

Лесоведение, 2020, № 4, стр. 377-384
Жизнеспособность пыльцы ели сибирской в южной тайге Красноярского края
Е. В. Бажина a, *, М. И. Седаева a, А. К. Экарт a
a Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: genetics@ksc.krasn.ru
Поступила в редакцию 21.12.2018
После доработки 26.03.2019
Принята к публикации 29.01.2020
Аннотация
Исследована жизнеспособность пыльцы ели сибирской (Picea obovata Ledeb.) в оптимальных условиях произрастания. Выявлена изменчивость и скоррелированность морфологических характеристик пыльцевых зерен, низкое число аномалий их развития. Проращивание in vitro показало значительное варьирование показателей жизнеспособности. Изменчивость функциональных показателей пыльцы у ели сибирской свидетельствует о высокой пластичности и адаптивности репродуктивных процессов данного вида.
Ель сибирская (Picea obovata Ledeb.), один из основных лесообразователей Сибири, занимает обширный ареал от северо-востока Европейской части России до побережья Охотского моря и горных районов Урала, Алтая и Саян (Атлас, 1973). В пределах ареала еловые леса приурочены в основном к влажным экотопам Европейского севера, гор Урала и Сибири. Распространение ели сибирской свидетельствует о приспособленности к воздействиям экстремальных факторов внешней среды, что в значительной степени определяется эффективностью размножения вида (Грант, 1991).
Одним из важнейших элементов репродуктивной системы растений, оказывающих непосредственное влияние на эффективность семенного размножения, являются мужские и женские гаметы (Singh, 1978; Delph et al., 1997; Runions, Owens, 1999). У хвойных мужских гамет (пыльцевых зерен) формируется на несколько порядков больше, чем женских, они более подвержены воздействию внешних факторов и, очевидно, играют ведущую роль в гаметном отборе (Friedman, Floyd, 2001). Дефицит жизнеспособной пыльцы у растений может быть одним из основных факторов, ответственных за нарушения оплодотворения и низкую продукцию жизнеспособных семян (Arista, Talavera, 1994; Bazhina, 2018).
Развитие и функции пыльцевых зерен различных видов рода Picea исследовались на протяжении ХХ в. (Hutchinson, 1915; Andersson, 1980; Owens et al., 1987). Показано, что жизнеспособность пыльцы зависит от особенностей генотипа, морфологических и физиологических характеристик пыльцевых зерен, химических и физических условий прорастания (Christiansen, 1972; Dawkins, Owens, 1993; Nikkanen et al., 2000). В экстремальных условиях произрастания – на границе ареала, при интродукции, загрязнении среды – фертильность и жизнеспособность пыльцы, как правило, снижаются (Владимирова и др., 2008; Калашник, 2008). Однако виды рода Picea демонстрируют удовлетворительную жизнеспособность пыльцы при интродукции (Макогон, Коршиков, 2012; Бажина, Седаева, 2017).
Перспективность существования вида в значительной степени определяется биотическим (репродуктивным) потенциалом, который проявляется в оптимальном для его биологических возможностей сочетании условий среды (Chapman, 1928; Грант, 1991). Исследования, проводимые в оптимальных условиях, позволяют охарактеризовать степень эволюционной адаптации вида, его репродуктивный потенциал, дать прогноз получения продуктивного потомства, а также вносят существенный вклад в развитие лесного семеноводства.
