

Лесоведение, 2019, № 5, стр. 366-374
Динамика популяции непарного шелкопряда в лесостепных дубравах Европейской России
Н. И. Лямцев *
ВНИИ лесоводства и механизации лесного хозяйства
141200 Московская обл, Пушкино, ул. Институтская, 15, Россия
* E-mail: nilyamcev@yandex.ru
Поступила в редакцию 04.02.2019
После доработки 04.02.2019
Принята к публикации 05.06.2019
Аннотация
Приведены результаты стационарных исследований изменения численности непарного шелкопряда (Lymantria dispar L.) в 1977–2018 гг., характеризующие пять вспышек массового размножения. На основе анализа их фазовых портретов, а также эколого-популяционных показателей фаз выделены эруптивные и условно продромальные градационные циклы. Особенностью динамики численности непарного шелкопряда в порослевых дубравах юго-востока Европейской России является перманентный характер вспышек массового размножения. В оптимальных экологических условиях (засушливый климат, ослабленные, простые по структуре насаждения) популяция непарного шелкопряда характеризуется более высоким средним уровнем численности, меньшими максимальной плотностью популяции и амплитудой колебаний, значительно более высоким уровнем минимальной плотности популяции, отсутствием длительных межвспышечных периодов. Периодичность массовых размножений составляет 10 лет. Для колебаний численности характерно чередование циклов разной интенсивности. После эруптивного этапа в многолетней динамике численности наблюдалась менее интенсивная вспышка массового размножения (условно продромальный этап). Его отличие от типичной продромальной градации – восстановление численности (прекращение разреживания популяции) на более высоком уровне, чем стабильная численность, что сразу приводит к ее интенсивному росту и новому массовому размножению. Причиной является недостаточно эффективный комплекс многоядных энтомофагов, регулирующих популяцию непарного шелкопряда в интервале от низкой до близкой к средней численности.
Непарный шелкопряд (Lymantria dispar L.) – один из наиболее распространенных и хозяйственно опасных вредителей леса. Очаги его массового размножения периодически возникают и приводят к повреждению лесов на большой территории. Способность к крупномасштабным вспышкам численности обусловлена значительной изменчивостью популяционных параметров, высокой плодовитостью, полифагией, полиморфизмом, повышенной миграционной активностью, большой инерционностью биоценотических механизмов регуляции (Исаев и др., 2001).
Динамика численности непарного шелкопряда определяется комплексом факторов, для которых характерна существенная географическая изменчивость. Известно, что экологические условия лесостепной зоны наиболее оптимальны для развития непарного шелкопряда, как тепло- и сухолюбивого вида (Ильинский, 1959; Лямцев и др., 2000). Очаги насекомого наблюдаются здесь практически ежегодно, а массовые размножения – с определенной периодичностью.
Не смотря на большое количество исследований (Ханисламов и др., 1958; Ильинский, 1959; Воронцов, 1977; Рафес, 1980; Рубцов, Рубцова, 1980; Семевский, Семенов, 1982; Бенкевич, 1984; Знаменский, 1984; Лямцев, Дмитриева, 1998; Пономарев и др., 2012), многие вопросы экологии непарного шелкопряда изучены недостаточно. Для их решения необходим системный подход и длительные комплексные наблюдения. Целью настоящей работы является анализ результатов многолетних исследований популяционной динамики непарного шелкопряда на стационарных участках для выявления закономерностей его массовых размножений.
МАТЕРИАЛ И МЕТОДИКА
Экспериментальные данные по изменению численности непарного шелкопряда и других эколого-популяционных показателей получены в 1977–2018 гг. в порослевых дубравах Базарно-Карабулакского лесхоза (лесничества) Саратовской обл. Они находятся на Приволжской возвышенности в юго-восточной части европейской лесостепи. Дуб произрастает совместно с липой и кленом остролистным. Дубравы подвержены интенсивному антропогенному воздействию. Преобладают порослевые, одноярусные, с небольшим количеством подроста и подлеска, биологически малоустойчивые древостои. Насаждения характеризуются значительной изменчивостью структуры и условий произрастания, что обуcловлено изрезанностью рельефа, обилием склонов разной крутизны и экспозиции. Возраст насаждений на начало исследований варьировал от 20 до 90 лет, полнота – от 0.3 до 0.9, класс бонитета – от II до V.
