

Лесоведение, 2019, № 5, стр. 375-384
Реакция лесных насекомых-филлофагов на современное изменение климата
В. В. Рубцов a, *, И. А. Уткина a
a Институт лесоведения Российской академии наук
143030 Московская обл., Успенское, ул. Советская, 21, Россия
* E-mail: VRubtsov@mail.ru
Поступила в редакцию 29.03.2019
После доработки 28.04.2019
Принята к публикации 05.06.2019
Аннотация
Особенности современного климата оказывают большое влияние на растения и насекомых и их биоценотические взаимосвязи. Это однозначно подтверждают выполненные исследования во многих странах мира, краткий обзор которых приведен в статье. Большинство авторов отмечают, что температура воздуха является самым важным фактором, непосредственно влияющим на развитие и численность популяций насекомых. Зафиксированы изменения ареалов многих видов растений и насекомых в виде сдвигов в северном направлении и вверх по высоте над уровнем моря, более раннее наступление весенних фенофаз и более позднее – осенних, отчего происходит удлинение вегетационного периода. Выполнен анализ временных рядов и выявлены тренды основных метеоэлементов на юге лесостепной зоны (Воронежская обл.). Показано уменьшение континентальности климата, увеличение количества осадков за вегетацию, возрастание частоты периодов с аномальными погодными ситуациями. Зафиксированы заметные изменения сложившихся в данных условиях местообитания циклов динамики численности популяций насекомых, нарушения баланса их взаимоотношений с кормовыми растениями, в частности, непарного шелкопряда, зеленой дубовой листовертки и зимней пяденицы. Происходит уменьшение вредоносности одних видов и увеличение – других. В настоящее время с помощью моделирования можно прогнозировать некоторые последствия прямого воздействия изменения климата на взаимоотношения насекомых-филлофагов с кормовыми растениями. Отдаленные последствия, связанные с косвенным воздействием климата, предвидеть сейчас крайне сложно. Они будут зависеть, с одной стороны, от характера изменений климата, с другой стороны – от реакции на это растений и насекомых, обусловленной их адаптационными механизмами.
Доказательства того, что происходящие изменения климата оказывают заметное влияние на природные системы, появляются на протяжении последних десятилетий, как в конце прошлого века, так и в начале нынешнего. В одном часто упоминаемом обзоре по этой проблеме обобщены данные 228 публикаций о том, что в наземных природных сообществах происходят изменения фенологии и растений, и животных – ареалы тех и других смещаются к северу или вверх по высоте; происходят нарушения в системах “хищник–жертва” и “растение–фитофаги” вследствие разных реакций видов на потепление; в пределах смещающихся видовых ареалов происходит эволюционная адаптация видов к новым условиям, используются новые пищевые ресурсы (Parmesan, 2006).
Влияние происходящих изменений климата на взаимоотношения между насекомыми-фитофагами и лесными породами также зафиксировано во множестве исследований. К их обзорам, появившимся на рубеже веков (Harrington et al., 1999; Hudges, 2000; McCarty, 2001; Root et al., 2003; и др.) добавляются все новые (Мусолин, Саулич, 2012; Jaworski, Hilszczański, 2013; Heimonen et al., 2015; Kolb et al., 2016; Pureswaran et al., 2018; и др.).
Ранее мы писали об основных выводах из этих обзорных работ (Рубцов, Уткина, 2008, 2010; Уткина, Рубцов, 2017; и др.). Здесь отметим, что публикации последних лет подтверждают уже сделанные выводы, свидетельствуют о сложности и указывают на то, что происходящие изменения по-разному влияют на виды насекомых из разных функциональных групп (Jaworski, Hilszczański, 2013), что до сих пор остается неясным, как прогнозируемое изменение количества и частоты осадков и перераспределение на фоне повышения температуры воздуха повлияет на лесных насекомых и возбудителей болезней (Kolb et al., 2016). Еще более непредсказуемо при меняющихся внешних условиях поведение инвазивных видов, способных нанести огромный вред природным сообществам, в которых они оказались (Ramsfield et al., 2016).
О том, что нарушение фенологической синхронности между насекомыми и их кормовыми породами в результате потепления имеет важные экологические последствия, также накоплено немало данных. В лесах умеренной и бореальной зон время начала весеннего развития растений и питающихся ими филлофагов играет ключевую роль, как показано на примере дуба и зимней пяденицы (Buse, Good, 1996; и др.), тополя осиновидного, березы бумажной и кольчатого шелкопряда (Schwartzberg et al., 2014).
