1986; Хари и др., 1985; Иванов, Дубинин, 1992), в результате сбалансированности системы водного транспорта деревьев, саморегулирующейся на основе отрицательной обратной связи. Недостаточная влагообеспеченность кроны во время засухи вызывает снижение эффективности работы ассимиляционного аппарата и, как следствие, снижение прироста ствола, корней и хвои из-за недостатка продуктов фотосинтеза. Улучшение же влагообеспеченности кроны приводит к последующему увеличению прироста всех органов дерева. Площадь водопроводящей зоны восходящего потока воды, составляющая около 10% площади сечения всей заболони, ежегодно изменяется по величине, возрастая в сухие периоды и снижаясь во влажные (Вомперский, Иванов, 1984; Иванов, Дубинин, 1992). Ткань ксилемы хвойных функционирует в благоприятных условиях роста деревьев всего 10–12 лет, а в экстремальных – от 36 до 76. Число годичных слоев заболони, способных проводить воду, соответствует при этом числу живых мутовок у дерева (Кайбияйнен, Хари, 1985). Отношение площади сечения заболони к площади сечения ствола на высоте 1.3 м у деревьев I–III классов Крафта изменяется в пределах от 44 до 67%, а у особей IV–V классов – от 19 до 46% (Вомперский, Иванов, 1984). С увеличением возраста деревьев величина этого отношения у нормально развитых деревьев снижается, а у угнетенных же остается практически неизменной.
Одной из причин дифференциации особей в ценопопуляциях может являться разная скорость их старения, проявляющаяся не только в снижении с возрастом ширины годичного кольца и водопроводящей зоны заболони, но и в сокращении числа живых ветвей, активности очищения ствола от сучьев и ускорении ядрообразования. Скорость старения дерева можно отобразить математически (Зейде, 1972). Этот процесс, как показали расчеты, лучше всего описывает функция Dk = = Dmax[1 – exp(–A × 10–3a)], аппроксимирующая изменение диаметра дерева (Dk, см) с его возрастом (a, лет). Параметр Dmax этой функции характеризует предельный диаметр, которого может достичь дерево в процессе своего роста, а параметр же A – скорость его старения. Функции роста каждого дерева в ценопопуляции, отражающие потенциальные возможности особей и их раскрытие при взаимоотношении с окружающей средой (Карманова, 1976; Демаков, 2000), имеют и другие характерные параметры, которыми помимо Dmax и A, являются, применительно к нашей выборке, диаметр в возрасте 5 и 55 лет, а также их отношения между собой. Каждый из этих параметров изменяется в очень больших пределах (табл. 4), что свидетельствует о неоднородности ценопопуляции по скорости старения в ней деревьев. Сильнее всего варьирует в выборке (с. v. = 72.2%) отношение предельно возможного диаметра дерева, которого оно может достичь в процессе роста, к диаметру в возрасте 5 лет. Самый же низкий коэффициент вариации (с. v. = 16.0%) имеет отношение диаметра дерева в возрасте 55 лет к предельно возможному диаметру. Значения коэффициента вариации остальных параметров изменяются в пределах от 24.8 до 49.8%. Деревья каждого класса Крафта четко различаются между собой по средним значениям параметров функции старения (табл. 5), большинство из которых закономерно снижается от высших рангов к низшим. Значения же параметров скорости старения дерева (A) и отношение диаметра дерева в возрасте 55 лет к предельно возможному диаметру (D55 : Dmax) изменяются противоположным образом.
Таблица 4.
