

Лесоведение, 2019, № 4, стр. 286-293
Горизонтальная структура древостоя и подроста ельника разнотравно-черничного средней тайги Республики Коми
А. В. Манов *
a Институт биологии Коми НЦ УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: manov@ib.komisc.ru
Поступила в редакцию 20.08.2011
После доработки 26.03.2018
Принята к публикации 04.02.2019
Аннотация
В статье рассматривается пространственная структура древостоя и подроста коренного разнотравно-черничного ельника, спонтанно развивающегося в лесном массиве подзоны средней тайги. Представлены результаты анализа размерной и горизонтальной структуры разновозрастного елового древостоя. Показана степень дифференциации изучаемого растительного сообщества с учетом значений изменчивости диаметров деревьев и высоты подроста, оцененная с использованием основных статистических показателей описательной статистики. В ельнике отмечена значительная вариабельность деревьев по диаметру, подроста – по высоте. Приводятся расчетные данные пространственной статистики, полученныен с применением анализа точечных процессов с помощью проверки нулевой гипотезы о полной пространственной случайности. Выявлен случайный тип пространственной структуры как при совместном размещении деревьев, так и раздельно по категориям крупности, видам и состоянию. Установлено, что деревья лиственных пород, входящие в состав древостоя, склонны к группированию в радиусе от 4 м. Отмечен групповой характер размещения подроста на площади. С помощью маркированных точечных процессов раскрыты пространственные отношения между подростом разных категорий крупности. Чем мельче соседний подрост, тем выше между ними конкурентные взаимоотношения.
К настоящему времени массивы коренных еловых экосистем на европейском Севере России сохранились в основном на заповедных и труднодоступных территориях (Ярошенко и др., 2001). Функционирование этих лесных сообществ, где отсутствует прямое антропогенное воздействие, обусловлено природными процессами и экологическими факторами, создающимися в той или иной экосистеме. Исследуемый нами разнотравно-черничный ельник расположен вдали от проезжих дорог, не затронут рубками и разработками других природных ресурсов, строительством и аэротехногенным загрязнением, не испытывает влияния рекреации.
Ельники черничные – основная растительная ассоциация незаболоченных хвойных сообществ средней тайги (Василевич, 2004). Здесь они формируют, как правило, смешанные по составу, разновозрастные древостои. В древесном ярусе всегда присутствует сухостой. Лесовозобновительный процесс в них непрерывен (Дыренков, 1984; Коренные …, 2006).
В смешанных по составу и сложных по структуре лесных насаждениях проявляются многосторонние взаимовлияния между особями одного и разных видов растений, а также между растениями разных ярусов, которые проявляются в конкурентных межвидовых и внутривидовых отношениях за питательные вещества и жизненное пространство (Цветков, 2004). Однако напряженность конкуренции в них неодинакова, что связано с неравномерностью размещения деревьев и различиями их жизненного состояния. В результате она приводит к дифференциации размеров особей. В итоге конкурентные взаимоотношения обусловливают формирование древостоев определенной пространственной структуры, которая во многом определяет устойчивость и продуктивность экосистем (Кузьмичев, 2013).
Цель работы – определение закономерностей горизонтальной структуры древостоя и подроста разновозрастного ельника разнотравно-черничного в естественно развивающемся лесном массиве средней тайги.
ОБЪЕКТЫ И МЕТОДИКА
Исследования горизонтальной структуры древостоя разновозрастного разнотравно-черничного ельника проводили в западной части Республики Коми в условиях средней тайги на территории государственного природного заказника регионального значения “Ляльский” площадью 700 га. Заказник имеет лесной профиль. Территория заказника представлена покрытыми лесом плоскими водораздельными поверхностями с уклонами 1°–3°, прорезанными долинами ручьев (Кадастр …, 2014). Почвы здесь преимущественно подзолистые, сформированные на двучленных и одночленных отложениях (Забоева, 1975).