Плотность популяции непарного шелкопряда на начальном этапе работ оценивали на 30 постоянных пробных площадях, затем их количество сократилось до 16. Кладки яиц учитывали ежегодно, осматривая на каждой пробной площади комлевые части стволов не менее 50 деревьев при высокой численности насекомого, а при низкой – не менее 100 деревьев, а также подрост и подлесок в пределах проекции крон.
Другие популяционные показатели на постоянных участках оценивали методом случайной выборки. Расчет оптимального размера учетных единиц, схемы их размещения и необходимой величины выборки для получения достоверных данных проводили с учетом характера варьирования параметров популяции (Знаменский, Белов, 1976; Белов, 1978; Панина, 1979; Знаменский и др., 1981; Куприянова, 1986).
Учеты проводили на стволах деревьев (кладки яиц и гусеницы), в кроне (гусеницы и куколки), искусственных укрытиях (гусеницы и куколки). Для оценки экологической плотности популяции численность непарного шелкопряда переводили на единицу пищевого субстрата, в качестве которой использовали побег текущего года (точка роста).
Пробные площади расположены в разных по породному составу, структуре и состоянию насаждениях, отличающихся по степени пригодности для развития непарного шелкопряда. В Саратовской обл. в большей степени повреждаются средневозрастные (30–50 лет), низкобонитетные, одноярусные порослевые дубравы с полнотой 0.4–0.7, имеющие в составе не более 30–40% других пород. Такие насаждения являются резервациями и первичными очагами вредителя (Знаменский, Лямцев, 1990). Они произрастают преимущественно на склонах южной экспозиции в неблагоприятных почвенно-гидрологических условиях и характеризуются максимальной степенью ослабления.
Дуб является предпочитаемой кормовой породой. Однако при высокой численности гусеницы непарного шелкопряда питаются на липе, осине, березе и др. При увеличении доли дуба в составе насаждений их заселенность обычно увеличивается. Наиболее четко это проявляется при низкой численности насекомых, когда доля предпочитаемой кормовой породы в составе насаждений объясняла до 50% варьирования плотности популяции.
Состояние насаждений существенно варьирует во времени. Основными факторами динамики состояния являются дефолиация насекомыми и засухи. Степень засушливости климата является также важным модифицирующим фактором динамики численности непарного шелкопряда, существенно влияющим на частоту образования очагов. Для характеристики засушливости использовали количество осадков в весенне-летний период и гидротермический коэффициент (ГТК) Селянинова (Лямцев и др., 2000). Число лет с засухами в мае–июне и в мае–июле составляет соответственно 45 и 35% всего периода наблюдений. При таких засухах гидротермический коэффициент меньше 0.8. Доля лет с недостаточным увлажнением в весенне-летний период (ГТК < 1.0) составляет около 50% (Лямцев, 2013). Анализ радиального прироста дуба как интегрального показателя состояния деревьев показал, что массовые размножения насекомых начинаются после достаточно сильного падения прироста, свидетельствующего о снижении устойчивости насаждений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В течение 45-летнего периода нами проведены ежегодные учеты численности и мониторинг 5 вспышек массового размножения непарного шелкопряда. Многолетняя динамика плотности популяции насекомого приведена в табл. 1.
Таблица 1.
Среднее число кладок яиц непарного шелкопряда (шт. на 1 дерево) осенью 1976–2018 гг.
| Год | Учтено деревьев |
Кладок яиц | Год | Учтено деревьев |
Кладок яиц | Год | Учтено деревьев |
Кладок яиц |
|---|---|---|---|---|---|---|---|---|
| 1976 | 2000 | 3.364 | 1991 | 3200 | 0.025 | 2006 | 1500 | 0.785 |
| 1977 | 3556 | 1.516 | 1992 | 3200 | 0.054 | 2007 | 1300 | 1.485 |
| 1978 | 9012 | 0.525 | 1993 | 1600 | 0.142 | 2008 | 1300 | 0.451 |
| 1979 | 9400 | 0.397 | 1994 | 1600 | 1.322 | 2009 | 1600 | 0.018 |
| 1980 | 8846 | 0.084 | 1995 | 1600 | 4.769 | 2010 | 1600 | 0.003 |
| 1981 | 11 600 | 0.009 | 1996 | 1400 | 0.650 | 2011 | 1600 | 0.004 |
| 1982 | 12 010 | 0.012 | 1997 | 1550 | 0.220 | 2012 | 1600 | 0.057 |
| 1983 | 12 200 | 0.021 | 1998 | 1550 | 0.198 | 2013 | 1400 | 0.646 |
| 1984 | 12 400 | 0.018 | 1999 | 1800 | 0.028 | 2014 | 800 | 1.125 |
| 1985 | 11 800 | 0.046 | 2000 | 1600 | 0.038 | 2015 | 1600 | 0.538 |
| 1986 | 5900 | 0.270 | 2001 | 1600 | 0.010 | 2016 | 1600 | 0.092 |
| 1987 | 4142 | 0.613 | 2002 | 1600 | 0.007 | 2017 | 1600 | 0.015 |
| 1988 | 3100 | 0.663 | 2003 | 1600 | 0.013 | 2018 | 2800 | 0.006 |
| 1989 | 3100 | 0.284 | 2004 | 1600 | 0.030 | |||
| 1990 | 3100 | 0.057 | 2005 | 1600 | 0.160 |
Временной ряд (табл. 1) можно увеличить, добавив оценки численности непарного шелкопряда в этих насаждениях (Знаменский, Лямцев, 1990) в 1973–1975 гг. (соответственно 0.08, 0.52, 2.20 кладок яиц на дерево). Однако необходимо учитывать, что они получены не на стационарных участках.