А.С. Исаев уделял много внимания проблеме изменения взаимоотношений разных видов насекомых-фитофагов и их кормовых пород под действием климата. Это, например, сосновая пяденица в сосновых лесах Сибири (Исаев и др., 1997, 1999; и др.) и непарный шелкопряд в дубовых и смешанных лесах Европейской России (Лямцев и др., 2000; Лямцев, Исаев, 2005; и др.). “Будущие климатические изменения могут существенно изменить характер взаимодействий в системе “лес–насекомые”, привести к учащению вспышек массового размножения вредителей и росту наносимого ими ущерба. В связи с опасностью значительного увеличения повреждаемости лесов вследствие потепления климата важно оценить степень влияния этих изменений на динамику численности насекомых и масштабы их воздействия на лесные экосистемы” (Исаев и др., 1999, с. 39).
А.С. Исаев придавал большое значение методу математического моделирования этих взаимоотношений для прогнозирования возможных последствий, хотя и понимал его ограничения: “Модели, описывающие взаимодействие “дерево–насекомые”, не дают однозначного ответа на вопрос: что произойдет с насаждениями и популяциями насекомых при возможных глобальных изменениях климата. В ответ на климатические изменения может произойти как значительное увеличение поврежденности лесов насекомыми, так и уменьшение их воздействия на лесные экосистемы. Для более точной оценки последствий возможных климатических изменений необходима детальная информация о параметрах взаимодействия “дерево–насекомые” для конкретных видов вредителей и их кормовых пород, а также знание эколого-физиологических реакций насекомых и деревьев в ответ на изменение температуры и влажности” (Исаев и др., 1999, с. 43).
Такое же заключение сделано через 20 лет, на примере двух видов филлофагов с близкими характеристиками: зимней пяденицы и пяденицы-обдирало, обитающими в дубовых насаждениях Германии (Hittenbeck et al., 2019). Верификация модели, построенной авторами для обоих видов сразу, показала, что необходимо собрать дополнительные данные и разработать модели для каждого вида отдельно.
Цель настоящей работы – показать состояние исследований вопроса взаимоотношений лесных насекомых-филлофагов с кормовыми растениями при наблюдающихся изменениях климата на примере дубовых насаждений и насекомых-филлофагов на юге лесостепной зоны (Воронежская обл., Теллермановская дубрава), дать общую количественную оценку изменениям основных метеорологических показателей в районе исследований, показать влияние современной климатической ситуации на цикличность массовых размножений филлофагов в последние десятилетия.
ОБЪЕКТЫ И МЕТОДИКА
Экспериментальные данные по динамике численности популяций основных филлофагов: зеленой дубовой листовертки (Tortrix viridana L.), зимней пяденицы (Operophtera brumata L.), непарного шелкопряда (Lymantria dispar L.) получены в процессе ежегодных длительных наблюдений в разных типах дубрав на территории Теллермановского опытного лесничества – филиала Института лесоведения РАН (ТОЛ ИЛАН РАН, далее ТОЛ), расположенного на востоке Воронежской обл.
Ежегодно все насаждения лесничества обследовались для выявления очагов повреждения листвы. Степень дефолиации крон определяли глазомерно с помощью бинокля, а для деревьев, на которых велись учеты гусениц и изучались особенности восстановления листвы после дефолиаций – расчетным путем. Состояние деревьев и их крон определяли с помощью перечетов на постоянных и временных пробных площадях, а также на маркированных заранее деревьях (около 900 экз.) различных феноформ дуба черешчатого и на модельных деревьях, дополнительно выбранных в очагах сильного повреждения листвы. Учеты яйцекладок зеленой дубовой листовертки проводили по комбинированной методике Н.Н. Егорова и др. (1953) и В.А. Ефремовой (1969). Учет непарного шелкопряда проводили ежегодно по яйцекладкам, осматривая нижние части стволов деревьев в основных типах леса. Зимнюю пяденицу в стадии гусениц учитывали в кронах вместе с другими филлофагами, а также с помощью солидольных ловчих колец в периоды лёта бабочек; плодовитость самок определяли путем вскрытия и подсчета яиц в них (Рубцов, Уткина, 2011).