В еловом насаждении заложена постоянная пробная площадь прямоугольной формы размером 20 × 95 м с размеченной колышками сеткой квадратов (5 × 5 м) для определения положения основания стволов деревьев в прямоугольной системе координат (X, Y).
Постоянная пробная площадь (62°15.661′ с.ш., 50°40.394′ в.д.) расположена в еловом массиве спонтанно развивающейся тайги на пологом склоне водосборной площади руч. Мая. При таксации насаждения установлено, что древостой образован елью (Picea obovata Ledeb.), березами (Betula pubescens Ehrh. и B. pubescens Ehrh), пихтой (Abies sibirica Ledeb.), осиной (Populus tremula L.) и сосной (Pinus sylvestris L.). Ярусность его не выражена. Возраст преобладающих по стволовому запасу деревьев ели колеблется от 55 до 295 лет. Подрост представлен теми же видами древесных растений, что и древостой. Возраст преобладающего по количеству мелкого елового подроста составляет 10–25 лет. Подлесок редкий из Juniperus communis L., Rosa sp., Salix sp., Sorbus aucuparia L., Lonicera pallasii Ledeb. В травяно-кустарничковом ярусе с общим покрытием 40–50% господствует Vaccinium myrtillus L., присутствуют V. vitis-idaea L., Rubus saxatilis L., Maianthemum bifolium (L.) F.W. Schmidt, Trientalis europaea L., Gymnocarpium dryopteris (L.) Newm., Galium boreale L., Vicia sylvatica L., Linnaea borealis L., Epilobium sp., Deschampsia cespitosa (L.) Beauv., Geranium sylvaticum L., Equisetum sylvaticum L., Solidago virgaurea L., Lycopodium clavatum L., Sonchus arvensis L., Carex sp., Luzula pilosa (L.) Willd., Fragaria vesca L., Pyrola rotundifolia L., Oxalis acetosella L. Моховой покров хорошо развит, его общее покрытие 80–90%. Преобладают Pleurozium schreberi (Brid.) Mitt. и Hylocomium splendens (Hedw.) B.S.G. Небольшими пятнами встречается Sphagnum sp.
Краткая таксационная характеристика объекта исследования приведена в табл. 1.
Таблица 1.
Таксационная характеристика древостоя и подроста ельника разнотравно-черничного
| Состав | Порода | Численность, шт. га‒1 | Сумма площадей сечения, м2 га‒1 | Запас древесины, м3 га‒1 | Средние | ||||
|---|---|---|---|---|---|---|---|---|---|
| растущих | сухих | растущих | сухих | растущих | сухих | высота, м | диаметр, cм | ||
| Древостой | |||||||||
| 4Е4Б2Пх + Ос,С | Ель Береза Пихта Осина Сосна Всего |
389 189 121 11 5 715 |
95 0 5 0 0 100 |
11.3 9.3 3.4 0.9 0.6 25.5 |
3.5 0 0.1 0 0 3.6 |
104.5 94.2 33.3 13.1 7.0 252.1 |
38.8 0 1.1 0 0 39.9 |
15 19 14 24 28 |
17 24 17 31 38 |
| Подрост | |||||||||
| 7Е3Пх + Б ед. Ос,С | Ель Пихта Береза Осина Сосна Всего |
4716 1489 226 15 5 6451 |
137 5 0 0 0 142 |
– – – – – |
– – – – – |
– – – – – |
– – – – – |
1.0 1.1 1.0 2.5 0.6 |
– – – – – |
В 2004 г. на пробной площади проведен сплошной перечет растущих и сухостойных деревьев с диаметром на высоте 1.3 м ≥6 см. Возраст деревьев находили по образцам древесины (кернам), отобранным у шейки корня. Подрост (древесные растения высотой ≥0.25 м и диаметром ствола на высоте 1.3 м <6 см) учитывали на всей пробной площади. Возраст мелкого хвойного подроста подсчитывали по мутовкам. Среднюю высоту древостоя определяли с помощью графика высот, тип леса – по В.Н. Сукачеву, С.В. Зонну (1961). Анализ таксационных материалов проводили согласно Лесотаксационному справочнику (1986).