Вспышки массового размножения непарного шелкопряда (табл. 1) имели как общие черты, так и различия. Три из них (2004–2011 и 2012–2018 гг., а особенно 1985–1992 гг.) отличались существенно меньшей интенсивностью. Две других (1973–1981 и 1993–2001 гг.) были типично эруптивного типа (по классификации А.С. Исаева с соавторами (2001).
В целом для популяционной динамики характерно отсутствие длительных межвспышечных периодов и интенсивного снижения численности непарного шелкопряда, сравнительно невысокие размах колебания плотности популяции и ее максимальная величина, небольшая величина зоны стабильной численности. Это свидетельствует о перманентном характере массовых размножений непарного шелкопряда в дубравах южной части лесостепи (Лямцев, Исаев, 2005). Благодаря более высокой эффективности внутрипопуляционных регуляторных механизмов во время массового размножения насаждения в сильной степени повреждаются в основном один раз.
Наименее интенсивное массовое размножение 1985–1992 гг. можно охарактеризовать как продромальное. Массовые размножения такого типа отличаются от эруптивных на порядок меньшей амплитудой изменения плотности популяции, отсутствием депрессии численности и быстрым ее восстановлением до уровня стабильной плотности популяции (Исаев и др., 2001).
Для выявления закономерностей динамики численности использовали метод фазовых портретов, который широко применяется в популяционной экологии. Выходной величиной системы “популяция насекомых–внешняя среда” является плотность популяции. Изменение этой величины изображается точкой на фазовой плоскости, на которой по оси абсцисс отложена плотность популяции, а по оси ординат – скорость ее изменения (коэффициент размножения). Поскольку плотность популяции меняется во времени, на фазовой плоскости получается определенная траектория. Она имеет более простую и наглядную форму, если использовать логарифмическую шкалу (Семевский, 1972). По виду фазовой траектории многолетней динамики численности насекомого можно судить о характере популяционной динамики. При затухающих колебаниях траектория имеет вид закручивающейся вокруг особой точки спирали. В случае периодических колебаний траектория представлена семейством замкнутых кривых с общим центром в особой точке. При нарастающих колебаниях траектория представляет собой раскручивающуюся из особой точки спираль.
На рис. 1 показана функциональная связь коэффициента размножения с плотностью популяции (кладок на дерево) в начале генерации. Фазовые портреты и все фазы двух отличающихся градаций непарного шелкопряда построены по экспериментальным данным.
Рис. 1.
Фазовая траектория интенсивной (1976–1982 гг.) и менее интенсивной (1983–1992 гг.) вспышек массового размножения непарного шелкопряда в дубравах Базарно-Карабулакского лесничества Саратовской обл. Фазы: bc – нарастания численности, cd – максимума, de – разреживания, eh – депрессии, ha – восстановления стабильной численности, ab – стабилизации; r-yr – пороговая линия.
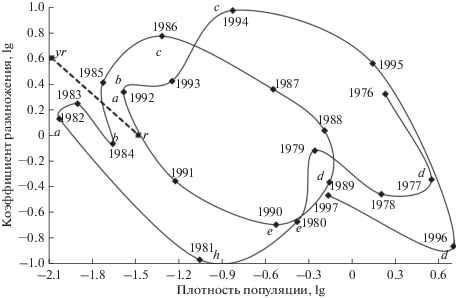
Как видно из рис. 1, массовые размножения закономерно развиваются во времени и проходят пять качественно различных этапов (фаз): нарастания численности, максимума, разреживания, депрессии и восстановления стабильной численности (Исаев и др., 2001). Для всех массовых размножений характерен тождественный характер изменения плотности популяции, симметричное расположение максимумов и минимумов численности относительно прямой у = 0, отсутствие метастабильного состояния. В то же время фазовые портреты различаются по структуре, предельным значениям плотности популяции и коэффициента размножения.
Возникновение вспышек массового размножения непарного шелкопряда связано с засушливой погодой, которая оказывает прямое и косвенное влияние на популяцию, приводит к росту выживаемости и плодовитости насекомого, снижению эффективности природных регулирующих механизмов и устойчивости насаждений. Пороговой численностью непарного шелкопряда является 0.03–0.05 кладок на дерево.
Фаза нарастания численности реализуется в области максимальной инерционности регулирующих механизмов. Отрицательная обратная связь меняется на положительную. Плотность популяции увеличивается наиболее интенсивно, коэффициент размножения на эруптивном этапе динамики численности значительно выше (9.0 в 1994 г.), чем на продромальном (6.3 в 1986 г.). Проявление положительной обратной связи обусловлено ростом выживаемости особей до максимальной величины (3.7–6.2%), высоким уровнем плодовитости (319 яиц). Отмечается небольшое преобладание самок в популяции (половой индекс 0.53) и минимальное воздействие естественных врагов; зараженность куколок не превышает 10% (табл. 2). При переходе популяции через пороговый уровень численности наблюдается максимальная плодовитость (427 яиц в 1984 г.).
Таблица 2.
Изменение параметров популяции непарного шелкопряда по фазам градации в Саратовской обл.
| Популяционный параметр | Изменение параметра по фазам градации | |||
|---|---|---|---|---|
| роста численности | максимума | разреживания | депрессии и восстановления | |
| Коэффициент размножения | $\frac{{1.9 - 9.0}}{{2.3 - 6.3}}$ | $\frac{{3.0 - 1.9}}{{1.9 - 0.9}}$ | $\frac{{0.2 - 0.8}}{{0.4 - 0.18}}$ | $\frac{{0.16 - 1.2}}{{0.44 - 2.2}}$ |
| Число яиц в кладке, шт. | $\frac{{319 - 280}}{{427 - 366}}$ | $\frac{{172}}{{285}}$ | $\frac{{268 - 286}}{{276 - 358}}$ | $\frac{{230 - 370}}{{358}}$ |
| Половой индекс | $\frac{{0.53 - 0.49}}{{0.52 - 0.43}}$ | $\frac{{0.21}}{{0.34}}$ | $\frac{{0.30 - 0.42}}{{0.37 - 0.42}}$ | $\frac{{0.38 - 0.67}}{{0.61}}$ |
| Паразитировано куколок, % | $\frac{{9.0 - 9.4}}{{10 - 5.4}}$ | $\frac{{24.0 - 34.4}}{{54.0 - 53.7}}$ | $\frac{{59.4 - 56.1}}{{48.3 - 37.7}}$ | $\frac{{52.6 - 12.5}}{{10}}$ |
На фазе максимума эруптивной вспышки численность насекомого продолжает расти до предельного значения, которое в Саратовской обл. составляет в среднем около 60 яиц на 100 точек роста и не является пределом, определяемым ресурсами. Максимальная плотность популяции при градации продромального типа в 5 раз меньше. Коэффициент размножения уменьшается (до единицы и несколько меньше). Обратная связь вновь становится отрицательной, а ее проявление обусловлено доминирующим влиянием внутрипопуляционных регуляторных механизмов. Они эффективны в Саратовской обл., где плодовитость (172 яйца) и доля самок (0.21) в популяции к концу фазы максимума эруптивной вспышки (1977 г.) достигают минимальной величины. Увеличивается смертность в популяции, но выживаемость особей еще близка к средней (0.68–0.78%). Зараженность куколок паразитами увеличивается до 34.4% при эруптивном цикле и достигает максимума (54%) при продромальном. Болезни не принимают форму эпизоотий. Возрастает миграционная активность. Непарный шелкопряд занимает разнообразные стации.