Анализ многолетней динамики метеоэлементов и выявление их трендов проведены с помощью регрессионных моделей и методов анализа временных рядов. Периоды с аномальными погодными условиями определены с использованием построенных нами климадиаграмм Госсена–Вальтера (Рубцов, Уткина, 2008). Влияние погодных условий на взаимодействие гусениц зеленой дубовой листовертки с листвой оценены с помощью имитационных экспериментов с построенной нами ранее математической моделью (Рубцов, Рубцова, 1984). Основу модели составляет система трех обыкновенных нелинейных дифференциальных уравнений, решаемая численным методом с помощью компьютера.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Представители рода Дуб (Quercus L.) относятся к предпочитаемым кормовым породам для многих насекомых. В частности, на дубе черешчатом (Q. robur L.), одной из лесообразующих древесных пород Европейской части России, обитает более 700 видов насекомых. Различные виды дуба и филлофаги, потребляющие их листву, часто используются как объекты для изучения универсальных механизмов, с помощью которых происходит взаимная адаптация растений и насекомых в разных природных зонах отечественными и зарубежными исследователями (подробнее см. Рубцов, Уткина, 2008; Уткина, Рубцов, 2019).
Климатические факторы оказывают на экосистемы прямое и косвенное воздействие. А.И. Воронцов (1978) отмечал, что при прямом воздействии сложившиеся погодные условия стимулируют или тормозят развитие, рост, питание насекомых и другие биологические процессы. Они же определяют уровень смертности от физических факторов среды (гибель от низких температур, ливней, засухи и т.д.). Очень часто в силу различных требований видов к физическим факторам наблюдается асинхронность в развитии вредителя и его энтомофагов. Косвенное воздействие осуществляется через пищу и местообитания отдельных стадий развития, через энтомофагов и болезни. Оно происходит через сложные цепи взаимодействий в экосистеме и, как правило, более инерционно. Поскольку изменение климата неодинаково в пространстве и времени, то и его влияние на экосистемы различно, что убедительно показано в работах А.С. Исаева (Исаев и др., 1999; Лямцев, Исаев, 2005; и др.).
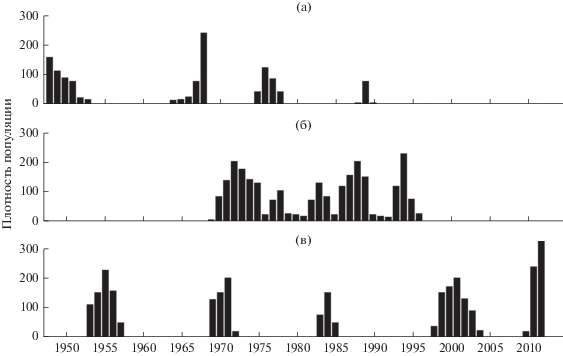
Регулярные наблюдения на территории ТОЛ за массовыми размножениями основных видов насекомых-филлофагов ведутся с момента образования лесничества в 1944 г., с 1975 г. – авторами данного сообщения. За это время зафиксировано четыре вспышки массового размножения непарного шелкопряда, пять вспышек зимней пяденицы. Размножение зеленой дубовой листовертки носило перманентный характер с пятью выраженными максимумами численности популяции.
Анализ длительных рядов основных метеоэлементов в близкорасположенных территориях юга лесостепной зоны: в Хоперском (Бирюков, Марченко, 2001), Воронежском (Венгеров и др., 2001) заповедниках, в Теллермановской дубраве (Рубцов, Уткина, 2008) выявил тенденцию уменьшения континентальности климата в этом регионе. Мы проанализировали временные ряды некоторых климатических показателей по данным метеостанции г. Борисоглебска (Воронежская обл.), которая расположена в восьми километрах от ТОЛ. Установлено, что среднегодовая температура воздуха с 1927 г. к настоящему времени возросла почти на 2°С. Средняя температура воздуха в период вегетации (май–сентябрь) немного понизилась. При этом сильно возросло количество осадков за вегетационный период – на 99 мм с 1949 г. С 1949 по 1975 гг. не было ни одного года с суммарным количеством осадков за вегетацию более 300 мм, а с 1976 по 2009 гг. таких сезонов было 11, в том числе четыре сезона с количеством осадков за вегетацию более 400 мм. Анализ построенных климадиаграмм Госсена-Вальтера показал тенденцию значительного увеличения в последние десятилетия количества периодов вегетации без засушливости, что очень существенно и благоприятно для зоны недостаточного увлажнения.