Степень дифференциации растительного сообщества изучаемого ельника рассматривалась с учетом значений изменчивости диаметров деревьев на высоте 1.3 м и высоты подроста по И.И. Гусеву (2002). Для этого использовались основные статистические показатели описательной статистики, рассчитанные средствами компьютерной программы Excel.
С целью выявления особенностей горизонтальной структуры древостоя и подроста рассмотрено положение растущих деревьев на постоянной пробной площади: крупных (диаметром на высоте 1.3 м ≥19 см), мелких (диаметром на высоте 1.3 м – 6–18.9 см), хвойных и лиственных; сухостойных, а также живого подроста: крупного (высотой ≥1.5 м), среднего (высотой – 0.5–1.4 м), мелкого (высотой <0.5 м), хвойного и лиственного. Сухой подрост в анализе не учитывали из-за малой выборки статистических показателей, что ведет к ослаблению пространственных эффектов (Фардеева и др., 2008).
С учетом оценки горизонтальной структуры древостоя ельника разнотравно-черничного применялся подход пространственной статистики, который включал в себя методы точечных процессов (Schabenberger, Gotway, 2005). Размещение деревьев и подроста на плоскости (двумерное пространство) может быть рассмотрено как точечный процесс, где “точки” – это положение оснований их стволов относительно координат X, Y. Дополнительная информация (размерные, видовые и другие характеристики растений) рассматривается как маркированные точечные процессы (Stoyan, Penttinen, 2000).
Математической моделью для анализа точечных процессов послужил однородный процесс Пуассона, при котором “точки” располагаются случайным образом (Illian et al., 2008). В качестве статистического инструмента для оценки пространственных взаимодействий в точечных процессах нами использовалась парная корреляционная функция g(r) (Stoyan, Penttinen, 2000). Классический анализ точечных процессов основан на проверке нулевой гипотезы о полной пространственной случайности, которую проводили методом Монте-Карло, заключающимся в оценке значимости отклонения эмпирического значения ĝ(r)-функции от теоретического g(r) (Wiegand, Moloney, 2004). Области с верхними (95%) и нижними (5%) доверительными интервалами принятия нулевой гипотезы о полной пространственной случайности получены с помощью 999 генераций модели однородного процесса Пуассона. Высокое число генераций позволяет уменьшить вероятность ошибки первого рода (Grabarnik, 2011). Выход кривой функции ĝ(r) на расстоянии (r) за пределы верхнего доверительного интервала (ĝ(r) > 1) свидетельствует об агрегировании (скоплении, группировании) точечных процессов. Выход за пределы нижнего доверительного интервала (ĝ(r) < 1) говорит о регулярности (равномерности, упорядоченности) точечных процессов на расстоянии (r). Случайные точечные процессы (однородный процесс Пуассона) отмечаются тогда, когда значение ĝ(r) находится в пределах области принятия вышеназванной гипотезы (ĝ(r) = 1).
Исследование пространственных отношений между древесными растениями проводилось с помощью кросс-корреляционной функции gij(r), которая расширяет парную корреляционную функцию g(r) для маркированных точечных процессов. Ее значения позволяют проверить гипотезу о независимости или случайности маркирования точечных процессов (Illian et al., 2008). Методы оценки функции gij(r) аналогичны методу определения функции g(r).
Обработка пространственных данных проводилась нами в статистическом пакете Spatstat (Baddeley, Turner, 2005) в программной среде R (http://r-project.org).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Обычно при описании характеристик фитоценоза особое внимание уделяется оценке степени дифференциации деревьев по диаметру и подроста по высоте. При этом размерная структура древостоя и его плотность находятся в прямой зависимости от размещения деревьев в пространстве (Кузьмичев, 2013). Статистический анализ отдельно взятых компонентов растительной среды и происходящих внутри ее процессов позволяет с определенной вероятностью и степенью достоверности познать биологические явления природы как в частностях, так и в целом (Гусев, 2002).