Вариабельность параметров функции скорости старения деревьев в 55-летних культурах сосны обыкновенной
| Параметр функции |
Значения статистических показателей (N = 138) | ||||||
|---|---|---|---|---|---|---|---|
| mx ± Mx | min | max | Sx | с. v. | a | e | |
| D5 | 4.1 ± 0.1 | 1.68 | 8.68 | 1.32 | 31.9 | 0.728 | 0.572 |
| D55 | 20.0 ± 0.4 | 10.96 | 35.32 | 4.95 | 24.8 | 0.552 | –0.288 |
| Dmax | 25.9 ± 1.1 | 12.40 | 77.6 | 12.90 | 49.8 | 3.139 | 16.55 |
| D55/Dmax | 0.83 ± 0.01 | 0.24 | 0.99 | 0.13 | 16.0 | –1.364 | 2.655 |
| D55 : D5 | 5.23 ± 0.16 | 2.39 | 13.23 | 1.93 | 36.9 | 1.541 | 3.363 |
| Dmax : D5 | 7.00 ± 0.43 | 2.42 | 44.53 | 5.05 | 72.2 | 4.023 | 23.61 |
| A | 40.88 ± 1.54 | 5.19 | 90.05 | 18.10 | 44.3 | 0.488 | –0.436 |
Примечание. mx ± Mx – среднее значение показателя и его ошибка; min, max – минимальное и максимальное значения в выборке,%; Sx – среднеквадратическое (стандартное) отклонение,%; с. v. – коэффициент вариации показателя,%; a, e – коэффициенты асимметрии и эксцесса. Параметры функции: D5 – диаметр древостоя в 5-летнем возрасте, см; D55 – диаметр древостоя в 55-летнем возрасте, см; Dmax – предельно возможный диаметр в возрасте свыше 100 лет, см; A – константа уравнения, характеризующая скорость старения дерева.
Таблица 5.
Значения параметров кривых роста деревьев разных классов Крафта в 55-летних культурах сосны
| Параметр функции |
Значения параметров у деревьев разных классов Г. Крафта: среднее значение и лимиты | ||||
|---|---|---|---|---|---|
| I (N = 7) | II (N = 14) | III (N = 60) | IV (N = 49) | V (N = 11) | |
| D5 | 4.38 (2.95–6.32) | 5.08 (2.22–8.68) | 4.25 (1.68–7.68) | 3.89 (1.76–6.04) | 3.27 (2.04–5.79) |
| D55 | 32.3 (30.1–35.3) | 27.6 (25.9–30.0) | 22.0 (18.4–25.8) | 16.0 (14.1–18.3) | 13.1 (11.0–14.0) |
| Dmax | 51.4 (48.8–57.2) | 41.0 (27.0–77.6) | 28.3 (19.6–41.3) | 17.6 (14.6–23.9) | 14.2 (12.4–15.9) |
| D55 : Dmax | 0.63 (0.54–0.71) | 0.72 (0.35–0.96) | 0.79 (0.60–0.96) | 0.91 (0.68–0.99) | 0.92 (0.83–0.99) |
| D55 : D5 | 7.99 (5.59–10.2) | 6.42 (3.00–13.2) | 5.63 (2.88–13.0) | 4.39 (2.39–9.07) | 4.30 (2.40–5.75) |
| Dmax : D5 | 13.1 (7.85–17.9) | 9.89 (3.12–24.9) | 7.44 (3.03–21.7) | 4.94 (2.42–15.6) | 4.69 (2.42–6.08) |
| A | 19.6 (13.9–24.7) | 28.4 (7.92–62.8) | 32.1 (16.6–59.5) | 53.8 (20.7–87.1) | 57.2 (34.9–90.1) |
Примечание. Обозначение параметров функции см. табл. 4.
Математическая модель динамики диаметра деревьев позволяет без особых затруднений оценить величину исчерпания потенциальных возможностей их роста по отношению Di : Dmax. Результаты проведенных нами расчетов (рис. 4), показали, что деревья IV–V классов Крафта практически полностью исчерпывают потенциал роста уже к 40–50 г., почти не увеличивая диаметр ствола. Оставлять их в древостое, исходя из этого, не имеет смысла. Рубку деревьев остальных классов в оцененной ценопопуляции целесообразно проводить в 85–90 лет, так как в это время они также исчерпывают свои потенциальные возможности роста.
Рис. 4.
Динамика исчерпания потенциальных возможностей роста деревьями сосны разных классов Крафта (a), а также величины пропорции между высотой и площадью сечения их ствола (б). 1–5 – классы роста Крафта.
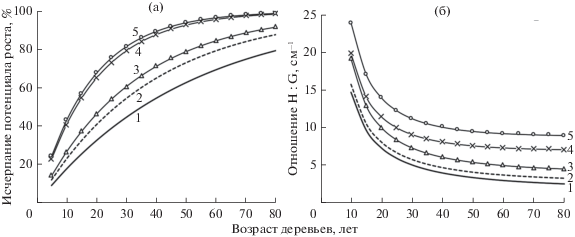
Любое дерево, представляющее собой высокоорганизованную саморегулирующуюся систему, в процессе своего роста приспосабливается к условиям среды, изменяя размеры своих органов таким образом, чтобы они наилучшим образом выполняли в сложившейся экологической обстановке возложенные на них функции. Достигается это за счет перераспределения ассимилянтов, отражаясь на скорости роста ствола в высоту и толщину. Величина пропорций между высотой дерева и диаметром или площадью сечения ствола, исходя из этой концепции, может являться хорошим диагностическим признаком состояния его жизнеспособности и рангового положения в ценопопуляции (Ключюс, 1986; Демаков, 2000). Чем больше величина отношения Н : D или Н : G, тем больше прирост в высоту преобладает над приростом в толщину и тем большая часть питательных веществ из имеющихся ресурсов направляется деревом на образование хвои и верхушечного побега. Деревья разных классов, как это видно на представленном графике, четко различаются между собой в любом возрасте по величине пропорции Н : G, которая, однако, отражает не причину, а следствие процесса их дифференциации.
Причины изменения рангового положения деревьев могут быть самыми разными: их наследственные свойства, особенности реакции на погодные условия, взаимоотношение в надземной и подземной сферах с соседними особями, повреждение вредными насекомыми и грибами. Одной из них являются также особенности архитектоники корневых систем деревьев (Baldwin, 1976; Wilson, 1988; Grubb, 1994; Kadmon, 1995; Тихонова, 2013; Милютин и др., 2013), которые у особей низших рангов расположены ближе к поверхности почвы и менее разветвлены, чем у наиболее крупных. Еще одной причиной дифференциации деревьев по их размерам в ценопопуляциях может являться неоднородность почвенного покрова в пределах биотопов (Карпачевский, 1977, 2005), приводящая к образованию их биогрупп различной величины, густоты и производительности (Внучков, 1976; Тябера, 1980; Бузыкин и др., 1985; Гордина, 1985; Усольцев, 1985, 2013; Сеннов, 1993; Ledermann, Stage, 2001; Mabvurira, Miina, 2002; Stadt et al, 2007; Грабарник, 2010). Результаты исследования этого явления, однако, довольно противоречивы и не позволяют пока сделать однозначного заключения. Взаимовлияние деревьев в ценозах проявляется, по данным наших исследований (Демаков и др., 2011, 2013), довольно слабо и их размеры зависят в большей степени не от расстояния до соседних особей и их размеров, а от экологической неоднородности биотопа, что подтверждает вывод И.С. Марченко (1995) о наличии зон, благоприятных или неблагоприятных для их роста. В благоприятных зонах густота древостоя и размеры деревьев выше, а в неблагоприятных, наоборот. Размеры же деревьев, произрастающих в активных зонах группами разной густоты, существенно не различаются между собой.
Выводы. 1. Дифференциация деревьев по их размерам начинается в ценопопуляциях очень рано и продолжается до глубокой старости. В большинстве древостоев преобладают деревья среднего рангового положения. Доля господствующих и лидирующих деревьев по числу стволов невелика, однако по площади сечения стволов составляет в среднем около 50%, достигая в ряде случаев даже 80%. Доля деревьев IV–V классов Крафта, не имеющих особых перспектив развития, составляет в среднем по числу стволов 31.6%, а по площади сечения – всего лишь 12.3–15.9%.
2. Влияние возраста и густоты древостоев на характер их ранговой структуры практически не проявляется. Только при крайне низкой густоте лесных культур, составляющей до 1.0 тыс. экз. га–1, резко увеличивается доля деревьев III класса Крафта, что указывает на снижение интенсивности процесса дифференциации особей.