Наблюдающиеся изменения климата заметно сказались на взаимоотношениях многих видов лесных филлофагов с их кормовыми породами, о чем мы уже писали (Рубцов, Уткина, 2008, 2010; Уткина, Рубцов, 2017, 2019; и др.). Это связано прежде всего с различием в реакциях на происходящие климатические изменения у разных видов растений и филлофагов. В ТОЛ нами зафиксированы существенные изменения в последние десятилетия в динамике численности указанных выше видов филлофагов, заметно снизилась повреждаемость ими листвы в дубравах. Нарушена имевшая место многие десятилетия довольно четкая цикличность вспышек размножения этих насекомых: с 1990 г. до настоящего времени не было массовых размножений непарного шелкопряда (характерный ранее интервал размножений составлял 10–12 лет); в глубокой депрессии более 20 лет находится зеленая дубовая листовертка (до этого плотность популяции имела непрерывные волнообразные подъемы и спады). Предпоследнее массовое размножение зимней пяденицы на территории ТОЛ было затяжным (1997–2004 гг.). Уже в 2009 г., спустя пять лет, началась новая вспышка (ранее межвспышечные интервалы составляли 10–11 лет), которая оказалась очень краткосрочной, с 2009 по 2011 гг., причем в течение ее было два периода с экстремальными погодными условиями – исключительно жаркое лето 2010 г., когда происходило развитие куколок, и зима 2011/2012 гг. с длительным периодом очень низких температур, что привело к почти полной гибели яйцекладок и затуханию вспышки в 2012 г. В период этой краткосрочной вспышки лёт бабочек пяденицы начинался на три недели позже, чем обычно, что напрямую связано с погодными условиями (рис. 1).
Рис. 1.
Динамика численности некоторых насекомых-филлофагов, массово размножавшихся в Теллермановской дубраве в последние 60 лет: а − непарный шелкопряд, число кладок яиц на 50 дер.; б − зеленая дубовая листовертка, число кладок яиц на 5 пог. м ветвей; в − зимняя пяденица, число бабочек-самок на 1 дер.
В это же время на территории ТОЛ необычно сильно размножилась популяция минёра – дубовой широкоминирующей моли (Acrocercops brongniardella L.), вспышка массового размножения которой в Воронежской обл. описана В.Б. Голубом с соавторами (Голуб и др., 2009, 2011). Более 20 лет подряд этот вид повреждает до 60–70% площади поверхности листвы дуба, минируя листья деревьев как ранней, так и поздней феноформ, независимо от их возраста. Степень повреждения каждой формы зависит от погодных условий и наличия других вредителей листвы (Рубцов, Уткина, 2008). Спустя некоторое время листья усыхают и скручиваются (рис. 2).

Во всех известных нам работах отмечается, что наиболее важный внешний фактор, влияющий на развитие и состояние популяций насекомых-фитофагов – температура воздуха. В контексте происходящих климатических изменений это отчетливо показано в обзоре J.S. Bale et al. (2002). Анализируя данные 134 источников, авторы утверждают, что температура – главный абиотический фактор, непосредственно воздействующий на насекомых-фитофагов, а действие концентрации СО2 и увеличения ультрафиолетового излучения намного слабее. Температура прямо влияет на развитие, выживаемость, ареал и численность насекомых. Виды с более обширным ареалом зависят от температуры в меньшей степени (Исаев и др., 1999; Bale et al., 2002; и др.).
Однако реакция насекомых на изменения климата не всегда линейна. Например, фазы развития насекомых по-разному испытывают влияние изменений, т.е. рост может усилиться при повышении температуры воздуха, в то же время продолжительность диапаузы может стать больше. При этом главный эффект температуры в умеренных регионах – влияние на зимнюю выживаемость; в более северных широтах повышение температуры удлиняет вегетационный период, отчего доступная сумма температур для роста и репродукции увеличивается.
Наличие у насекомых-фитофагов разных жизненных стратегий позволяет им использовать в качестве корма растения разных жизненных форм и с различной стратегией, на которых происходящие изменения климата действуют по-разному. В уже упомянутых работах А.С. Исаева с соавторами по прогнозированию влияния изменений климата на динамику численности филлофагов, их ареалов и нанесению вреда древостоям методологическая сторона исследований основана на статистических методах анализа и имитационном моделировании (Исаев и др., 1997; Лямцев, Исаев, 2005; и др.).