По расчетным данным, приведенным в табл. 2, выявлена значительная изменчивость (C.V. > 30%) диаметра стволов живых деревьев в исследуемом древостое. Ряд распределения деревьев по диаметру растянут (E < 0) и имеет малое правостороннее смещение (0 < A < 0.5) относительно среднего диаметра дерева, что указывает на небольшое преобладание тонкомерных деревьев. Вариабельность высоты живого подроста значительная (C.V. > 30%). Преобладают древесные растения ниже средней высоты (A > 1), густо сгруппированные около среднего значения (E > 0).
Таблица 2.
Статистика рядов распределения деревьев по диаметру и подроста по высоте
| Показатель | $\bar {x}$ | m | σ | p | C.V. | A | E |
|---|---|---|---|---|---|---|---|
| Диаметр, см | 19.2 | 0.8 | 9.2 | 3.9 | 48.1 | 0.2 | –0.8 |
| Высота, м | 1.1 | 0.02 | 0.8 | 2.3 | 78.9 | 1.7 | 3.2 |
Статистический анализ данных по пространственной структуре смешанного по составу и сложного по строению древостоя ельника разнотравно-черничного показал, что размещение растущих деревьев на площади носит случайный характер, т.е. модель пространственных точечных процессов соответствует однородному процессу Пуассона (рис. 1а). В рассматриваемом древостое отсутствуют пространственные взаимосвязи в размещении деревьев крупной и мелкой размерных категорий по отдельности (рис. 1б, 1в). Структура размещения хвойных деревьев на площади также является случайной (рис. 1г). Небольшое отклонение от пуассоновского процесса прослеживается в размещении лиственных деревьев на расстояниях от 4 м (рис. 1д). Не отмечается значимых зависимостей в размещении сухостойных деревьев на площади (рис. 1е).
Рис. 1.
Оценка эмпирической парной корреляционной функции ĝ(r) (1) и области принятия нулевой гипотезы о случайности точечных процессов (2). Для деревьев: а – всех размеров; б – крупных; в – мелких; г – хвойные; д – лиственных; е – сухостойных; подроста: ж – в целом; з – крупного; и – среднего; к – мелкого; л – хвойного; м – лиственного.
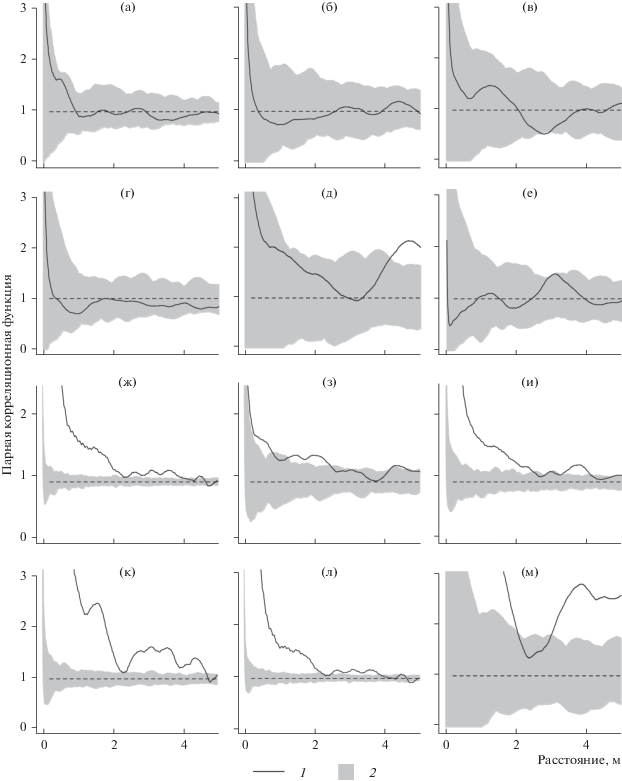
Процессы саморегуляции горизонтальной структуры фитоценоза исследуемого ельника отмечаются в размещении растущего подроста, для которого характерно групповое распространение (рис. 1ж–1м). При этом степень его агрегированности снижается с увеличением категории крупности растений (рис. 1з–1к). Подрост как хвойных, так и лиственных видов деревьев образует большие скопления на площади в радиусе до 2 м (рис. 1л–1м).