На фазе разреживания снижение скорости роста популяции безинерционными регуляторными механизмами способствует повышению эффективности биоценотических факторов. Происходит дальнейшее снижение численности и коэффициента размножения непарного шелкопряда. При возвращении системы в область все возрастающего воздействия инерционных механизмов регуляции реализуется фаза разреживания. Обратная связь становится положительной. Выживаемость особей за генерацию составляет 0.31–0.42%. Зараженность предкуколок и куколок паразитическими двукрылыми в конце фазы достигает максимума (59.4%) при эруптивном цикле и начинает снижаться (до 37.7%) при продромальном цикле. Внутрипопуляционные механизмы регуляции продолжают действовать, но менее интенсивно, особенно при градации продромального типа. Величина плодовитости (268 яиц) и доля самок (0.3) в популяции ниже среднего, хотя и увеличиваются по мере снижения плотности популяции до 358 яиц и 0.42 соответственно. Фаза разреживания популяции начинает формироваться еще в области действия внутрипопуляционных регуляторов, так как год кульминации массового размножения одновременно является и началом его кризиса.
Фаза депрессии в Саратовской обл. характеризуется наиболее низкими коэффициентами размножения (0.16) и выживаемостью за генерацию (0.07%), все еще значительным воздействием естественных врагов (52.6% паразитированных куколок при эруптивном цикле). В конце фазы депрессии происходит резкое изменение структуры популяции и ее оздоровление, что связано, по-видимому, с избирательной смертностью. Существенно увеличивается плодовитость (до 370 яиц) и доля самок в популяции (до 0.67), смертность куколок снижается в 4 раза (до 12.5%). Это свидетельствует об усилении внутрипопуляционной регуляции (появлении отрицательной обратной связи), которая компенсирует снизившуюся, но все еще избыточную смертность от энтомофагов и при эруптивном градационном цикле возвращает популяцию к устойчивому состоянию. Этот этап характеризуется фазой восстановления стабильной численности. Снижение плотности популяции прекращается, коэффициент размножения увеличивается до 1. Затем (1983–1984 гг.) популяция находится в фазе стабильной численности (рис. 1).
Менее высокие темпы прироста численности непарного шелкопряда и ее более низкий уровень при продромальном массовом размножении (1985–1992 гг.) повысили эффективность специализированных энтомофагов как регуляторов численности (рис. 2). Их реакция на изменение плотности популяции хозяина в такой ситуации характеризуется минимальной величиной запаздывания и становится положительной. При увеличении численности непарного шелкопряда его смертность от паразита также увеличивается, происходит регулирование популяции. Противоположная ситуация была при эруптивном массовом размножении 1976–1982 гг., когда при интенсивном росте плотности популяции непарного шелкопряда специализированный паразит снижал, но не регулировал его численность (рис. 2).
Рис. 2.
Зависимость зараженности гусениц непарного шелкопряда личинками специализированного паразита (тахина Parasetigena silvestris R.-D.) от плотности его популяции в начале генерации.
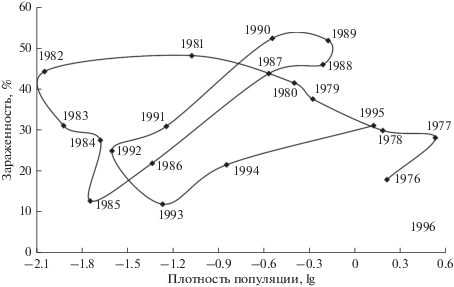
Однако уровень смертности от специализированных паразитов оказался недостаточным. Действие внутрипопуляционных регуляторов также было менее значимо. Поэтому быстрое разреживание популяции (1989–1990 гг.) не перешло в фазу депрессии, она практически отсутствовала. Восстановление стабильной численности (1991–1992 гг.) произошла на более высоком ее уровне. В связи с этим после завершения продромального градационного цикла популяция не попадает в зону стабильной численности (не стабилизирует численность на низком уровне), а сразу переходит к новой вспышке массового размножения. Реализация массового размножения после менее интенсивной вспышки продромального типа приводит к интенсивному нарастанию численности насекомого. В динамике популяции проявляются черты бумеранг-эффекта.
Таким образом, в дубравах Саратовской обл. в многолетней динамике популяции непарного шелкопряда этапы эруптивного типа чередуются с этапами продромального типа. Это уменьшает риск дестабилизации лесных экосистем, непарный шелкопряд даже при интенсивной вспышке не повреждает дуб в сильной степени два года подряд, и его популяция находится в режиме перманентной вспышки.
В отличие от североамериканской популяции непарного шелкопряда, на юго-востоке европейской России не наблюдались стабилизация численности на высоком уровне и образование хронических очагов (Знаменский, Лямцев, 1990), хотя такая попытка имела место в 1979 г. (рис. 1). Однако эффективный на этом уровне комплекс специализированных энтомофагов, а также ухудшение качества корма и структуры популяции после дефолиации привели к дальнейшему снижению численности. Наличие комплекса эффективных внутрипопуляционных и биоценотических (специализированные паразиты) регуляторов, которые существенно сокращают период сильных повреждений дубрав, являются характерной особенностью этой популяции.
Массовое размножение 1985–1992 гг. оказалось наиболее близким к продромальному типу. Дефолиация насаждений и площадь очагов непарного шелкопряда были меньше (Знаменский, Лямцев, 1990). Особенностью цикла являются невысокие амплитуда и темпы роста численности, реализация массового размножения после эруптивной вспышки и достаточно длинного лаг-периода (1982–1985 гг.), отсутствие выраженной депрессии. Это типичные критерии продромального цикла (Исаев и др., 2001). Однако имеются и различия. Первый год восстановления численности является одновременно и годом депрессии: они совпадают. Главное отличие заключается в том, что разреживание популяции прекращается и ее восстановление происходит не в зоне стабильной плотности, а на более высоком уровне. Популяция не стабилизируется, и сразу начинается фаза нарастания численности нового массового размножения. Поэтому такой градационный цикл можно считать только условно продромальным. В нем имеются черты нарушенного (прерванного) массового размножения эруптивного типа.
Реализация продромального цикла динамики численности непарного шелкопряда после эруптивного может быть обусловлена рядом факторов. После интенсивной вспышки ухудшаются качество корма и структура популяции насекомого. В результате она задерживается в зоне стабильности (1982–1984 гг.), и темпы прироста численности снижаются. Эти причины, а также факторы, имеющие циклический характер, например солнечная активность (Лямцев, Дмитриева; 1998), могут вносить определенную периодичность в чередование интенсивных и менее интенсивных вспышек. Однако этот вопрос еще недостаточно изучен, а имеющиеся немногочисленные сведения основаны на анализе площадей очагов и свидетельствуют лишь о существовании такого явления.
Наиболее вероятно развитие вспышки по продромальному типу в результате длительного, на протяжении ряда лет, действия фактора, уменьшающего количество корма. Так, в 1983–1989 гг. наблюдалось размножение комплекса листогрызуших насекомых, среди которых доминировала зеленая дубовая листовертка. Она оказалась более конкурентноспособной и по мере роста численности популяций вытеснила непарный шелкопряд в менее пригодные для развития местообитания (Знаменский, Лямцев, 1990).
При максимальном насыщении местообитаний (1988–1989 гг.) плотность популяции листовертки была в 50 раз выше, чем непарного шелкопряда. Плотность популяции шелкопряда составила лишь 20% от наблюдавшейся в предыдущую вспышку (1976 г.). Гусеницы непарного шелкопряда были вынуждены концентрироваться в нижней, еще не поврежденной части кроны, в смешанных более высокополнотных насаждениях или частично питались вторичной листвой. Поэтому весной 1989 г., когда численность листовертки достигла пика, плотность популяции непарного шелкопряда в менее благоприятных для его развития насаждениях (14.02 яиц на 100 точек роста) была существенно (на 40%) выше, чем в благоприятных условиях (9.85 яиц на 100 точек роста). На такой же фазе предыдущей градации заселенность шелкопрядом благоприятных для развития насаждений (64.79 яиц на 100 точек роста), наоборот, была в 1.46 раза выше, чем менее благоприятных (44.47 яиц на 100 точек роста) (Лямцев, 2013).
Ухудшение трофических и микроклиматических условий способствовали снижению выживаемости и коэффициента размножения непарного шелкопряда в комплексных очагах. Кроме того, в комплексных очагах при максимуме вспышки характерно преимущественное сосредоточение популяции непарного шелкопряда в смешанных насаждениях и высокая его зараженность специализированными паразитами.
Проведенный анализ и многолетние данные позволяют, прежде всего, оценить продолжительность градационного цикла и отдельных фаз. Это важный итог, так как вследствие значительной изменчивости массовых размножений (табл. 3) он может быть получен только в результате длительных наблюдений.
Таблица 3.
Продолжительность массовых размножений непарного шелкопряда и их фаз за период с 1973 по 2018 г.
| Фаза градационного цикла | Продолжительность фаз (лет) для градаций | |||||
|---|---|---|---|---|---|---|
| 1973–1984 | 1985–1992 | 1993–2003 | 2004–2011 | 2012–2018 | Средняя (мода) | |
| Нарастания | 2 | 2 | 2 | 2 | 2 | 2 |
| Максимума | 2 | 2 | 1 | 2 | 1 | 2 |
| Разреживания | 4 | 2 | 3 | 2 | 3 | 3 |
| Депрессии | 1 | 0 | 1 | 1 | 1 | 1 |
| Восстановления | 1 | 2 | 1 | 1 | – | 1 |
| Стабилизации | 2 | 0 | 3 | 0 | – | 1 |
| Продолжительность градации | 12 | 8 | 11 | 8 | – | 10 |
Как видно из табл. 3, фаза нарастания численности наименее изменчива и продолжается 2 года. Длительность фаза максимума – 1–2 года. Наиболее продолжительной является фаза разреживания (2–4 года). Для всех массовых размножений характерно отсутствие глубокой депрессии (1 год), быстрое восстановление стабильной численности (1–2 года). Наиболее изменчивым и специфичным является этап стабилизации. Он длительный (3 года) после наиболее интенсивной вспышки (плотность популяции 4.8 кладок на дерево в 1995 г.). После продромального цикла (менее интенсивного массового размножения) стабилизации популяции не наблюдается, а сразу начинается ее рост. Переход популяции в фазу нарастания численности в ослабленных дубравах юго-востока Европейской России в большей степени определяется не модифицирующим воздействием погоды (она, как правило, благоприятная), а текущей эффективностью биоценотических регуляторов (энтомофагов). Накопление энтомофагов, более значительное запаздывание их реакции и более глубокая депрессия после эруптивных вспышек являются одной из причин увеличения продолжительности периода стабилизации численности непарного шелкопряда.
Средняя продолжительность градационного цикла непарного шелкопряда в порослевых дубравах (табл. 3) составляет 10 лет. Периодичность массовых размножений, установленная по колебаниям площадей очагов в Саратовской обл., в 1954–1994 гг. в среднем также равна 10 годам, а в период 1944–1994 гг. – 12.5 лет. Это свидетельствует, что частота вспышек массового размножения может меняться.
При интенсивной вспышке массового размножения плотность популяции колебалась в течение градации от 0.009 до 4.8 кладок на дерево при наименее интенсивной вспышке – от 0.023 до 0.663 кладок на дерево. При высокой численности плотность популяции в разных насаждениях колебалась от 0.064 до 8.3 кладок на дерево, при низкой – от 0 до 0.115 кладок на дерево. При продромальном массовом размножении на фазе максимума в разных насаждениях плотность популяции варьировала от 0.2 до 2.1 кладок на дерево. О высокой изменчивости численности насекомого свидетельствует антилогарифм утроенного стандартного отклонения логарифма плотности популяции (antilog 3σ), равный 820.
С интенсивностью колебаний плотности популяции непарного шелкопряда и фазами градации были связаны абсолютные значения и направление изменения плодовитости, коэффициента асимметрии распределения кладок по числу яиц, полового индекса, средней массы яйца в кладке и других эколого-популяционных показателей (Знаменский, Лямцев, 1985; Лямцев, 2013). В комплексных очагах (менее интенсивная вспышка) размах их варьирования был гораздо меньше (табл. 2). Однако направление и относительное изменение величин указанных показателей остаются такими же, как и при более интенсивной вспышке в 70-ые годы, что позволяет использовать их для диагностики и прогнозирования фаз массового размножения непарного шелкопряда.
Заключение. Результаты многолетних стационарных исследований позволяют существенно уточнить эколого-популяционные показатели фаз и типы вспышек массового размножения непарного шелкопряда. Интегральной характеристикой динамики популяций насекомого являются параметры фазовых портретов изменения численности. Особенностью динамики численности непарного шелкопряда в дубравах юго-востока Европейской России является перманентный характер вспышек массового размножения. В оптимальных экологических условиях (засушливый климат, ослабленные, простые по структуре насаждения) популяция непарного шелкопряда характеризуется более высоким средним уровнем численности, меньшими максимальной плотностью популяции и амплитудой колебаний, значительно более высоким уровнем минимальной плотности популяции, отсутствием длительных межвспышечных периодов (депрессии). Продолжительность градационных циклов составляет 8–11 лет, а их периодичность в среднем 10 лет. Для колебаний численности характерно чередование циклов разной интенсивности. После эруптивного этапа в многолетней динамике численности наблюдалось менее интенсивное массовое размножение. От классического продромального оно отличалось тем, что практически не было периода депрессии и необходимого разреживания популяции до стабильного уровня. Переход к росту численности на более высоком уровне плотности популяции, чем стабильный, приводит к эруптивной вспышке массового размножения.
Список литературы
Белов А.Н. Размер выборки при учете кладок яиц непарного шелкопряда в дубовых древостоях // Лесоведение. 1978. № 3. С. 77–83.
Бенкевич В.И. Массовые появления непарного шелкопряда в европейской части СССР. М.: Наука, 1984. 143 с.
Воронцов А.И. Некоторые итоги изучения непарного шелкопряда // Насекомые вредители лесов Башкирии. Уфа: БФАН СССР, 1977. С. 3–25.
Знаменский В.С. Динамика численности непарного шелкопряда в дубравах лесостепи // Лесоведение. 1984. № 4. С. 12–20.
Знаменский В.С., Белов А.Н. Оптимизация системы учета гусениц и куколок непарного шелкопряда // Экспресс-информация. М.: ЦБНТИлесхоз, 1976. Вып. II. С. 14–15.
Знаменский В.С., Лямцев Н.И., Полякова Л.А. Метод учета численности непарного шелкопряда в разреженных популяциях // Лесное хоз-во. 1981. № 6. С. 49–51.
Знаменский В.С., Лямцев Н.И. Индикаторы массового размножения непарного шелкопряда // Лесное хоз-во. 1985. № 2. С. 60–62.
Знаменский В.С., Лямцев Н.И. Особенности динамики численности непарного шелкопряда в комплексных очагах листогрызущих насекомых // Защита леса от вредных насекомых и болезней: Сб. трудов. М.: ВНИИ лесоводства и механизации, 1990. С. 11–21.
Ильинский А.И. Непарный шелкопряд и меры борьбы с ним. М.: Гослесбумиздат, 1959. 63 с.
Исаев А.С., Хлебопрос Р.Г., Недорезов Л.В., Кондаков Ю.П., Киселев В.В., Суховольский В.Г. Популяционная динамика лесных насекомых. М.: Наука, 2001. 374 с.
Куприянова В.А. Влияние биопрепаратов на зараженность непарного шелкопряда и златогузки паразитическими насекомыми // Защита леса от вредителей и болезней: сб. тр. М.: ВНИИ лесоводства и механизации, 1986. С. 15–19.
Лямцев Н.И. Динамика численности непарного шелкопряда в лесостепных дубравах Европейской России. Пушкино: ВНИИ лесоводства и механизации, 2013. 98 с.
Лямцев Н.И., Дмитриева И.В. Влияние солнечной активности на изменение численности непарного шелкопряда // Биофизика. 1998. Т. 43. Вып. 4. С. 603–609.
Лямцев Н.И., Исаев А.С. Модификация типов вспышек массового размножения непарного шелкопряда в зависимости от эколого-климатической ситуации // Лесоведение. 2005. № 5. С. 3–9.
Лямцев Н.И., Исаев А.С., Зукерт Н.В. Влияние климата и погоды на динамику численности непарного шелкопряда в европейской России // Лесоведение. 2000. № 1. С. 62–67.
Панина Н.Б. Определение эффективности тахин и саркофагид-энтомофагов непарного шелкопряд // Лесное хоз-во. 1979. № 12. С. 57–59.
Пономарев В.И., Гниненко Ю.И., Ильиных А.В., Соколов Г.И., Андреева Е.М. Непарный шелкопряд в Зауралье и Западной Сибири. Екатеринбург: УрО РАН, 2012. 321 с.
Рафес П.М. Биогеоценологические исследования растительноядных лесных насекомых. М.: Наука, 1980. 168 с.
Рубцов В.В., Рубцова Н.Н. Анализ взаимодействия листогрызущих насекомых с дубом. М.: Наука, 1984. 184 с.
Семевский Ф.Н. Оценка регуляторной роли факторов динамики численности // Журн. общей биологи. 1972. Т. 33. № 5. С. 569–578.
Семевский Ф.Н., Семенов С.М. Математическое моделирование экологических процессов. Л.: Гидрометеоиздат, 1982. 280 с.
Ханисламов М.Г., Гирфанова Л.Н., Яфаева З.Ш., Степанова Р.К. Массовые размножения непарного шелкопряда в Башкирии // Исследование очагов вредителей леса Башкирии. Уфа: БФАН СССР, 1958. Т. 1. С. 5–45.
Дополнительные материалы отсутствуют.