3. Средний диаметр деревьев каждого класса Г. Крафта очень тесно связан со средним диаметром всего древостоя, что аппроксимирует уравнение Dk = aDb. Опираясь на него можно провести оценку ранговой структуры ценопопуляций не только по материалам фактического учета, но также таблицам хода роста насаждений и данным таксационных описаний. Это позволяет объективно оценить возможный запас и стоимость древесины, получаемой при рубках ухода, и оценить их потенциальную рентабельность.
4. Ранговое положение деревьев в ценопопуляциях не остается стабильным в течение их жизни, изменяясь по мере роста в ту или иную сторону, что полностью подтверждает выводы многих исследователей. Особенно неустойчиво положение деревьев III класса, половина из которых понижают со временем свой ранг. Статус деревьев высших и низших рангов более постоянен. Так, подавляющая часть деревьев IV–V классов Крафта со временем неизбежно погибает, а из деревьев I–II классов в низшие переходит не более 6–8%, что позволяет производить отбор перспективных экземпляров при уходе за лесом довольно уверенно уже с 10–15-летнего возраста.
5. Причины изменения рангового положения деревьев могут быть самыми разными и остаются пока до конца не выявленными, требующими проведения дальнейших исследований. Одной из этих причин является разная скорость исчерпания жизненного потенциала особей, которую можно отобразить математически через скорость изменения с возрастом диаметра дерева. Деревья сосны IV–V классов Крафта практически полностью исчерпывают потенциал роста уже к 40–50 г., поэтому оставлять их в древостое не имеет смысла. Реализация жизненного потенциала у деревьев I–II классов завершается не ранее 80–90 лет.
6. Хорошим диагностическим признаком жизнеспособности деревьев и их дальнейшего статуса в ценопопуляциях является отношение их высоты к диаметру или площади сечения ствола.
Список литературы
Барнишкис Е.К. Выживание деревьев при естественном изреживании в сосновых насаждениях // Лесоведение. 1972. № 5. С. 33–41.
Белов С.В. Лесоводство. М.: Лесн. пром-сть, 1983. 352 с.
Бузыкин А.И., Гавриков B.Л., Секретенко О.П., Хлебопрос Р.Г. Анализ структуры древесных ценозов. Новосибирск: Наука, 1985. 95 с.
Внучков В.Т. Горизонтальная структура древостоев сосны Казахского мелкосопочника // Лесоведение. 1976. № 5. С. 56–62.
Вомперский С.Э., Иванов А.И. Связь площади поперечного сечения заболони с массой хвои сосны обыкновенной // Лесоведение. 1984. № 3. С. 60–65.
Гордина Н.П. Пространственная структура и продуктивность сосняков нижнего Енисея. Красноярск: Институт леса и древесины, 1985. 126 с.
Грабарник П.Я. Анализ горизонтальной структуры древостоя: модельный подход // Лесоведение. 2010. № 2. С. 77–85.
Демаков Ю.П. Диагностика устойчивости лесных экосистем: методологические и методические аспекты. Йошкар-Ола: Периодика Марий Эл, 2000. 415 с.
Демаков Ю.П., Исаев А.В., Нехаев И.Н. Характер освоения деревьями жизненного пространства в пойменных биогеоценозах // Науч. труд государственного природного заповедника “Большая Кокшага”. Вып. 6. Йошкар-Ола: Поволжский государственный технологический университет, 2013. С. 163–184.
Демаков Ю.П., Исаев А.В., Сафин М.Г. Закономерности динамики прироста деревьев сосны в различных типах леса заповедника // Там же. Вып. 7. Йошкар-Ола: Поволжский государственный технологический университет, 2015. С. 101–138.
Демаков Ю.П., Мухортов Д.И., Нуреева Т.В. Pro motus coniferarum – Программа для расчета динамики состояния хвойных древостоев на постоянных пробных площадях. Свидетельство о государственной регистрации программы для ЭВМ. Заявка № 2017614494; заявитель и правообладатель ПГТУ; Заяв. 16.05.2017, зарег. 05.07.2017, опубл. 05.07.2017.
Демаков Ю.П., Сафин М.Г., Нехаев И.Н. Пространственное распределение и взаимовлияние деревьев в чистых сосновых древостоях // Лесные экосистемы в условиях изменения климата: биологическая продуктивность и дистанционный мониторинг: Матер. междунар. науч.-практ. семинара. Йошкар-Ола, 2–3 сентября 2011:, 2011. Йошкар-Ола. Изд-во МарГТУ. 2011. С. 93–102.
Демаков Ю.П., Пуряев А.С., Черных В.Л., Черных Л.В. Использование аллометрических зависимостей для оценки фитомассы различных фракций деревьев и моделирования их динамики // Вестник Поволжского государственного технологического университета. Серия: Лес. Экология. Природопользование. 2015. № 2. С. 19–36.
Дударев А.Д. Итоги 30-летних лесоводственно-таксационных наблюдений на постоянных пробных площадях в Брянском опытном лесничестве // Научные записки Воронежского лесотехнического института. Воронеж, 1956. Т. 15. С. 121–128.
Зейде Б.Б. О математической природе процесса старения деревьев // Дендроклиматохронология и радиоуглерод. Каунас, 1972. Изд-во: Каунас, Институт ботаники АН Литовской ССР – ЛФТИ, 1972. С. 169–174.
Иванов А.И., Дубинин А.И. Площадь сечения заболони и площадь зоны транзита влаги в ней у сосны обыкновенной // Лесоведение. 1992. № 5. С. 28–37.
Кайбияйнен Л.К., Хари П. Сбалансированность системы водного транспорта у сосны обыкновенной. I. Пути движения влаги в ксилеме // Лесоведение. 1985. № 5. С. 23–28.
Кайбияйнен Л.К., Хари П., Сазонова Т.А., Мякеля А. Сбалансированность системы водного транспорта у сосны обыкновенной. III. Площадь проводящей ксилемы и масса хвои // Лесоведение. 1986. № 1. С. 31–37.
Кан-Ихи-Сакай. Конкурентоспособность растений, ее наследуемость и некоторые связанные с ней проблемы // Механизмы биологической конкуренции. М.: Мир, 1964. С. 309–331.
Каплина, Н.Ф., Жиренко Н.Г., Ильюшенко А.Ф. Погодичная динамика фитомассы и продукции древостоя дуба южной лесостепи: методические подходы и результаты // Продукционный процесс и структура лесных биогеоценозов: теория и эксперимент (памяти А.И. Уткина) / Отв. ред. М.Г. Романовский. М.: Товарищество научных изданий КМК, 2009. С. 68–92.
Карманова И.В. Математические методы изучения роста и продуктивности растений. М: Наука, 1976. 223 с.
Карпачевский Л.О. Пестрота почвенного покрова в лесном биогеоценозе. М.: Изд-во Московского государственного университета, 1977. 312 с.
Карпачевский Л.О. Экологическое почвоведение. М.: ГЕОС, 2005. 336 с.
Ключюс А.А. Метод выделения гомогенных групп деревьев для изучения и прогноза вероятности их отпада // Мониторинг лесных экосистем. Каунас, 5–6 июня 1986. Изд-во: ЛитСХА АН ЛитССР, 1986. С. 325–327.
Комин Г.Е. Изменение рангов деревьев по диаметру в древостое // Лесообразовательный процесс на Урале. Свердловск: Изд-во Уральского филиала АН СССР, 1970. Вып. 67. С. 252–262.
Марченко И.С. Биополе лесных экосистем. Брянск: Придесенье, 1995. 188 с.
Маслаков Е.Л. Формирование сосновых молодняков. М.: Лесн. пром-сть, 1984. 168 с.
Милютин Л.И., Новикова Т.Н., Тараканов В.В., Тихонова И.В. Сосна степных и лесостепных боров Сибири. Новосибирск: Гео, 2013. 128 с.
Нестеров В.Г. Вопросы современного лесоводства. М.: Сельхозгиз, 1961. 384 с.
Полюшкин Ю.В. Изменчивость радиального прироста древостоев как источник информации для прогнозирования динамики экосистем // Проблемы прогностических исследований природных явлений. Новосибирск: Наука, 1979. С. 154–167.
Романовский М.Г. Полиморфизм древесных растений по количественным признакам. М.: Наука, 1994. 96 с.
Романовский М.Г., Щекалев Р.В. Система вида у древесных растений. М.: Товарищество научных изданий КМК, 2014. 212 с.
Романовский М.Г., Щекалев Р.В. Продукционный бюджет кроны дерева: гиганты и карлики в древостое // Лесоведение. 2016. № 6. С. 438–444.
Санников С.Н. Петрова И.В. Дифференциация популяций сосны обыкновенной, Екатеринбург: Изд-во БС УрО РАН, 2003. 246 с.
Сеннов С.Н. Итоги экспериментального изучения конкуренции в древостоях // Известия Санкт-Петербургской лесотехнической академии. 1993. № 11. С. 160–172.
Сукачев В.Н. Растительные сообщества (введение в фитосоциологию). Л.-М.: Книга, 1928. 232 с.
Тихонова И.В. Карликовые сосны Ширинской лесостепи // Лесоведение. 2013. № 2. С. 22–30.
Тябера А.П. Вопросы территориального размещения деревьев в сосновых древостоях // Известия высших учебных заведений: Лесной журн. 1980. № 5. С. 5–8.
Усольцев В.А. Оценка показателей продуктивности в биогруппах разной густоты // Лесоведение. 1985. № 2. С. 68–78.
Усольцев В.А. Продукционные показатели и конкурентные отношения деревьев. Исследование зависимостей. Екатеринбург: Уральский государственный лесотехнический университет, 2013. 553 с.
Хари П., Кайбияйнен Л.К., Сазонова Т.А., Мякеля А. Сбалансированность системы водного транспорта у сосны обыкновенной. II. Активная ксилема // Лесоведение. 1985. № 5. С. 74–77.
Baldwin J.P. Competition for plant nutrients in sol; a theoretical approach // The J. Agricultural Science. 1976. V. 87. № 2. P. 341–356.
Grubb P.J. Root competition in soils of different fertility: a paradox resolved? // Phytocoenologia. 1994. V. 24. № 1–4. P. 495–505.
Kadmon R. Plant competition along soil moisture gradients: A field experiment with the desert annual Stipa capensis // J. Ecology. 1995. V. 83. № 2. P. 253–262.
Keddy P.A. Competitive hierarchies and centrifugal organization in plant communities / J.B. Grace and D. Tilman. Perspectives on Plant Competition. U.S.A., San-Diego: A-cademic Press, 1990. P. 266–290 (Eds.).
Ledermann T., Stage A.R. Effects of competitor spacing in individual-tree indices of competition // Canadian J. Forest Research. 2001. V. 31. № 12. P. 2143–2150.
http://web.forestry.ubc.ca/biometrics/Documents/leMay_ staudhammer_paper_March291.pdf/. Дата обращения 15.11.2017 г.
Mabvurira D., Miina J. Individual-tree growth and mortality models for Eucalyptus grandis (Hill) Maiden plantations in Zimbabwe // Forest Ecology and Management. 2002. V. 161. № 1. P. 231–245.
Pastorella F., Paletto A. Stand structure indices as tools to support forest management: an application in Trentino forests (Italy). J. Forest Science. 2013. V. 59. № 4. P. 159–168.
Stadt K.J., Huston C., Coates K.D., Feng Z., Dale M.R.T., Lieffers V.J. Evaluation of competition and light estimation indices for predicting diameter growth in mature boreal mixed forests // Annals of Forest Science. 2007. V. 64. № 5. P. 477–490.
Szmyt J., Korzeniewicz R. Do natural processes at the juvenile stage of stand development differentiate the spatial structure of trees in artificially established forest stands? Leśne Prace Badawcze (Forest Research Papers). 2014. V. 75. № 2. P. 171–179.
Wedin D., Tilman D. Competition among grasses along a nitrogen gradient: initial conditions and mechanisms of competition // Ecological Monographs. 1993. V. 63. № 2. P. 199–229.
Wilson J.B. Shoot competition and root competition // J. Applied Ecology. 1988. V. 25. № 1. P. 279–296.
Дополнительные материалы отсутствуют.