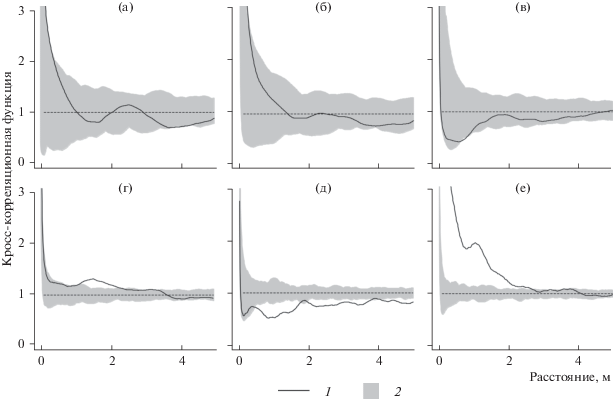
Протестированные пространственные отношения между деревьями крупной и мелкой категорий, хвойными и лиственными видами, а также между растущими и сухостойными демонстрируют независимое друг от друга размещение (рис. 2а–2в). В отличие от деревьев, позиция подроста на площади показывает конкурентные взаимоотношения (рис. 2г–2е). Степень этих взаимоотношений зависит от размера соседних особей. Чем мельче окружающие растения, тем сильнее между ними конкуренция.
Рис. 2.
Оценка эмпирической кросс-корреляционной функции ĝij(r) (1) и области принятия нулевой гипотезы о случайности маркирования точечного процесса (2) для каждой пары категорий древесных растений. Деревья: а – крупные/мелкие; б – хвойные/лиственные; в – растущие/сухостойные; подрост: г – крупный/средний; д – крупный/мелкий; е – средний/мелкий.
Отмеченная случайность размещения деревьев, в том числе сухостойных, на площади согласно результатам исследований ряда авторов (Ипатов, Тархова, 1975; Плотников, 1979; Проскуряков, 1983; Стороженко, 2007; Кузьмичев, 2013; и др.), характерна для устойчиво развивающихся древостоев коренных лесных сообществ. Такая структура обусловлена смешанным составом древостоя, его разновозрастностью и теневыносливостью ели – породы-лесообразователя (Цветков, 2004). Здесь ель – мощный трансформатор экологических условий для подчиненных ценопопуляций – обусловливает их пространственный ритм размещения.
Известно, что групповое размещение растений в естественно развивающихся фитоценозах происходит на стадии заселения территории, что обусловлено их генезисом. Ослабление агрегированности особей древесных растений связано с элиминацией в группах растений с высокой плотностью произрастания вследствие ограничения жизненного пространства и снижения количества питательных веществ в почве (Ипатов, Тархова, 1975). В таких лесных сообществах конкуренция между древесными растениями выражается в основном за элементы минерального питания, в частности за доступные формы азота (Факторы …, 1983). Разрушение групп деревьев вследствие конкурентных взаимоотношений на определенном этапе онтогенеза ведет к выравниванию напряженности между характеризуемыми категориями деревьев, ввиду чего размещение особей древесных растений на площади становится случайным. Таким образом, в еловом сообществе создаются такие условия, которые необходимы для непрерывного обеспечения постоянной смены поколений эдификатора и сопутствующих пород, что является одним из основных характеристик устойчивого развития леса (Стороженко, 2007).
Заключение. Выявленные пространственные закономерности размещения древесных растений на площади свидетельствуют о сложной горизонтальной структуре древостоя и подроста коренного ельника разнотравно-черничного. По мере роста и отпада деревьев положение их на площади меняется. Для древесных растений, появившихся на этапе поселения, характерно скопление особей. С развитием древостоя при увеличении размеров деревьев степень их сгруппированности снижается. Деревья, образующие древесный ярус, имеют случайный характер размещения на площади. Пространственные взаимоотношения между деревьями проявляются лишь в молодом возрасте. Представленные экспериментальные данные о характере горизонтальной структуры исследуемого древостоя соответствуют природе коренных разновозрастных ельников севера таежной зоны.
Список литературы
Василевич В.И. Ельники черничные европейской России // Ботанический журн. 2004. Т. 89. № 11. С. 1728–1739.
Гусев И.И. Моделирование экосистем. Архангельск: Изд-во АГТУ, 2002. 112 с.
Дыренков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174 с.
Забоева И.В. Почвы и земельные ресурсы Коми АССР. Сыктывкар: Коми книжн. изд-во, 1975. 344 с.
Ипатов В.С., Тархова Т.Н. Количественный анализ ценотических эффектов в размещении деревьев по территории // Ботанический журн. 1975. Т. 60. № 9. С. 1237–1250.
Кадастр особо охраняемых природных территорий Республики Коми / Под ред. С.В. Дегтевой, В.И. Пономорева. Сыктывкар: ИБ Коми НЦ УрО РАН, 2014. 428 с.
Коренные еловые леса Севера: биоразнообразие, структура, функции / Под ред. К.С. Бобковой, Э.П. Галенко. СПб.: Наука, 2006. 337 с.
Кузьмичев В.В. Закономерности динамики древостоев: принципы и модели. Новосибирск: Наука, 2013. 208 с.
Лесотаксационный справочник для Северо-Востока европейской части СССР (нормативные материалы для Архангельской, Вологодской областей и Коми АССР). Архангельск: Изд-во АИЛиЛХ, 1986. 358 с.
Плотников В.В. Эволюция структуры растительных сообществ. М.: Наука, 1979. 276 с.
Проскуряков М.А. Горизонтальная структура горных темнохвойных лесов. Алма-Ата: Наука, 1983. 216 с.
Стороженко В.Г. Устойчивые лесные сообщества. Теория и эксперимент. Тула: Гриф и К, 2007. 192 с.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Факторы регуляции экосистем еловых лесов / Под ред. В.Г. Карпова. Л.: Наука, 1983. 318 с.
Фардеева М.Б., Исламова Г.Р., Чижикова Н.А. Анализ пространственно-возрастной структуры растений на основе информационно-статистических подходов // Ученые записки Казанского гос. университета. Серия: Естественные науки, 2008. Т. 150. Кн. 4. С. 226–240.
Цветков В.Ф. Лесной биогеоценоз. Архангельск: Изв-во АГТУ, 2004. 267 с.
Ярошенко А.Ю., Потапов П.В., Турубанова С.А. Россия. Европейская часть (сев.) Атлас. Малонарушенные лесные территории европейского севера России: методика и результаты картирования малонарушенных лесных территорий Европейского Севера России на основе космических снимков высокого разрешения. М.: Гринпис России, 2001. 75 с.
Baddeley A., Turner R. Spatstat: an R package for analyzing spatial point patterns // J. Statistical Software. 2005. V. 12. № 6. P. 1–42.
Grabarnik P., Myllymäki M., Stoyan D. Correct testing of mark independence for marked point patterns // Ecological Modelling. 2011. V. 222. № 23–24. P. 3888–3894.
Illian J., Penttinen A., Stoyan H., Stoyan D. Statistical analysis and modelling of spatial point patterns. Chichester: John Wiley & Sons, 2008, 560 p.
Schabenberger O., Gotway C.A. Statistical methods for spatial data analysis. Boca Raton: Chapman & Hall/CRC, 2005. 505 p.
Stoyan D., Penttinen A. Recent applications of point process methods in forestry statistics // Statistical Science. 2000. V. 15. № 1. P. 61–78.
Wiegand T., Moloney K.A. Rings, circles, and null-models for point pattern analysis in ecology // Oikos. 2004. V. 104. № 2. P. 209–229.
Дополнительные материалы отсутствуют.