Авторы с помощью моделирования детально проанализировали влияние 100 различных климатических сценариев на динамику численности популяции сосновой пяденицы (Bupalus piniarius L.). Были определены критические значения климатических показателей, вызывающих регулярные вспышки ее массового размножения. Установлено, что повышение среднегодовой температуры воздуха на 2–4°С, что, возможно согласно существующим глобальным климатическим сценариям, вызовет в Центральной Сибири потенциальную опасность для сосновых лесов в связи с размножением пяденицы. По мнению авторов, при анализе влияния глобальных изменений климата на динамику численности насекомых следует учитывать не только среднегодичные климатические сдвиги, но и климатические изменения в течение всего года, поскольку развитие популяций в течение сезона зависит от различных климатических факторов. Общий вывод авторов состоит в том, что с потеплением климата вероятность возникновения вспышек массового размножения энтомовредителей может возрастать, а зона их потенциального заселения – расширяться. Вместе с этим констатируется, что в настоящее время нельзя получить однозначный ответ о последствиях глобального изменения климата для динамики численности насекомых и повреждаемости ими лесов. Для этого необходима более детальная информация о взаимоотношениях “дерево–насекомое” для конкретных видов насекомых и их кормовых пород (Исаев и др., 1997, 1999).
Как отмечено выше, на дубе обычно обитает многовидовой комплекс филлофагов, находящихся в сложных взаимоотношениях. В периоды вспышек размножения массовых вредителей численность прочих видов подавляется. При резком увеличении численности какого-либо вида возникает межвидовая, а затем и внутривидовая конкуренция. Важную роль для победы в межвидовой конкуренции играет скорость развития насекомых в период питания, которая тесно связана с погодными условиями, прежде всего с температурным режимом воздуха.
Как показали наши наблюдения, развитие гусениц зеленой дубовой листовертки и зимней пяденицы происходит быстро, в условиях ТОЛ в среднем в течение трех недель. Это дает им преимущества перед многими другими конкурирующими видами насекомых (Рубцов, Рубцова, 1984; Рубцов, Уткина, 2008).
Колебания численности особей в популяции листовертки происходят под действием большого числа разнообразных модифицирующих и регулирующих факторов, последние из которых обычно не образуют эффективный и гибкий механизм отрицательной обратной связи, что предопределяет длительность вспышек размножения этого вида, часто принимающих перманентный характер. Основными факторами динамики численности популяции листовертки являются:
модифицирующие факторы – метеорологические факторы, прямое и косвенное влияние которых на популяцию листовертки наиболее существенно проявляется в виде: 1) сильных зимних морозов, когда температура воздуха ниже значений, равных среднему из абсолютных минимумов, что вызывает гибель яиц листовертки и сказывается на выживаемости ее энтомофагов); 2) теплой засушливой погоды в период фазы гусеницы, что способствует улучшению кормовых свойств листвы, быстрому развитию гусениц, увеличению их выживаемости и плодовитости бабочек; 3) характера (численных значений и особенностей распределения) температур воздуха зимой и в ранневесенний период, который определяет ход эмбрионального развития листовертки и почек дуба и иногда сильно влияет на выживаемость гусениц; 4) весенних заморозков, приводящих к гибели почек и листвы и увеличивающих смертность гусениц листовертки;
регулирующие факторы, связанные с внутривидовой конкуренцией, которые обладают высоким порогом активности и оказывают существенное регулирующее воздействие при перенаселенности. Для зеленой дубовой листовертки это снижение плодовитости и бесплодие бабочек-самок, а также миграции, увеличивающие смертность особей и выравнивающие плотность популяции. Меньшим порогом активности обладают биоценотические регуляторные механизмы. Важная роль здесь принадлежит энтомофагам, видовой состав которых в Теллермановской дубраве очень богат. Определенная регулирующая роль принадлежит птицам и периодически болезням.
Процесс регулирования плотности популяции листовертки часто удается удовлетворительно имитировать и прогнозировать с помощью разработанной математической модели. Прогнозировать же модификацию плотности очень сложно вследствие ее вероятностного характера и существенной нестационарности.
Имитационные эксперименты с построенной математической моделью для изучения прямого действия погодных факторов, прежде всего температурного режима воздуха, на взаимоотношения гусениц зеленой дубовой листовертки с дубом показали, что при очень высоких плотностях популяции листовертки средний вес ее гусениц в конце питания листвой перед окукливанием оказывается равным или ниже при благоприятных для развития гусениц погодных условиях, чем при средних или плохих (Рубцов, Рубцова, 1984). Мы объясняем это внутривидовой конкуренцией гусениц за пищу. При благоприятных погодных условиях интенсивность питания и миграций гусениц возрастает, они быстро уничтожают всю листву на деревьях, при этом средняя масса их мала. Часть гусениц не в состоянии окуклиться из-за недокорма и погибает, образовавшиеся куколки очень мелкие, а вылетевшие из них бабочки - неплодовитые. В то же время, при худших для развития гусениц листовертки погодных условиях интенсивность их питания ниже, а смертность обычно выше. Нарастание массы листвы опережает степень ее уничтожения, конкуренция за пищу уменьшается, выжившая часть гусениц имеет более высокий вес. В условиях перенаселенности это может способствовать сохранению популяции.
Степень дефолиации крон деревьев к моменту окукливания филлофагов определяется массой и скоростью роста листвы в период питания гусениц, интенсивностью питания и динамикой численности гусениц. С помощью моделирования показано, что при малой численности филлофагов погодные условия в период питания гусениц существенно влияют на степень дефолиации крон, а при высокой – значительно меньше. То есть, при достижении популяцией вредителя достаточно высокой численности регулирующая роль погодного фактора в фазе личинки уменьшается. При дальнейшем увеличении численности филлофагов, в условиях перенаселенности, значение регулирующей роли погодного фактора опять возрастает, о чем говорилось выше. Эти примеры наглядно говорят об относительности “благоприятной погодной ситуации” для развития конкретной фазы определенного вида насекомых.
Из приведенных результатов моделирования мы видим довольно сложное и неоднозначное воздействие погодных факторов на взаимоотношения популяции филлофага с кормовыми растениями. Наши выводы совпадают с результатами других исследований (Исаев и др., 1999; Лямцев, Исаев, 2005; Ramsfield et al., 2016; Kolb et al., 2016; и др.).
Важнейшим фактором, влияющим на дальнейшую судьбу деревьев после интенсивной дефолиации, является количество и сроки образования регенеративной листвы – уровень рефолиации. Потенциальная способность растений компенсировать листовую поверхность взамен утраченной характеризует толерантность растений к филлофагам. Выводы исследователей о влиянии рефолиации (восстановительного побего- и листо- образования в кронах) на состояние дерева различны, что в значительной мере объясняется сложным характером связи регенеративного побегообразования с дефолиацией и другими биотическими и абиотическими факторами.
При исследовании взаимоотношений между филлофагами и их кормовыми породами необходимо учитывать, помимо абиотических факторов и особенностей насекомых, также и экофизиологические особенности конкретной древесной породы (типы формирования побегов, скорость роста побегов и листьев, особенности формирования кроны, корневой системы и др.), а также индивидуальные характеристики конкретных деревьев: условия произрастания, возраст, положение в древостое и др. К важнейшим показателям видовой стратегии относится то, как и какие побеги образует дерево. Например, у дуба черешчатого и других видов дуба рост побегов детерминированный (в течение одного или нескольких коротких периодов), у березы – недетерминированный (происходит в течение всего периода вегетации). Имеют большое значение и сроки повреждения листвы – в начале вегетационного периода, в середине или конце, и, безусловно, степень повреждения, т.е. дефолиации. Процессы регенеративного побего- и листообразования в кронах дуба черешчатого после полной или частичной дефолиации филлофагами подробно рассмотрены нами ранее (Рубцов, Уткина, 2008).
Любопытные факты, подтверждающие полученные нами данные для дуба черешчатого, содержатся в работе, посвященной защитным реакциям разных видов дуба (Pearse, Hipp, 2012). Авторы с помощью метода филогенетической регрессии проанализировали различные характеристики 56 видов дуба, произрастающих в Европе, Азии и Северной Америке. По их мнению, климат оказывает решающее влияние как на химические, так и на физические защитные реакции. Защитные реакции листьев в ответ на нападение листогрызущих насекомых и минеров у одного и того же вида дуба были выше в местообитаниях на меньшей географической широте, что авторы интерпретируют как косвенное действие климата. При сравнении разных видов было установлено, что защитные свойства были выше у тех видов дуба, которые произрастают в регионах с низкой температурой, умеренной зимой и низким минимальным количеством осадков. Установленная связь между признаками листьев дуба и абиотической средой согласуется с набором климатических параметров, которые оказывают влияние на фитофагов, и не противоречит гипотезе доступности ресурсов, согласно которой чем более ограничены кормовые растения в ресурсах, тем сильнее воздействуют на них фитофаги, осуществляя естественный отбор.
Заключение. Выполнен анализ временных рядов и выявлены тренды основных метеоэлементов в южной лесостепи. Показано уменьшение континентальности климата в этом регионе, значительное увеличение количества осадков за вегетацию, увеличение частоты периодов с аномальными погодными ситуациями. Особенности современного климата оказывают большое влияние на растения и насекомых и их биоценотические взаимосвязи. Это однозначно подтверждают выполненные исследования во многих странах мира, краткий обзор которых приведен в статье. Большинство авторов отмечают, что температура воздуха является самым важным фактором, непосредственно влияющим на развитие и численность насекомых в популяциях. Зафиксированы изменения ареалов многих видов растений и насекомых в виде сдвигов в северном направлении и вверх по высоте над уровнем моря, более раннее наступление весенних фенофаз и более позднее – осенних, что приводит к удлинению вегетационного периода.
Наблюдения в Теллермановской дубраве показали, что особенности современного климата приводят к заметным изменениям сложившихся в данных условиях циклов динамики численности популяций насекомых, нарушают баланс их взаимоотношений с кормовыми растениями. Это относится, в частности, к популяциям некоторых хозяйственно значимых филлофагов – непарного шелкопряда, зеленой дубовой листовертки, зимней пяденицы. Происходит уменьшение вредоносности одних видов и увеличение – других.
В настоящее время с помощью моделирования можно прогнозировать некоторые последствия прямого воздействия изменения климата на взаимоотношения насекомых-филлофагов с кормовыми растениями. Отдаленные последствия, связанные с косвенным воздействием климата, предсказать сейчас почти невозможно. Они будут зависеть, с одной стороны, от характера изменений климата, с другой стороны – от реакции на это растений и насекомых, обусловленной их адаптационными механизмами.
Список литературы
Бирюков В.И., Марченко Н.Ф. Характеристика погодно-климатических показателей территории Хоперского заповедника и их изменения за период 1939–2000 гг. // Влияние изменения климата на экосистемы. Охраняемые природные территории России: Анализ многолетних наблюдений. М.: Русский университет, 2001. С. II-82–86.
Венгеров П.Д., Сапельникова И.И., Базильская И.В., Масалыкин А.И. Климатические изменения и вызываемые ими прямые и косвенные эффекты в Воронежском заповеднике // Влияние изменения климата на экосистемы. Охраняемые природные территории России: Анализ многолетних наблюдений. М.: Русский университет, 2001. С. II-39–47.
Воронцов А.И. Патология леса. М.: Лесн. пром-сть, 1978. 270 с.
Голуб В.Б., Бережнова О.Н., Корнев И.И. Массовое размножение дубовой широкоминирующей моли, Acrocercops brongniardella F. (Lepidoptera, Gracillariidae), в Воронежской области // Известия Санкт-Петербургской лесотехнической академии. 2009. Вып. 187. С. 96–102.
Голуб В.Б., Простаков Н.И., Хицова Л.Н. Динамика поврежденности кроны дуба широкоминирующей молью (Acrocercops brongniardella F., Lepidoptera, Gracillariidae) в Усманском бору (Воронежская область) // Известия Санкт-Петербургской лесотехнической академии. 2011. Вып. 196. С. 29–36.
Егоров Н.Н., Рубцова Н.Н., Соложеникина Т.Н. Учет дубовой листовертки по яйцекладкам // Лесное хозяйство. 1953. № 10. С. 47–49.
Ефремова В.А. Учет движения численности зеленой дубовой листовертки: Автореф. дис…. канд. биол. наук. М.: МЛТИ, 1973. 21 с.
Исаев А.С., Овчинникова Т.М., Пальникова Е.Н., Суховольский В.Г. Имитационное моделирование динамики численности сосновой пяденицы при различных климатических сценариях // Лесоведение. 1997. № 4. С. 40–48.
Исаев А.С., Овчинникова Т.М., Пальникова Е.Н., Суховольский В.Г., Тарасова О.В. Оценка характера взаимодействий “лес – насекомые” в лесах бореальной зоны в ходе возможных климатических изменений // Лесоведение. 1999. № 6. С. 39–44.
Лямцев Н.И., Исаев А.С. Модификация типов вспышек массового размножения непарного шелкопряда в зависимости от эколого-климатической ситуации // Лесоведение. 2005. № 5. С. 3–9.
Лямцев Н.И., Исаев А.С., Зукерт Н.В. Влияние климата и погоды на динамику численности непарного шелкопряда в европейской России // Лесоведение. 2000. № 1. С. 62–67.
Мусолин Д.Л., Саулич А.Х. Реакции насекомых на современное изменение климата: от физиологии и поведения до смещения ареалов // Энтомологическое обозрение. 2012. Т. 41. № 1. С. 3–35.
Рубцов В.В., Рубцова Н.Н. Анализ взаимодействия листогрызущих насекомых с дубом. М.: Наука, 1984. 184 с.
Рубцов В.В., Уткина И.А. Адаптационные реакции дуба на дефолиацию. М.: Гриф и К, 2008. 302 с.
Рубцов В.В., Уткина И.А. Филлофаги лесных экосистем в условиях изменяющегося климата // Вестник Марийского государственного технического университета. Серия: Лес. Экология. Природопользование. 2010. № 3. С. 3–15.
Рубцов В.В., Уткина И.А. Многолетняя динамика численности зимней пяденицы в дубравах лесостепи // Лесоведение. 2011. № 5. С. 36–45.
Уткина И.А., Рубцов В.В. Современные представления о влиянии изменений климата на взаимодействие лесных деревьев и насекомых-фитофагов // Лесной вестник / Forestry Bulletin. 2017. Т. 21. № 6. С. 5–12.
Уткина И.А., Рубцов В.В. Общие закономерности и видовые различия в реакции представителей рода Quercus на дефолиацию // Мониторинг и биологические методы контроля вредителей и патогенов древесных растений: от теории к практике. Матер. Второй Всерос. конф. с междунар. участием. Москва, 22–26 апреля 2019 г. М.; Красноярск: ИЛ СО РАН, 2019. С. 178–179.
Bale J.S., Masters G.J., Hodkinson I.D., Awmack C., Bezemer T.M., Brown V.K, Butterfield J., Buse A., Coulson J.C., Farrar J., Good J.G., Harrington R., Hartley S., Jones T.H., Lindroth R.L., Press M.C., Symrnioudis I., Watt A.D., Whittaker J.B. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores // Global Change Biology. 2002. V. 8. № 1. P. 1–16.
Buse A., Good J.E.G. 1996. Synchronization of larval emergence in winter moth (Operophtera brumata L.) and budburst in pedunculate oak (Quercus robur L.) under simulated climate change // Ecological Entomology. 1996. V. 21. № 4. P. 335–343.
Harrington R., Woiwod I., Sparks T. Climate change and trophic interactions // Trends in Ecology & Evolution. 1999. V. 14. № 4. P. 146–150.
Heimonen K., Valtonen A., Kontunen-Soppela S., Keski-Saari S., Rousi M., Oksanen E., Roininen H. Colonization of a host tree by herbivorous insects under a changing climate // Oikos. 2015. V. 124. № 8. P. 1013–1022.
Hittenbeck A., Bialozyt R., Schmidt M. Modelling the population fluctuation of winter moth and mottled umber moth in Central and Northern Germany // Forest Ecosystems. 2019. V. 6. № 4. 18 p.
Hudges L. Biological consequences of global warming: is the signal already apparent? // Trends in Ecology & Evolution. 2000. V. 15. № 2. P. 56–61.
Jaworski T., Hilszczański J. The effect of temperature and humidity changes on insects development and their impact on forest ecosystems in the expected climate change // Forest Research Papers. 2013. V. 74. № 4. P. 345–355.
Kolb T.E., Fettig C.J., Ayres M.P., Bentz B.J., Hicke J.A., Mathiasen R., Stewart J.E., Weed A.S. Observed and anticipated impacts of drought on forest insects and diseases in the United States // Forest Ecology and Management. 2016. V. 380. P. 321–334.
McCarty J.P. Ecological consequences of recent climate change // Conservation Biology. 2001. V. 15. № 2. P. 320–331.
Parmesan C. Ecological and evolutionary responses to recent climate change // Annual Review of Ecology, Evolution, and Systematics. 2006. V. 37. № 1. P. 637–669.
Pearse I.S., Hipp A.L. Global patterns of leaf defenses in oak species // Evolution. 2012. V. 66. № 7. P. 2272–2286.
Pureswaran D.S., Roques A., Battisti A. Forest insects and climate change // Current Forestry Reports. 2018. V. 4. № 2. P. 35–50.
Ramsfield T.D., Bentz B.J., Faccoli M., Jactel H., Brockerhoff E.G. Forest health in a changing world: effects of globalization and climate change on forest insect and pathogen impacts // Forestry. 2016. V. 89. № 3. P. 245–252.
Root T.L., Price J.T., Hall K.R., Schneider S.H., Rosenzweig C., Pounds J.A. Fingerprints of global warming on wild animals and plants // Nature. 2003. V. 421. № 6918. P. 57–60.
Schwartzberg E.G., Jamieson M.A., Raffa K.F., Reich P.B., Montgomery R.A., Lindroth R.L. Simulated climate warming alters phenological synchrony between an outbreak insect herbivore and host trees // Oecologia. 2014. V. 175. № 3. P. 1041–1049.
Дополнительные материалы отсутствуют.