Цель настоящих исследований заключалась в изучении жизнеспособности пыльцы ели сибирской (Picea obovata Ledeb.) в природных популяциях юга Средней Сибири.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводились в лесных биоценозах долин рр. Еловка и Сухая. Для территории исследований характерен холмисто-увалистый рельеф, с чередованием плоских водораздельных участков и широких логов как с пологими, так и резко крутыми склонами. Микрорельеф развит в виде бугров и понижений. Почвенный покров представлен темно-серыми и серыми лесными почвами, преобладает разнотравная группа типов леса. Климат района резко континентальный, с господствующим юго-западным направлением ветров, холодной зимой и жарким летом. Среднегодовая температура воздуха составляет 0.5°С, среднегодовое количество осадков – 485 мм/год. Континентальность климата выражена значительной годовой (38°С по среднемесячным значениям) и суточной (12–14°С) амплитудой колебаний температуры воздуха (Агроклиматический справочник, 1961). Средняя продолжительность безморозного периода – 120 дней, средняя дата последнего заморозка – 22 мая. В зимний и летний периоды над районом устанавливается отрог Сибирского антициклона, который приносит холодные воздушные массы и формирует зимой холодную ясную погоду с сильными морозами, а летом – ясную, жаркую. Весной и осенью характер погоды неустойчив. Влияние вредных выбросов промышленных центров незначительно ввиду удачного географического положения и господствующего направления ветров.
Сбор пыльцы проводился в 2017 и 2018 г. в период массового пыления (первая декада июня) с деревьев ели сибирской, произрастающих в ельниках разнотравных в долине р. Еловка (56°08′ с.ш. и 92°32′ в.д.) – пробная площадь “Еловка” и долине р. Сухая (56°22′ с.ш. и 92°45′ в.д.) – пробная площадь “Сухая”. Погодные условия в период исследований оказались благоприятными для развития пыльцы и пыления (положительные среднесуточные температуры воздуха). Количество осадков за период пыления составляло (мм) 0.6 (2018 г.) – 13.6 (2017 г.), дожди наблюдались, в основном, в ночные и вечерние часы. Сумма эффективных температур (Tэф > 5°) рассчитывалась согласно общепринятой методике (Агроклиматический справочник, 1961; Sarvas, 1967). Пыльца собиралась с деревьев, растущих в центре насаждений, а в долине р. Еловка дополнительно и на опушке на обращенном к открытому пространству склоне. На каждой пробной площади с верхней части кроны 20–25 типичных деревьев секатором срезалось несколько веток, пыльники отделялись от ветвей, пыльца просеивалась через марлю и упаковывалась в кальку.
В лабораторных условиях у свежесобранной пыльцы определялись размеры пыльцевых зерен: длина и высота тела, длина и высота воздушных мешков, а также частота встречаемости и спектр аномалий, потенциальная и реализованная жизнеспособность. Для исследования морфологии пыльцу окрашивали ацетокармином. Морфометрические параметры пыльцы определялись для каждого дерева у 25–30 нормально развитых пыльцевых зерен в трехкратной повторности, затем данные усреднялись для каждого дерева и далее – для каждой пробной площади. Потенциальная жизнеспособность определялась по гистохимической реакции на содержание крахмала (Дженсен, 1965). Для определения фактической жизнеспособности пыльцу каждого дерева проращивали in vitro на 10%-ном растворе сахарозы при температуре 26°С (Паушева, 1980) в течение 5 суток. Оценка проводилась по числу проросших пыльцевых зерен (%) – по 100–120 шт. для каждого дерева, и длине пыльцевых трубок (мкм) – по 30–40 шт. на дерево; данные усреднялись для каждого дерева и пробной площади. Статистическая обработка проводилась по общепринятым методикам с использованием пакета анализа MICROSOFT EXCEL 2000. Достоверность различий оценивалась дисперсионным анализом, уровень изменчивости определялся согласно С.А. Мамаеву (1972).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Весеннее развитие микростробилов у ели сибирской начинается в третьей декаде апреля при сумме эффективных температур Тэф > 5° = 48.8–55.6 град.-дней (3.0–3.4% от Тэф>5° за год). Пыление наблюдается во второй декаде мая при сумме эффективных температур 150.0–151.0 град.-дней (9.2–9.3% от Тэф > 5° за год), что соответствует значениям, установленным для ели сибирской (Некрасова, 1976). Зрелая пыльца, как и у других видов рода Picea (Бажина, Седаева, 2017), состоит из остатков двух проталлиальных, сифоногенной, спермиогенной и стерильной клеток.
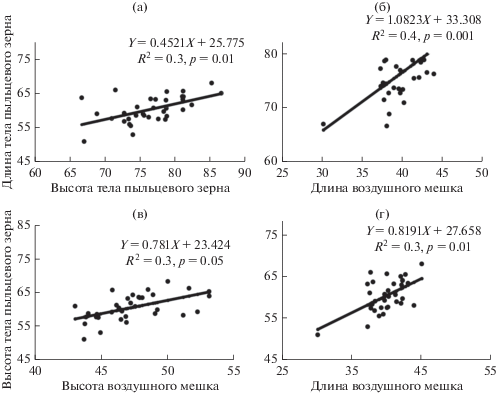
Размеры пыльцевых зерен характеризуются достаточно низким уровнем изменчивости (табл. 1). Средние различия морфометрических показателей размеров пыльцевых зерен у деревьев, растущих в пределах одного насаждения (7.7–18.0%) статистически не достоверны, между насаждениями составляли 8.8–15.3%. Морфометрические показатели тела и воздушных мешков пыльцевых зерен достоверно коррелировали при р = 0.05–0.001 (рис. 1), однако связи слабо функциональны (коэффициент детерминации не превышал 0.3–0.4).
Таблица 1.
Морфометрические показатели пыльцы ели сибирской
| Пробная площадь | Размеры пыльцевого зерна, мкм | |||
|---|---|---|---|---|
| тело | воздушный мешок | |||
| длина* | высота | длина* | высота | |
| 2017 г. | ||||
| Еловка | $\frac{{78.4 \pm 0.27}}{{57.9 - 97.0}}$ | $\frac{{60.3 \pm 0.26}}{{38.1 - 76.8}}$ | $\frac{{40.7 \pm 0.20}}{{25.5 - 53.9}}$ | $\frac{{47.2 \pm 0.21}}{{34.0 - 62.5}}$ |
| Сухая | $\frac{{74.6 \pm 0.33}}{{53.3 - 103.7}}$ | $\frac{{60.6 \pm 0.31}}{{34.1 - 85.6}}$ | $\frac{{39.0 \pm 0.27}}{{21.1 - 65.6}}$ | $\frac{{47.2 \pm 0.28}}{{31.3 - 68.6}}$ |
| 2018 г. | ||||
| Еловка | $\frac{{71.0 \pm 0.28}}{{49.3 - 95.5}}$ | $\frac{{49.9 \pm 0.28}}{{29.3 - 73.0}}$ | $\frac{{43.4 \pm 0.21}}{{24.1 - 61.1}}$ | $\frac{{41.6 \pm 0.22}}{{26.3 - 64.4}}$ |
| Сухая | $\frac{{69.5 \pm 0.33}}{{50.8 - 92.1}}$ | $\frac{{49.1 \pm 0.36}}{{31.2 - 72.8}}$ | $\frac{{41.6 \pm 0.25}}{{21.6 - 59.3}}$ | $\frac{{42.0 \pm 0.35}}{{25.2 - 72.5}}$ |
Примечание. В табл. 1 и 2 в числителе приведены среднее значение ± ошибка среднего, в знаменателе – минимальные и максимальные значения признака. * Различия на разных пробных площадях достоверны при p = 0.05.
Рис. 1.
Взаимосвязь между морфометрическими показателями, мкм: длиной тела пыльцевых зерен и высотой тела (а), длиной воздушных мешков (б), между высотой тела пыльцевых зерен и высотой воздушных мешков (в), длиной воздушных мешков (г). R2 – коэффициент детерминации.
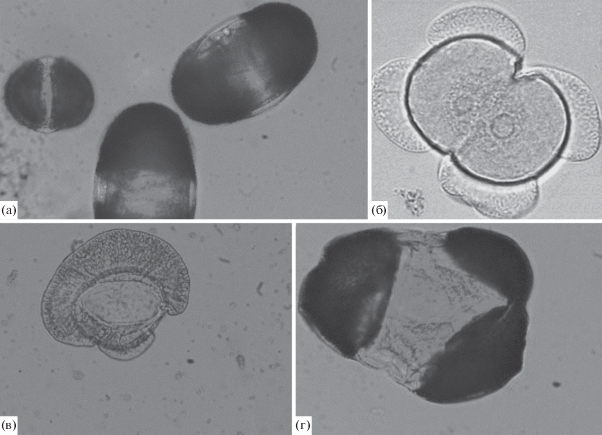
Обследованные популяции характеризуются достаточно низким уровнем нарушений пыльцы. Аномальные пыльцевые зерна выявлены у 41.2% обследованных деревьев, доля у отдельных из них не превышала 8.16% (табл. 2). Наиболее часто встречающейся аномалией являются мелкие, стерильные пыльцевые зерна (рис. 2а). Предполагается, что они формируются вследствие нарушений мейоза, когда часть генетического материала утрачивается в результате аномалий веретена деления или хромосомных мутаций (Rana et al., 2013). Другие типы нарушений (менее 0.28%) представлены аномалиями по числу воздушных мешков – с 1 воротничковым или 3–4 воздушными мешками, а также сросшимися по проксимальной стороне пыльцевыми зернами (рис. 2б–2г).
Таблица 2.
Жизнеспособность пыльцы ели сибирской
| Пробная площадь | Доля, % | Длина пыльцевых трубок, мкм | ||
|---|---|---|---|---|
| окрашенных пыльцевых зерен | проросших пыльцевых зерен | аномальных пыльцевых зерен | ||
| 2017 г. | ||||
| Еловка | $\frac{{98.2 \pm 0.31}}{{94.9 - 99.7}}$ | $\frac{{62.3 \pm 3.47}}{{35.8 - 89.3}}$ | $\frac{{1.51}}{{0 - 7.94}}$ | $\frac{{118.81 \pm 2.313}}{{15.88 - 414.31}}$ |
| Сухая | $\frac{{98.1 \pm 0.39}}{{94.2 - 99.6}}$ | $\frac{{60.9 \pm 5.40}}{{15.2 - 92}}$ | $\frac{{0.79}}{{0 - 2.73}}$ | $\frac{{149.32 \pm 4.116}}{{18.76 - 524.27}}$ |
| 2018 г. | ||||
| Еловка | $\frac{{94.1 \pm 0.95}}{{83.0 - 100}}$ | $\frac{{63.5 \pm 4.88}}{{10.3 - 100}}$ | $\frac{{2.21}}{{0 - 7.41}}$ | $\frac{{194.58 \pm 2.255}}{{16.36 - 1219.47}}$ |
| Сухая | $\frac{{95.4 \pm 0.66}}{{90.1 - 98.0}}$ | $\frac{{89.7 \pm 3.17}}{{56.3 - 94.3}}$ | $\frac{{2.43}}{{0 - 8.16}}$ | $\frac{{212.46 \pm 4.888}}{{15.27 - 1166.67}}$ |
Примечание. Пояснение – см. табл. 1.
Рис. 2.
Аномалии пыльцы: мелкое пыльцевое зерно (а), сросшиеся пыльцевые зерна (б), пыльцевое зерно с воротничковым воздушным мешком (в), пыльцевое зерно с тремя воздушными мешками (г).
Жизнеспособность пыльцы в значительной степени зависит от содержащихся в ней питательных и физиологически активных веществ, так как в начальный период прорастания пыльцевые трубки развиваются, главным образом, за счет собственных запасов (Поддубная–Арнольди, 1964). Пыльца видов рода Picea, как и у большинства представителей голосеменных, относится к так называемому “крахмальному типу”. Гистохимический анализ пыльцевых зерен показал, что в отдельные годы исследований у 76–97% деревьев более 90% пыльцевых зерен дают положительную реакцию на основное питательное вещество – крахмал (табл. 2).
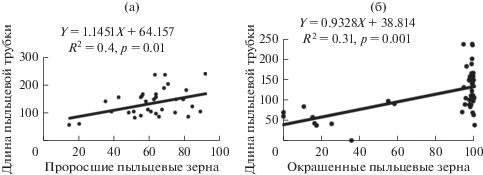
Высокая потенциальная жизнеспособность реализуется при прорастании пыльцы – практически все деревья продуцировали жизнеспособную пыльцу. При проращивании на искусственных средах пыльца считается жизнеспособной, если длина пыльцевых трубок превышает размеры тела пыльцевого зерна (Hak, Russel, 2004). Показатели жизнеспособности достоверно коррелируют между собой (рис. 3а), при этом наблюдается значительное их варьирование у отдельных деревьев в разные годы и в зависимости от условий произрастания: по числу проросших пыльцевых зерен – в 1.6–9.7 раз, по длине пыльцевых трубок – в 26–76 раз. Исследования выявили положительные зависимости между длиной пыльцевых трубок и содержанием крахмала в пыльце (рис. 3б), однако дисперсионный анализ показал, что связи не всегда функциональны.
Рис. 3.
Регрессия между долей прорастания пыльцы, % и длиной пыльцевых трубок, мкм, (а), длиной пыльцевых трубок и числом пыльцевых зерен, давших положительную реакцию на крахмал (б).
Морфологические и функциональные характеристики пыльцы в различных географических и экологических условиях характеризуют адаптационные процессы хвойных видов (Козубов, 1974; Некрасова, 1976). Родиной ели сибирской считаются горные районы Южной Сибири, она хорошо адаптирована к резко континентальному климату (Поликарпов, 1970; Попов, 1999). Развитие мужских генеративных структур и пыление у ели сибирской начинается при более низкой сумме эффективных температур, чем у других видов рода Picea (Sarvas, 1967; Major et al., 2005), что, вероятно, является биологической особенностью данного вида и свидетельствует о высокой подготовленности репродуктивных структур к весеннему развитию.
Условия формирования микроспор оказывают влияние на их репродуктивный потенциал, благоприятные способствуют повышению адаптационных возможностей (Sarvas, 1967; Prasad et al., 2011). При отклонении условий среды от оптимальных у растений происходит снижение морфогенетического гомеостаза и стабильности развития. Виды с высоким адаптивным потенциалом демонстрируют низкий уровень изменчивости морфологических признаков и аномалий развития (Мамаев, 1972; Holsinger, 2000). Исследования показали низкий уровень аномалий развития, скоррелированность морфологических характеристик пыльцевых зерен и более высокую их изменчивость в исследованных популяциях по сравнению с северными и южными (Козубов, 1974; Владимирова, 2008). Фенотипическая пластичность свидетельствует об оптимальных для вида условиях в популяциях юга Средней Сибири.
Основная функция пыльцевых зерен – доставка мужских гамет к женской яйцеклетке – у ели сибирской, как и у большинства семенных растений, осуществляется посредством сифоногамии (роста пыльцевых трубок), на развитие которых негативное влияние оказывают как эндогенные (генотип, возраст, состояние растения, обеспеченность питательными веществами), так и экзогенные (химические и физические показатели среды) факторы (Stanley, 1971; Christiansen, 1972; Dawkins, Owens, 1993; Taylor, Hepler, 1997; Nikkanen et al., 2000; Galen, Stanton, 2003). Изменчивость функциональных показателей пыльцы у ели сибирской свидетельствует о высокой пластичности и адаптивности репродуктивных процессов данного вида.
Функциональные свойства пыльцы, как правило, тесно коррелируют с ее составом. Пыльца хвойных растений характеризуется высоким разнообразием химических соединений, таких как крахмал, углеводы, жиры, белки, аминокислоты, фитогормоны (Цингер, Размологов, 1972). Пыльца, содержащая большее количество углеводов, окислительных ферментов, питательных и физиологически активных веществ, характеризуется лучшей жизнеспособностью, так как на первых этапах прорастание происходит главным образом за счет собственных веществ, а затем используются запасные вещества среды, на которой она растет (Поддубная-Арнольди, 1964; Stanley, 1971). Настоящие исследования в целом подтверждают направление зависимостей между жизнеспособностью и обеспеченностью питательными веществами пыльцы, однако связи не всегда функциональны.
Заключение. Пыление ели сибирской в экосистемах Средней Сибири наблюдается во второй декаде мая, при сумме эффективных температур, соответствующей значениям, установленным для данного вида. Исследования показали скоррелированность морфологических характеристик пыльцевых зерен и их более высокую, чем в экстремальных условиях среды, изменчивость, функциональные зависимости между жизнеспособностью пыльцы и обеспеченностью пыльцевых зерен питательными веществами. Фенотипическая пластичность, достаточно низкий процент аномальных пыльцевых зерен, функциональные свойства пыльцы ели сибирской свидетельствуют об оптимальных для вида условиях, которые в настоящее время сложились в популяциях юга Средней Сибири. Наблюдающиеся вариации могут быть обусловлены генетическими особенностями и различиями реакций отдельных деревьев, определяющими их способность к адаптации, а также локальными различиями в условиях произрастания.
Список литературы
Агроклиматический справочник по Красноярскому краю и Тувинской автономной области. Л.: Гидрометеоиздат, 1961. 288 с.
Атлас лесов СССР. М.: ГУГК, 1973. 222 с.
Бажина Е.В., Седаева М.И. Жизнеспообность пыльцы некоторых видов Picea (Pinaceae) в условиях Красноярска // Ботанический журн. 2017. Т. 102. № 6. С. 768–779.
Владимирова О.С., Муратова Е.Н., Седаева М.И. Пыльца ели сибирской, произрастающей в различных экологических условиях // Хвойные бореальной зоны. 2008. Т. 25. № 1–2. С. 98–102.
Грант В. Эволюционный процесс: Критический обзор эволюционной теории. М.: Мир, 1991. 488 с.
Дженсен У.Д. Ботаническая гистохимия. М.: Мир, 1965. 377 с.
Калашник Н.А., Ясовиева С.М., Преснухина Л.П. Аномалии пыльцы хвойных видов деревьев при промышленном загрязнении на Южном Урале // Лесоведение. 2008. № 2. С. 33–40.
Козубов Г.М. Биология плодоношения хвойных на Севере. Л.: Наука, 1974. 136 с.
Макогон И.В., Коршиков И.И. Качество пыльцы в связи с генетическими особенностями Picea abies (L.) Karst. в интродукционном насаждении // Бюллетень государственного Никитского ботанического сада. 2012. Вып. 105. С. 107–112.
Мамаев С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). М.: Наука, 1972. 283 с.
Некрасова Т.П. Влияние температуры воздуха на формирование пыльцы хвойных древесных пород // Лесоведение. 1976. № 6. С. 37–43.
Паушева З.П. Практикум по цитологии растений. М.: Колос, 1980. 304 с.
Поддубная–Арнольди В.А. Общая эмбриология покрытосеменных растений. М.: Наука, 1964. 482 с.
Поликарпов Н.П. Комплексные исследования в горных лесах Западного Саяна // Вопросы лесоведения. 1970. Т. 1. С 26–79.
Попов П.П. Ель на востоке Европы и в Западной Сибири: популяционно-географическая изменчивость и ее лесоводственное значение. Новосибирск: Наука, 1999. 169 с.
Цингер Н.В., Размологов В.П. Эволюция мужского гаметофита голосеменных // Биохимия и филогения растений. М.: Наука, 1972. С. 163–198.
Andersson E. Temperature-Conditioned Irregularities in Pollen Mother Cells of Picea abies (L.) Karst. // Hereditas. 1980. V. 92. P. 27–35.
Arista M., Talavera S. Pollen Dispersal Capacity and Pollen Viability of Abies pinsapo Boiss. // Silvae Genetica. 1994. V. 43. P. 155–158.
Bazhina E. Siberian fir seed productivity in V.N. Sukachev Institute of Forest Arboretum, Russia // Proceedings of the EuroGard VII Congress European Botanic gardens in the decade on biodiversity challenges and responsibilities in the count-down towards 2020 / Eds. Joly E., Larpin D., Delmas V., Carmine B. Paris: BGCI, 2018. P. 312–321.
Chapman R.N. The quantitative analysis of environmental factors // Ecology. 1928. V. 9. P. 111–122.
Christiansen H. On the development of pollen and the fertilization mechanism of Picea abies (L.) Karst. // Silvae Genetica. 1972. V. 21. P. 51–61.
Dawkins M.D., Owens J.N. In vitro and in vivo pollen hydration, germination, and pollen-tube growth in white spruce, Picea glauca (Moench) Voss. // International J. Plant Sciences. 1993. V. 154. P. 506–521.
Delph L.F., Johannsson M.H., Stephenson A.G. How environmental factors affect pollen performance: ecological and evolutionary perspectives // Ecology. 1997. V. 78. P. 1632–1639.
Friedman W.E., Floyd S.K. The Origin of Flowering Plants and Their Reproductive Biology // Evolution. 2001. V. 55. P. 217–231.
Galen C., Stanton M.L. Sunny-side up: flower heliotropism as a source of parental environmental effects on pollen quality and performance in the snow buttercup, Ranunculus adoneus (Ranunculaceae) // American J. Botany. 2003. V. 90. P. 724–729.
Hak O., Russell J.H. Environmental Effects on Yellow-Cedar Pollen Quality // Forest Genetic Council of British Columbia. 2004. Extension Note № 5. P. 1–9.
Holsinger K.E. Reproductive systems and evolution in vascular plants // Proceeding NAS. 2000. V. 97(13). P. 7037–7042.
Hutchinson A. H. On the Male Gametophyte of Picea canadensis // Botanical Gazette. 1915. № 69. P. 457–472.
Major J.E., Mosseler A., Johnsen K.H., Rajora O.P., Barsi D.C., Kim K.-H., Park J.-M., Campbell M. Reproductive barriers and hybridity in two spruces, Picea rubens and Picea mariana, sympatric in eastern North America // Canadian J. Botany. 2005. V. 83. P. 163–175.
Nikkanen T., Aronen T., Häggman H., Venäläinen M. Variation in pollen viability among Picea abies genotypes – potential for unequal paternal success // Theoretical and A-pplied Genetics. 2000. V. 101. P. 511–518.
Owens J.N., Simpson S.J., Caron G. The pollination mechanism of Engelmann spruce (Picea engelmannii Parry) // Canadian J. Botany. 1987. V. 65. P. 1439–1450.
Prasad P.V., Boote K.J., Allen L.J. Longevity and temperature response of pollen as affected by elevated growth temperature and carbon dioxide in peanut and grain sorghum // Environmental and Experimental Botany. 2011. V. 1. P. 51–57.
Rana P.K., Kumar P., Singhal V.K. Spindle irregularities, chromatin transfer, and chromatin stickiness during male meiosis in Anemone tetrasepala (Ranunculaceae) // Turkish J. Botany. 2013. № 37. P. 167–176.
Runions C. J., Owens J.N. Sexual reproduction in interior spruce (Pinaceae). I. Pollen germination to archegonial maturation // International J. Plant Sciences. 1999. V. 160. P. 631–640.
Sarvas R. The annual period of development of forest trees // Proceedings of the Finnish Academy of Science and Letters. 1967. P. 211-231.
Singh H. Embryology of gymnosperms. Berlin: Gerbruder Borntraeger, 1978. 302 p.
Stanley R.G. Pollen chemistry and tube growth // Pollen: development and physiology / Ed. J. Heslop-Harrison. London: Butterworths, 1971. P. 131–155.
Taylor L.P., Hepler P.K. Pollen germination and tube growth // Annual Review of Plant Physiology and Plant Molecular Biology, 1997. V. 48. P. 461–491.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение