

Лесоведение, 2020, № 5, стр. 387-398
Пространственно-временная структура и динамика позднесукцессионного черневого кедровника Западного Саяна
Д. М. Данилина a, *, Д. И. Назимова a, М. Е. Коновалова a
a Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: dismailova@mail.ru
Поступила в редакцию 29.01.2018
После доработки 11.10.2018
Принята к публикации 06.06.2020
Аннотация
Приведены результаты стационарных исследований (с 1965 по 2017 г.) закономерностей динамики пространственной структуры малонарушенных черневых кедровников на примере разновозрастного кедровника осочково-крупнотравно-папоротникового на позднесукцессионной стадии развития (Западный Саян, 53°01′ с.ш., 92°59′ в.д., размер постоянной пробы 1.56 га). Показано, что на протяжении полувекового периода поддерживается динамическое равновесие в составе эдификаторного яруса, тогда как субэдификаторный ярус (пихтовый) и его возрастная структура меняются быстрее и влияют на взаимоотношения синузий нижних ярусов и их роль в процессе возобновления сосны кедровой сибирской (Pinus sibirica (Du Tour) и пихты сибирской (Abies sibirica (Ledeb.). Каждая синузия характеризуется относительным постоянством числа видов, состава доминантов и соотношения эколого-ценотических групп. Крупнотравно-папоротниковая, вейниково-щитовниковая, борцово-осочковая, осочковая, кисличная синузии разной степени сложности остаются постоянными элементами структуры одного типа леса. Выявлена роль отдельных синузий травяного покрова в возобновлении кедра в кедровниках крупнотравно-папоротниковой группы типов леса. В господствующей крупнотравно-папоротниковой синузии возобновление кедра протекает медленно и приурочено к микроповышениям, валежу и ветровальным комплексам. Напротив, в осочковой синузии, где снижена конкуренция трав, возможны периодические вспышки возобновления кедра в годы высокого плодоношения, но вероятность последующего выживания подроста низка из-за конкуренции с материнским древостоем. Полученные результаты позволяют прогнозировать возрастную динамику малонарушенного черневого кедровника на ближайшие десятилетия при колебаниях погодно-климатических условий, типичных для последнего векового цикла, но при отсутствии катастрофических нарушений. Разнообразие микробиотопов и синузий внутри биогеоценоза поддерживает устойчивость горной экосистемы в целом и будет иметь важное значение для моделирования процесса возобновления и размещения деревьев в пространстве на последующих стадиях развития пихтово-кедрового древостоя в черневом поясе гор.
Сложность пространственной структуры позднесукцессионных лесных сообществ является одним из важнейших факторов поддержания устойчивости лесных экосистем в условиях погодно-климатических флюктуаций и других форм внешнего воздействия. Однако вопросы упорядоченности элементов структуры зрелых (позднесукцессионных) сообществ в ходе возрастной динамики остаются недостаточно изученными в различных климато-географических районах.
Выявлению структурной организации лесных биогеоценозов на разных стадиях посвящено большое число работ. При этом особого внимания заслуживают многолетние комплексные исследования на постоянных объектах мониторинга (Дыренков, 1966; Лащинский, 1981; Комарова, 1992; Восточноевропейские …, 2004; Москалюк, 2012; Санников и др., 2012; Рубцов, Рыбакова, 2016; Bakker, 1996; Korb, Ranker, 2001; Closset-Kopp et al., 2006; Ismailova, Nazimova, 2010; Bilek et al., 2011; Vacek et al., 2017; и др.). Детальные исследования структуры черневых кедровых лесов в горах Западного Саяна проводятся с 1960 г. как часть программы по изучению лесных биогеоценозов, обладающих естественной устойчивостью и имеющих высокую научную ценность (Поляков, 2007; Назимова и др., 2015). Они могут служить эталонными лесами, сохранившими сложную пространственную структуру, как вертикальную (ярусную), так и горизонтальную (мозаичную), с выраженной “синузиальностью разного порядка” (Сукачев, 1972). Наряду с синузиальностью нижних ярусов существуют также “возрастные парцеллы” – как стадии популяционной динамики древесных видов внутри биогеоценоза (Дылис, 1969). Они играют основную роль в поддержании устойчивости эдификаторных видо–лесообразователей и непрерывном развитии лесного биогеоценоза. Горный рельеф обостряет проблему неоднородности экотопического фона внутри конкретного участка биогеоценоза, однако не менее выражена и роль фитоценоза, наиболее активного блока экосистемы, преобразующего внутреннюю среду обитания всей биоты и создающего разнообразие микроусловий (микробиотопов) внутри биогеоценоза.
Целью исследования послужило изучение пространственной структуры позднесукцессионного черневого кедровника, развивавшегося многие века в условиях избыточно влажного климата, горного рельефа и полного отсутствия пожаров и рубок. Выявление движущих сил и процессов динамики структуры малонарушенных пихтово-кедровых лесов важно для понимания закономерностей функционирования и прогнозирования их развития в условиях текущих и будущих природно-антропогенных изменений. С позиций учения о лесном биогеоценозе в ходе комплексных исследований предстояло раскрыть роль отдельных элементов структуры лесного фитоценоза в поддержании многолетней динамической устойчивости коренного биогеоценоза.
Кедровник осочково-крупнотравно-папоротниковый, один из самых распространенных типов леса в черневом поясе Западного Саяна, является репрезентативным для демонстрации разнообразия и динамики фитоценотической структуры черневого кедровника и специфики лесообразовательного процесса. Особенность черневых кедровников составляет мощное развитие травяного яруса, выполняющего важную роль в биологическом круговороте, сопоставимую с ролью эдификаторного яруса. Это принципиальное отличие черневых кедровников от горно-таежных неоднократно подчеркивали лесоводы, таксаторы, физиологи и другие специалисты (Кедровые …, 1985; Nazimova et al., 2014).
ОБЪЕКТЫ И МЕТОДИКА
Многолетнее изучение структуры черневых кедровников проводится с 1960-хх годов на базе Ермаковского научного стационара Института леса им. В.Н. Сукачева СО РАН. Постоянная пробная площадь (пр. пл.) 3 заложена в 1960 г. под руководством И.В. Семечкина. Объект расположен в черневом высотно-поясном комплексе типов леса (ВПК) на абсолютной высоте 530 м (координаты 53°01′ с.ш., 92°59′ в.д). Типы леса крупнотравно-папоротниковой группы являются наиболее широко распространенными в пределах черневого пояса на высотах от 350 до 800–900 м над ур. моря (Типы …, 1980). Доля кедровников крупнотравно-папоротниковых среди всех черневых кедровников на территории Танзыбейского участкового лесничества составляет 59%, доля пихтарников с кедром крупнотравно-папоротниковых – 32%. Они занимают 16 582 га (28% от общей площади черневого ВПК).
Район исследований характеризуется суммой температур за период выше +10°С порядка 1650–1700°С и суммой осадков за год 700–1300 мм. Продолжительность периода активной вегетации составляет 110–115 дней (Типы лесов …, 1980). Пр. пл. 3 общей площадью 1.56 га заложена в бассейне р. Малый Кебеж, в средней части склона юго-западной экспозиции крутизной 22°–25°.
Почва территории горная лесная бурая высокощебнистая на делювии хлоритовых сланцев, сформирована в условиях промывного режима и хорошего дренажа, развитого внутрипочвенного стока. В верхних горизонтах отмечено высокое содержание гумуса (15–26%). Плиты сланцев начинаются уже с глубины 20–40 см и покрыты глинистыми натеками, их количество и размеры увеличиваются с глубиной. Общая мощность почвенного профиля – 130 см (Вишнякова и др., 1968).
Насаждение представляет собой позднесукцессионную стадию развития со сложной пространственной (как вертикальной, так и горизонтальной) структурой всего сообщества, абсолютно разновозрастным строением ценопопуляций эдификаторных видов, единичным валежом различной степени разложения и отсутствием следов масштабных нарушений (рубок и пожаров). Проведено три этапа перечетной таксации всего древостоя (1965, 1990, 2017 гг.). Возраст деревьев, каждое из которых пронумеровано, актуализировали по данным инструментального определения, выполненного в 1999 г. В соответствии классификацией, предложенной С.А. Дыренковым (1984) и И.В. Семечкиным (2002) возрастная структура древостоя сосны кедровой сибирской ((Pinus sibirica (Du Tour), далее – кедра) отнесена к циклично-разновозрастному типу, а пихты сибирской ((Abies sibirica (Ledeb.), далее – пихты) – к абсолютно разновозрастному. Условное разделение древостоя на поколения осуществлялось в соответствии с принятым в таксации правилом: возраст деревьев одного поколения не должен отличаться более чем на два класса (для кедра – 80 лет, для пихты – 40 лет). Проводились повторные геоботанические описания с привязкой к определенному сезонному состоянию по общей методике В.Н. Сукачева и С.В. Зонна (1961). Вертикальная и горизонтальная неоднородность фитоценотической структуры изучались методами картирования и профилирования. Горизонтальная структура древостоя и травяного яруса закартирована в масштабе 1 : 100, для чего вся площадь разбивалась на квадраты 25 м2. Мозаичность травяного покрова изучалась на уровне синузий. Под синузиями (термин предложенный Х. Гамсом в 1918 г.) понимаются “…структурные части фитоценозов и биогеоценозов, характеризующиеся определенным видовым составом, определенным экологическим характером видов их составляющих и пространственной обособленностью, а, следовательно, и особой фитоценотической средой (микросредой), создаваемой растениями данной синузии” (цит. по: (Сукачеву, 1972, с. 271)). Термин трактуется широко и предполагает разное время существования синузий в биогеоценозе (фитоценозе). Описания каждой синузии выполнялись в разные сезонные сроки (1972, 1981, 1989, 2008, 2017 гг.). Состояние естественного возобновления пород оценивалось путем учета подроста на площадках 4 м2 с привязкой к синузиям и микробиотопам в 1967, 2005, 2007, 2011 и 2017 гг. Возраст подроста определен по мутовкам. Структура видового разнообразия каждой синузии оценена по соотношению обилия видов эколого-ценотических групп (ЭЦГ) (Молокова, 1992; Назимова и др., 2012).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Пространственное строение черневого кедровника осочково-крупнотравно-папоротникового характеризуется сложностью и выраженной согласованностью динамики структурных компонентов. Особенности состава и структуры древостоя влияют на формирование пространственной мозаики нижних ярусов, что в свою очередь определяет особенности возобновительного процесса. В результате такого взаимодействия на протяжении жизни многих поколений ключевых видов происходит формирование устойчивого позднесукцессионного сообщества.
Древостой. Древостой имеет типичный для черневых лесов Западного Саяна смешанный кедрово-пихтовый состав, сложную вертикальную структуру и неравномерную сомкнутость крон (0.3–0.9) (рис. 1, 2). Изменения таксационных показателей древостоя за весь период наблюдения приведены в табл. 1.
Рис. 1.
Вертикальный профиль черневого кедровника осочково-крупнотравно-папоротникового на горном склоне крутизной 22°–25°.
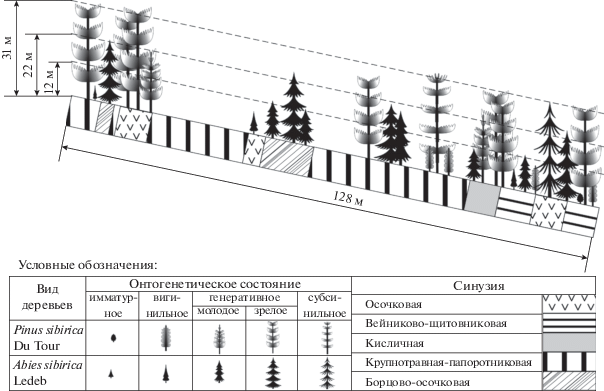
Рис. 2.
Горизонтальная структура древостоя черневого кедровника осочково-крупнотравно-папоротникового по данным 1972 г. (а); синузиальная структура травяного покрова в 1972 г. (б) и 2017 г. (в). Виды деревьев: 1 – сосна кедровая сибирская (Pinus sibirica Du Tour.); 2 – пихта сибирская (Abies sibirica Ledeb.); синузии: 3 – осочковая; 4 – вейниково-щитовниковая; 5 – кисличная; 6 – крупнотравно-папоротниковая; 7 – борцово-осочковая; 8 – крупнотравно-папоротниковая (лог).

Таблица 1.
Динамика основных показателей древостоя на пр.пл. 3 по ярусам
| Год | Ярус | Состав* | Hср, м | М, м3 га–1 | Полнота | N, шт. га–1 | |
|---|---|---|---|---|---|---|---|
| кедр | пихта | ||||||
| 1965 | I | 6K(193)3K(282)1П(136) + К(370) ед.Б(101) | 27.5 | 486 | 0.71 | 164 | 44 |
| II | 8П(87)1К(110)1П(43) | 13.9 | 27 | 0.13 | 13 | 269 | |
| 1990** | I | 7К(219)3К(306) + П(155), К(395) | 28.1 | 457 | 0.65 | 135 | 15 |
| II | 8П(107)2П(57) + К(145) | 16.4 | 45 | 0.17 | 8 | 287 | |
| 2017 | I | 7К(246)3К(333) + П(182) ед.К(422) | 31.4 | 482 | 0.75 | 128 | 8 |
| II | 9П(134)1К(172) | 22.3 | 60 | 0.22 | 8 | 129 | |
| III | 6П(74)3П(50)1Б(50) | 12.2 | 16 | 0.19 | – | 347 | |
Горизонтальная структура кедровника слагается чередованием плотных биогрупп деревьев и световых окон в пологе древостоя. Кедр по площади размещен группами (по 5 ± 1 шт. со смыканием крон) в форме линейных структур и куртин, приуроченных к микроповышениям рельефа, образованным ветровальными комплексами и старым валежом. Редкостойность и мозаичность горизонтальной структуры древостоя кедра связаны с особенностями его возобновления в черневых лесах, обусловленными высокой конкуренцией за свет и элементы минерального питания с мощно развитым травяным покровом из папоротников и крупнотравья (Назимова, Ермоленко, 1980). Пихтовая часть древостоя также имеет неравномерное размещение по площади (по 7 ± 1 шт. со смыканием крон), в связи с приуроченностью возобновления пихты к участкам вблизи крон крупных деревьев. Как результат образуются довольно крупные биогруппы деревьев кедра и пихты (по 10 ± 2 шт. деревьев первого яруса со смыканием крон) с высокой сомкнутостью крон (0.6–0.9). Незначительная часть площади (около 5–10%) занята разреженным пологом (сомкнутость 0.4–0.6). Окна в пологе древостоя занимают ровные или слегка пониженные участки с мощно развитым ярусом крупнотравья и папоротников.
За прошедший период наблюдения вертикальная структура древостоя усложнилась вследствие активного пополнения древостоя молодыми особями пихты (табл. 1). Следует отметить, что деление на ярусы (как и на поколения) сильно затруднено разновозрастностью структуры древостоя. Господствующий полог древостоя сложен тремя поколениями кедра (246, 333 и 422 лет) и старшим поколением пихты (182 лет). Во второй ярус древостоя входит второе поколение (134 лет) пихты и четвертое поколение кедра, самое неоднородное по своей возрастной структуре (172 лет). Третий ярус сформирован третьим (74 лет) и четвертым (50 лет) поколениями пихты, представленными молодыми генеративными и виргинильными деревьями.
Подлесок средней густоты, представлен двумя подъярусами (общая сомкнутость – 0.3). Первый подъярус разреженный (высотой 2–6 м): Sorbus sibirica, Padus avium, Sambucus racemosa. Второй подьярус более сомкнут, высотой 1–1.2 м, господствуют Spirea chamaedrifolia, S. media, с участием Rubus idaeus, Ribes nigrum. Распределение подлеска находится в прямой зависимости от состава и строения древесного яруса. В ходе естественной динамики древостоя локально при вывалах крупных кедров отмечается разрастание спиреи, караганы древовидной.
Травяной покров. Особенностью сезонного развития травяного яруса черневого кедровника осочково-крупнотравно-папоротникового является выраженный весенний аспект эфемероидов (Corydalis bracteata, Anemone altaica, A. reflexa, Viola uniflora, V. selkirkii) общим обилием 40–50%, чуть позже – с участием Pulmonaria mollis, Adoxa moshatellina, Allium microdiction, Anemone baicalensis. В этот период папоротники и виды крупнотравья только начинают вегетацию.
В начале лета (июнь) в травяном ярусе четко выделяются следующие синузии (экологически и флористически, структурно обособленные мозаики): крупнотравно-папоротниковая, вейниково-щитовниковая, борцово-осочковая, осочковая, кисличная. Уровень их сложности и степень приуроченности к эдификаторному ярусу различны, как и продолжительность существования во времени. Первое описание основных структурных элементов травяного яруса приведено в работе Т.С. Кузнецовой (1969).
Наиболее полно отражает условия местопроизрастания (экотоп) мощно развитая крупнотравно-папоротниковая синузия. Она представлена под разреженным пологом древостоя, в просветах и окнах верхнего полога (общее проективное покрытие (ОПП) до 100%, высота 80–120 см, сомкнутость 0.4 и менее). Доминируют: Athyrium filix-femina, Dryopteris expansa, Diplazium sibiricum, Aconitum septentrionale, Paeonia anomala, Crepis sibirica, Calamagrostis obtusata, Anemone baicalensis. С обилием Sol встречаются: Polystichum braunii, Dryopteris filix-mas, D. carthusiana, Euphorbia pilosa, Senecio nemorensis, Pleurospermum uralense, Delphinium elatum, Cacalia hastata, Pulmonaria mollis, Cruciata krylovii и др. Она наиболее богата по составу (41–45 видов, обильны борео-неморальные и неморальные гигромезофиты). Вес воздушно-сухой массы трав – 187.2 г м–2 (данные по надземной фитомассе синузий – по Т.С. Кузнецовой, 1969).
Вейниково-щитовниковая синузия приурочена к участкам c сомкнутостью крон (0.4–0.6) и близко расположенных выходов хлоритовых сланцев, где идет интенсивный вынос продуктов почвообразования и выветривания. В составе доминируют Dryopteris expansa и Calamagrostis obtusata, с участием таежного мелкотравья: Oxalis acetosella, Majanthemum bifolium, а также Gymnocarpium dryopteris, Phegopteris connectilis, Equisetum pratense, Milium effusum, Viola uniflora, Polemonium caeruleum и др. (ОПП 60–70%, высота 60–70 см). Число видов в составе синузии 30–35, это бореальные и борео-неморальные мезо- и гигромезофиты. Вес воздушно-сухой массы трав – 113.1 г м–2.
Борцово-осочковая и осочковая синузии приурочены к более сухим и слегка выпуклым участкам (под крупными кедрами), а также к участкам древостоя со вторым ярусом пихты. Структуру борцово-осочковой синузии формируют Carex macroura и Aconitum septentrionale (ОПП 60–70%, высота 70 (120) см). Общее число видов 20–26. Возрастает роль подлеска из Spiraea chamaedryfolia и S. media, Caragana arborescens, Sorbus sibirica. Вес воздушно-сухой массы трав – 75.7 г м–2. Осочковая синузия занимает подкроновые участки одиночных кедров и их групп с участием молодых пихт (Carex macroura с примесью видов таежного мелкотравья и теневых трав). Она представляет собой обедненный вариант борцово-осочковой синузии (ОПП 70–80%, высота 20–40 см, число видов 16–18, с высокой долей участия ксеромезофильной осочки). Вес воздушно-сухой массы трав – 67.9 г м–2.
Кисличная синузия (Oxalis acetosella, с незначительным участием теневых трав и таежного мелкотравья) развивается под сомкнутыми биогруппами пихт в условиях сильного затенения (ОПП 20–30%, высота 3–8 см). Здесь проявляется наибольшее влияние эдификаторного яруса. В отличие от остальных ранее рассмотренных синузий время ее существования ограничено немногими десятками лет, периодом изреживания и выхода крупного пихтового подроста в древесный ярус. Число видов, слагающих синузию, 11–13.
Фрагментарно на незначительной площади встречается крупнотравно-страусниковая синузия, приуроченная к вогнутому микрорельефу и более характерная для долинного кедровника крупнотравно-страусникового.
В ходе возрастной динамики насаждения (1965–2017 гг.) каждая синузия сохраняет относительное постоянство числа видов, состава доминантов, при варьировании обилия отдельных видов. Степень трансформации основных экологических и почвенных параметров определяется особенностями древостоя и его размещением по площади, различиями в составе, структуре и биомассе синузий. Это отражается на составе подстилок, температуре, влажности и химическом составе почв и других параметрах. По данным З.В. Вишняковой с соавторами (1968) и Т.С. Кузнецовой (1969) наименьшая весовая влажность почвы отмечается под кронами кедра в осочковой синузии (28–50%), что связано с их способностью задерживать атмосферные осадки. Крупнотравно-папоротниковая синузия приурочена к разреженным участкам верхнего полога, поэтому задержка влаги кронами минимальна (весовая влажность почвы 120–160%). Чем кислее подстилка, тем хуже идет разложение органического вещества. В осочковой синузии наиболее медленно разлагается опад кедра и осочки большехвостой (кислотность подстилки 4.5–5.0). Кислотность снижается при улучшении ее разложения, количество мобилизованного азота увеличивается. Глубина снегового покрова оказывает влияние на температурный режим почв и глубину ее промерзания. Более благоприятны условия для развития эфемероидов в крупнотравно-папоротниковой синузии (глубина снегового покрова 75 см).
Таким образом, в пределах одного эталонного участка типа леса (биогеоценоза) в сходных микробиотопах складываются устойчивые группировки растений одной формы ассоциирования, сходные по экологическим требованиям. Их сочетание формирует мозаичный травяной ярус. Наличие столь отличных синузий является показателем достаточно выраженной неоднородности экотопических условий, которая значима для травяного яруса, возобновления, но не играет определяющей роли для существующего на этом пространстве эдификаторного яруса.
Среди структурных элементов кедровника, усиливающих неоднородность и создающих разнообразие микробиотопов, отмечены редкие ветровально-почвенные комплексы, сформировавшиеся после вывалов с корнями старовозрастных кедров и пихт; единичных обломков стволов кедра и пихты (называемых “остолопами”), скоплений валежа на разных стадиях разложения и отдельных экземпляров сухостоя пихты. От размеров отдельного или группы отмерших деревьев зависит формирование окон. Крупные окна в кедровнике довольно длительное время сохраняют занимаемую площадь, поскольку мощно развитый покров из папоротников и видов лугово-лесного крупнотравья препятствует успешному возобновлению в них. Однако по периферии, а иногда и в центре окон на валеже формируются группы пихт и подлеска, которые могут явиться начальной стадией освоения пространства древесными видами.
С различиями в травяном покрове связано и развитие синузий мхов. Тонкий прерывистый слой борео-неморальных влаголюбивых мхов отмечается только в вейниково-щитовниковой и крупнотравно-папоротниковой синузиях (проективное покрытие до 20–40%): Plagiomnium cuspidatum, Cirriphyllum piliferum, Drepanocladus sp., Mnium sp., Bryum sp., Rhodobrium roseum, Climacium dendroides. В осочковой и борцово-осочковой синузиях моховой покров практически отсутствует. На полуразложившемся валеже формируют синузии главным образом таежные мхи (общая мощность слоя мхов 2–5 см): Rhytidiadelphus triquetrus, R. squarrosus, Hylocomium splendens, Ptilium crista-castrensis, Pleurozium schreberi. Присутствие валежа, покрытого зелеными мхами, играет важную роль в появлении следующей генерации кедра и пихты, хотя его площадь невелика (10–15% от общей поверхности почвы).
Пространственно-временная изменчивость структуры по данным картирования и анализа спектров ЭЦГ. Результаты повторного картирования (1972 и 2017 гг.) отражают изменения локализации синузий на пробной площади (рис. 2, табл. 2). Господствующая (основная) крупнотравно-папоротниковая синузия, связанная с наиболее разреженными участками верхнего полога, сохраняет видовой состав, структуру, долю занимаемой площади, но меняет отчасти контуры и местоположение. Наиболее заметны изменения мозаики осочковой и кисличной синузий. Пятна борцово-осочковой синузии местами трансформируются в осочковые. Carex macroura характеризуется активной стратегией, занимая новые участки у приствольных кругов, в подкроновых пространствах кедра и под пологом пихтовых биогрупп. Пихта, выходя во второй ярус, начинает активнее преобразовывать микросреду (изменяя световой режим, влажность и температуру воздуха и почв). Под группами густого пихтового подроста обозначаются мелкие участки кисличной синузии. По мере роста пихты и увеличения освещенности осочка и кислица вытесняются вейником и щитовником. Отмечена трансформация и других (вейниково-щитовниковой, борцово-осочковой) синузий, в которых меняется обилие и степень развития трав в годы повышенной влажности (разрастание борца) или периоды относительно сухого лета (“низкорослость” папоротников и крупнотравья, снижение надземной массы трав на 20–30%, по данным Т.С. Кузнецовой (1969) и собственным наблюдениям). Это не меняет принципиального хода естественной динамики, но временно перераспределяет роли отдельных ЭЦГ и видов-доминантов (разногодичные флюктуации фитоценоза в связи с погодными особенностями лет).
Таблица 2.
Динамика доли площадей синузий (%) в кедровнике осочково-крупнотравно-папоротниковом
| Синузия | Доля площади синузий, % | ||
|---|---|---|---|
| 1967 г. | 1972 г. | 2017 г. | |
| Крупнотравно-папоротниковая | 33 | 31 | 38 |
| Вейниково-щитовниковая | 13 | 19 | 12.5 |
| Борцово-осочковая | 20.5 | 12.5 | 13 |
| Осочковая | 19* | 23.5 | 20 |
| Кисличная | 6.5 | 6 | 7.5 |
| Крупнотравно-папоротниковая (лог) | 8 | 8 | 9 |
Соотношение мозаик синузий по площади (по результатам картирования 1967, 1972 и 2017 гг.) несколько колеблется, но в целом остается стабильным в течение 50 лет: сохраняется господство крупнотравно-папоротниковой (основной) синузии (33, 31, 38%), при заметной доле вейниково-щитовниковой, с преобладанием теневых мезофильных трав (13, 19 и 12.5%) и осочковой, наиболее ксеромезофильной (19, 23.5 и 20%). Переходная борцово-осочковая синузия год за годом уменьшается по площади (20.5, 12.5 и 13%) в пользу основной. Выявленная картина логична и говорит об устойчивости фитоценотической структуры черневого кедровника в отсутствие резких погодно-климатических колебаний, в том числе засух. Последние не свойственны избыточно влажному климату черневого пояса.
На протяжении 50 лет естественного развития кедровника осочково-крупнотравно-папоротникового все отмеченные синузии, различные по функциям в биогеоценозе, остаются постоянными элементами структуры одного типа леса (климаксовые, или коренные синузии по Т.А. Комаровой (1992)). Динамика популяции лесообразователей (появление окон, вывалов, рост биогрупп пихты) играет ведущую роль на части пространства, изменяя синузиальную структуру нижних ярусов и верхние горизонты почвенных профилей. К ним полностью приложимо понятие парцелла, введенное Н.В. Дылисом (1969). В молодых пихтовых биогруппах парцеллярность проявляется более четко. В отличие от них кедровый подрост не образует явных биогрупп, и вместо парцелл формируются лишь синузии травяного яруса. Почвенно-гидрологические факторы имеют решающее влияние на формирование этой неоднородности.
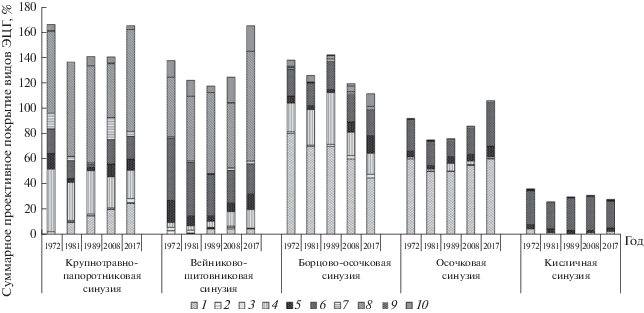
Соотношение эколого-ценотических спектров видов травяного покрова дает расширенное представление о разногодичной изменчивости и постоянстве состава ЭЦГ в кедровнике. Каждая синузия на протяжении почти полувекового периода наблюдения сохраняет свой видовой состав, соотношение ЭЦГ в спектре и состав доминантов (рис. 3). Эдификаторная крупнотравно-папоротниковая синузия, развиваясь в условиях наибольшего освещения, сохраняет господство видов лугово-лесного крупнотравья и крупных лесных папоротников с участием неморальных реликтов, нарастанием доли осочки к 2017 г. Вейниково-щитовниковая синузия, развитая под более сомкнутым пологом кедра и пихты, представлена щитовником и вейником тупоколосковым с заметным участием видов таежного мелкотравья. Борцово-осочковая синузия сложена видами лугово-лесного крупнотравья и осочкой большехвостой. Ее обедненный вариант – осочковая синузия – приурочена к более сухим и выпуклым участкам микрорельефа и подкроновым участкам кедров. Кисличная синузия сохраняет доминирование таежного мелкотравья с незначительным участием теневых трав. Отдельные колебания долей ЭЦГ в спектре во времени связаны как с незначительными трансформациями синузий, так и с погодными флюктуациями.
Рис. 3.
Изменение спектров ЭЦГ видов в синузиях. 1 – группа осочки; 2 – группа боровых видов; 3 – группа лугового и лугово-лесного разнотравья и злаков; 4 – лугово-лесное крупнотравье; 5 – теневые травы; 6 – таежное мелкотравье; 7 – неморальные реликты; 8 – крупные лесные папоротники; 9 – травяно-болотные виды; 10 – группа вейника тупоколоскового.
Возобновление кедра и пихты. Этот вопрос заслуживает особого внимания и изучался достаточно подробно (Бабинцева, 1966; Кедровые …, 1985; Ермоленко, 1981, 1991; и др.). В данной статье показана типичная для всех черневых кедровников картина слабого естественного возобновления, получившая на пр. пл. 3 подтверждение в течение всего срока мониторинга (50 лет). Обилие подроста кедра в среднем по годам составило 399 ± 57 шт. га–1. Подрост пихты преобладал по численности (1309 ± 72 шт. га–1) во все годы учета.
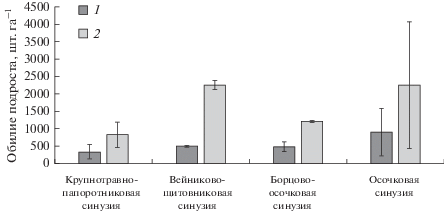
Мозаика микробиотопических условий и горизонтальной структуры нижних ярусов растительности определяют неравномерность размещения молодого поколения древесных видов (рис. 4). Особенно неравномерно расположен по площади подрост кедра (встречаемость 37%), в отличие от пихты (62% встречаемости).
Рис. 4.
Густота возобновления кедра (1) и пихты (2) (средняя за период наблюдений) в синузиях черневого кедровника осочково-крупнотравно-папоротникового.
В целом в горно-черневом кедровнике осочково-крупнотравно-папоротниковом наиболее благоприятными структурными элементами для поселения кедрового и пихтового подроста являются вейниково-щитовниковая, борцово-осочковая и осочковая синузии травяного яруса, связанные с участками сомкнутых куртин пихтово-кедрового древостоя и приствольными кругами единичных крупных деревьев кедра. Однако выживанию молодых поколений лесообразователей препятствует корневая и световая конкуренция со стороны материнского полога, особенно под пологом плотных биогрупп пихты. В связи с этим происходит постоянная гибель подроста: кедра в возрасте 5–10, пихты – 20–40 лет. Как отмечал П.М. Ермоленко (1991), в кедровниках отпад подроста кедра достигает 40–60%, но восполнение подроста значительно превышает его отпад. Большая возобновительная способность и теневыносливость пихты позволяет ей постепенно внедряться в основной полог древостоя. И только после формирования и дальнейшего распада пихтового древостоя происходит переход выжившего подроста кедра в древостой. Таким образом, непрерывное образование молодых поколений кедра в борцово-осочковой и осочковой синузиях представляет собой постоянно обновляющийся резерв возобновления, переходящего в древостой только в случае распада господствующего полога древостоя. В то же время в вейниково-щитовниковых и крупнотравно-папоротниковых синузиях под относительно разреженным пологом древостоя и в окнах формирование новых поколений коренных (позднесукцессионных) пород приурочено исключительно к ветровальным комплексам и крупному валежу, образующимся при отпаде кедра, достигшего возраста 260–300 лет. Как следствие, для кедра характерно чередование условно-разновозрастных, а для пихты – абсолютно-разновозрастных поколений.
Постепенный распад перестойных поколений кедра улучшает условия роста оставшихся поколений лесообразователей и влечет за собой образование новых молодых поколений леса.
Исследования на экспериментальных пробных площадях в других коренных типах леса горно-черневого пояса позволяют прийти к некоторым обобщениям. Высокий уровень биоразнообразия, многокомпонентность состава, сосуществование разных эколого-ценотических групп видов, формирующих устойчивые синузии, а также видов с разными стратегиями выживания – все это вместе обеспечивает устойчивость позднесукцессионного лесного биогеоценоза в условиях неравновесности среды обитания в горах. Имеет большое значение, в частности, высокая толерантность кедра к колебаниям гидротермических условий, способность к адаптации в случаях воздействия насекомых, грибных заболеваний, повреждения ветром и другими неблагоприятными факторами. Основным фактором риска для кедра в черневом поясе является конкуренция за свет со стороны трав, не позволяющая ему выжить на первых стадиях роста и развития. Шанс появиться подросту и выжить предоставляется в случае появления окон (после буреломов и ветровалов) и заноса семян в подходящие микробиотопы с разреженным травяным покровом и пятнами мхов. С этих позиций становится более понятной ведущая роль разнообразия микробиотопов и наличия подходящих синузий для поддержания жизнеспособности популяции кедра как эдификатора.
Заключение. Подведены итоги стационарного изучения пространственно-временной структуры и динамики кедровника осочково-крупнотравно-папоротникового (1965–2017 гг.), широко распространенного в черневом поясе Западного Саяна. Полученные результаты позволяют обсуждать и прогнозировать возрастную динамику малонарушенного черневого кедровника с учетом развития верхнего кедрового полога древостоя, роста биогрупп пихты, особенностей формирования окон, вывалов, разнообразия микробиотопов и синузий нижних ярусов в горных условиях. Исследованный участок леса может рассматриваться как эталонный биогеоценоз, не затронутый рубками и пожарами. Его пространственная структура сформирована в ходе многовековой сукцессии и дает образец устойчивости во времени. Постоянство его функционирования поддерживается разновозрастной структурой эдификаторного яруса и способностью кедрового подроста долго выдерживать световую конкуренцию трав и пихты, накапливать свою численность и формировать несколько поколений под материнским пологом. Наряду с парцеллярной структурой, определяемой возрастной мозаикой пихтового подроста и биогруппами взрослых кедров, в биогеоценозе четко фиксируется неоднородность (синузиальность) травяного покрова, связанная с различиями микробиотопов. Ослабленная зависимость от эдификатора при более тесной связи с почвенно-гидрологическим режимом позволяет структурным элементам травяного покрова поддерживать устойчивость (резистентность) биогеоценоза. Это ключ к пониманию устойчивости многокомпонентного черневого кедровника на протяжении длительного времени.
На примере изученного кедровника показано, что тип леса в горных условиях всегда имеет внутренне неоднородную структуру, которая может быть раскрыта в полном объеме лишь на постоянных участках, охватывающих достаточно крупную площадь с однородным древостоем, в границах одного типа лесорастительных условий. ЭЦГ видов играют роль функциональных звеньев, реагирующих на изменения внешних и внутренних факторов среды. Выявлена связь основных элементов мозаики нижних ярусов с возрастной структурой эдификаторного яруса. Синузии травяного покрова (крупнотравно-папоротниковая, вейниково-щитовниковая, борцово-осочковая, осочковая, кисличная) остаются постоянными элементами фитоценотической структуры одного типа леса на протяжении полувекового периода и дольше.
Список литературы
Бабинцева Р.М. Природа возобновительного процесса в кедровых лесах Западного Саяна: Автореф. дисс. … канд. с.-х. наук: 06.03.03. Красноярск: Изд-во Института леса, 1966. 21 с.
Вишнякова З.В., Зуева К.Г., Кузнецова Т.С., Чагина Е.Г. О взаимодействии леса и почвы // Лес и почва. Тр. Всесоюз. науч. конф. по лесному почвоведению (15–19 июля 1968). Красноярск: Красноярское книж. изд-во, 1968. С. 494–531.
Восточноевропейские леса: история в голоцене и современность / Отв. ред. Смирнова О.В. М.: Наука, 2004. Кн. 2. 575 с.
Дылис Н.В. Структура лесного биогеоценоза. М.: Наука, 1969. 55 с.
Дыренков С.А. Структура еловых древостоев в юго-западной части Вычегодско-Мезенской равнины // Ботанический журн. 1966. № 8. С. 1149–1156.
Дыренков С.А. Структура и динамика таежных ельников. Л.: Наука, 1984. 174 с.
Ермоленко П.М. Высотно-поясные особенности роста кедра и пихты в Западном Саяне // Формирование и продуктивность древостоев. Новосибирск: Наука, 1981. С. 19–53.
Ермоленко П.М. Возрастная и восстановительная динамика структуры темнохвойных лесов Западного Саяна // Теория лесообразовательного процесса: Тезисы докладов Всесоюзного совещания. Красноярск: Изд-во КНЦ СО АН СССР, 1991. С. 48–50.
Кедровые леса Сибири / Отв. ред. Исаев А.С. Новосибирск: Наука, 1985. 256 с.
Комарова Т.А. Послепожарные сукцессии в лесах Южного Сихотэ-Алиня. Владивосток: Изд-во ДВО АН СССР, 1992. 224 с.
Кузнецова Т.С. Фитоценотическая структура кедровников Западного Саяна // Типы лесов Сибири. Красноярск: Изд-во ИЛ СО РАН, 1969. Вып. 2. С. 25–77.
Лащинский Н.Н. Структура и динамика сосновых лесов Нижнего Приангарья. Новосибирск: Наука, 1981. 272 с.
Молокова Н.И. Эколого-ценотический анализ и феноиндикация высотно-поясных комплексов типов леса (на примере гумидных районов Саян): Автореф. дис. … канд. биол.наук: 03.00.16, 03.00.5. Красноярск: Институт леса СО РАН, 1992. 16 с.
Москалюк Т.А. Ценотическая структура лиственничников зеленомошных на юге Магаданской области // Лесоведение. 2012. № 3. С. 19–32.
Назимова Д.И., Ермоленко П.М. Динамика синузиальной структуры при восстановительных сукцессиях в черневых кедровниках Западного Саяна // Динамика лесных биогеоценозов Сибири. Новосибирск: Наука, 1980. С. 54–87.
Назимова Д.И., Дробушевская О.В., Данилина Д.М., Коновалова М.Е., Кофман Г.Б., Бугаева К.С. Биоразнообразие и динамика низкогорных лесов Саян: региональный и локальный уровни // Разнообразие и динамика лесных экосистем России. В 2-х кн. М.: Товарищество научных изданий КМК, 2012. Кн. 1. С. 131–172.
Назимова Д.И., Коновалова М.Е., Данилина Д.М., Пономарев Е.И., Сташкевич Н.Ю., Бабой С.Д. Исследования долговременной динамики лесов в пергумидном климате Западного Саяна (Ермаковский стационар Института леса им. В.Н. Сукачева СО РАН) // Сибирский лесной журн. 2015. № 4. С. 3–17.
Поляков В.И. Черневые кедровники Западного Саяна: контроль и прогнозирование хода роста. Новосибирск: Изд-во СО РАН, 2007. 181 с.
Рубцов М.В., Рыбакова Н.А. Динамика парцеллярной структуры лесных фитоценозов в процессе восстановления популяции ели в южнотаежных березняках // Лесоведение. 2016. № 5. С. 323–331.
Санников С.Н., Санникова Н.С., Петрова И.В. Очерки по теории лесной популяционной биологии. Екатеринбург: Изд-во УрО РАН, 2012. 272 с.
Семечкин И.В. Структура и динамика кедровников Сибири. Новосибирск: Изд-во СО РАН, 2002. 253 с.
Сукачев В.Н. Основы лесной типологии и биогеоценологии. Л.: Наука, 1972. Т. 1. 420 с.
Сукачев В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: Изд-во АН СССР, 1961. 144 с.
Типы лесов гор Южной Сибири. Новосибирск: Наука, 1980. 336 с.
Bakker F.A. Why do we need permanent plots in the study of long-term vegetation dynamics? // J. Vegetation Science. 1996. V. 7. P. 145–156.
Bilek L., Remes J., Zahradnik D. Managed vs. unmanaged. structure of beech forest stands (Fagus sylvatica L.) after 50 years of development, Central Bohemia // Forest Systems. 2011. V. 20. Is. 1. P. 122–138.
Closset-Kopp D., Schnitzler A., Aran D. Dynamics in natural mixed-beech forest of the Upper Vosges // Biodiversity & Conservation. 2006. V. 15. P. 1063–1093.
Ismailova D.M., Nazimova D.I. Long-term dynamics of mixed fir-aspen forests in West Sayan (Altai-Sayan Ecoregion) / Environmental Change in Siberia: Earth Observation, Field Studies and Modelling, Advances in Global Change Research. London: Springer Science + Business Media B.V., 2010. P. 37–51.
Korb J.E., Ranker T.A. Changes in stand composition and structure between 1981 and 1996 in four Front Range plant communities in Colorado // Plant Ecology. 2001. V. 157. P. 1–11.
Nazimova D.I., Danilina D.M., Stepanov N.V. Rain-Barrier Forest Ecosystems of the Sayan Mountains // Botanica Pacifica. 2014. V. 3(1). P. 39–47.
Vacek S., Cerny T., Vacek Z., Podrazsky V., Mikeska M., Kralicek I. Long-term changes in vegetation and site conditions in beech and spruce forests of lower mountain ranges of Central Europe // Forest Ecology & Management. 2017. V. 398. P. 75–90.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение