

Лесоведение, 2020, № 5, стр. 433-441
Вертикально-фракционная структура надземной фитомассы древесного яруса сосняков Северного Предуралья
И. Н. Кутявин *
Институт биологии ФИЦ Коми НЦ УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: kutjavin-ivan@rambler.ru
Поступила в редакцию 05.04.2018
После доработки 26.10.2018
Принята к публикации 06.06.2020
Аннотация
Приводятся результаты вертикального распределения фитомассы надземных органов спелых и перестойных древостоев среднетаежных сосняков Северного Приуралья. Показаны возможности математического описания распространения отдельных фракций фитомассы древесных растений по вертикальному профилю древесного яруса. Надземная часть фитомассы сосновых древостоев в зависимости от типа леса располагается на высоте от 17 до 26 м, где основная масса органического вещества приходится на стволовую древесину, концентрирующуюся в слое 0–4 м. Протяженность кронового слоя деревьев в сосняках разных типов изменяется от 15 до 26 м. Основная масса ассимиляционного аппарата аккумулируется в толще 8–24 м. Индекс листовой поверхности в древостоях сосняков разных условий произрастания составляет от 4.9 до 5.2 га га–1. Вертикальный профиль распределения зависит от структуры и состава древостоев сосняков, хотя его максимум в большинстве типов сосняков свойственен верхней части полога.
Вертикально-фракционная структура фитомассы является основной при оценке энергомассообмена в лесных биогеоценозах. Сведения о вертикальном распределении фитомассы и ее продукции имеют большую значимость при определении биосферной, климаторегулирующей функций леса (Уткин, Дылис, 1966; Алексеев, 1975; Гульбе с соавт., 1983; Вертикально-фракционное …, 1986; Stenberg et al, 1994; Cermak et al, 1998; Усольцев, 2013; и др.). В последнее время немаловажное значение вертикально-фракционная структура древесного полога представляет при проведении дистанционного зондирования земной поверхности (Korpela еt al., 2007; Данилин, Фаворская, 2013; Vauhkonen, Mehtätalo, 2014; и др.), математического моделирования, динамики пространственной структуры и функционирования лесных сообществ (Пулы …, 2007; Stinson et al., 2011; Shanin et al., 2014). Следовательно, для получения этих знаний необходимы данные о запасах органического вещества и его распределении в пространстве лесных насаждений.
Сосновые леса на европейском Севере России занимают порядка 32% лесопокрытой площади (Лесной …, 1999) и тем самым выполняют важные продуцирующие функции в составе фитомассы лесных сообществ. Биологическая продуктивность сосняков европейского Севера освещена в работах (Молчанов, 1971; Смирнов, 1971; Казимиров и др., 1977; Бобкова, 1987; Лукина, Никонов, 1996; Феклистов и др., 2015; Кутявин, Бобкова, 2017; и др.). Однако работы по вертикально-фракционному распределению фитомассы деревьев и древостоев в сосняках рассматриваемого региона практически отсутствуют. В работах P. Stenberg с соавт. (1994), J. Cermak с соавт. (1998) основное внимание уделяется анализу кронового пространства полога сосновых древостоев. Большая роль исследованиям пространственной вертикальной структуры древостоев отведена в работе В.А. Усольцева (2013). Автором проанализированы регрессионные модели вертикально-фракционного распределения фитомассы древостоя сосняков различного происхождения в зависимости от возраста, морфологической структуры дерева и древостоев сосняков лесной зоны Казахстана. В Республике Коми анализ вертикального распределения фитомассы древостоев проведен только в ельниках (Бобкова и др., 2000).
Цель данной работы состояла в изучении вертикально-фракционной структуры надземной фитомассы древостоев основных типов среднетаежных сосняков Северного Предуралья.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проведены в юго-восточной части Республики Коми бассейна верхней и средней Печоры в подзоне средней тайги. Для оценки биологической продуктивности старовозрастных сосняков было заложено четыре постоянных пробных площади (пр.пл.). Лесоводственно-таксационная характеристика исследованных сосняков представлена в табл. 1. Спелые и перестойные сосняки формируют как чистые, так и смешанные по составу древостои IV–Va классов бонитета. Для них характерна высокая дифференциация деревьев по возрасту. Древостои состоят из нескольких поколений сосны с максимальным возрастом деревьев 370 лет.
Таблица 1.
Таксационная характеристика старовозрастных сосняков
| Состав (ярус) | Средние | Возраст, лет | Густота, шт. га–1 | Запас стволовой древесины, м3 га–1 | Класс бонитета | Надземная фитомасса древостоя, т га–1* | |
|---|---|---|---|---|---|---|---|
| диаметр, см | высота, м | ||||||
| Сосняк лишайниковый (пр.пл. 11) | |||||||
| 10Cед.Б (I) | 26 | 13.8 | 40–220 | 411 | 163 | Va | 92.6 |
| Сосняк бруснично-лишайниковый (пр.пл. 9A) | |||||||
| 10C (I) | 14.3 | 14 | 56–370 | 1192 | 177.1 | V | 99.4 |
| Сосняк черничный (пр. пл. 10) | |||||||
| 10C (I) | 25 | 21.6 | 101–320 | 473 | 286 | IV | 146.6 |
| 7Е3С+Б (II) | 12.8 | 12.6 | 101–160 | 250 | |||
| Сосняк чернично-сфагновый (пр.пл. 9) | |||||||
| 8С1Е1Б+К (I) | 16 | 11.5 | 60–210 | 1020 | 109 | Va | 56.8 |
Массу органического вещества (ОВ) древостоев определяли по методу модельных деревьев (Уткин, 1975; Repola, 2006). На каждой пробной площади было проанализировано 5–8 деревьев из разных ступеней толщины древостоя. Такое число модельных деревьев обеспечивает достоверную оценку запасов и продукции органического вещества в насаждениях. Срубленные модельные деревья разделялись на двух метровые секции. В каждой секции кронового пространства проводилась обрубка ветвей с разделением их на фракции: сухие (отмершие), неохвоенные и охвоенные. Во всех секциях из охвоенной части отбиралась навеска 300–400 г, которая разбиралась на побеги и хвою, с учетом их возраста. Масса ствола по секциям взвешивалась на месте. Данные по фитомассе и структуре исследуемых сосняков нами опубликованы ранее (Кутявин и др., 2016; Кутявин; Бобкова, 2017). Модельные деревья послужили основой для изучения вертикально-фракционного распределения фитомассы древостоев. Так, по двух метровым секциям модельных деревьев строились графики зависимости массы отдельных фракций (стволовая древесина, кора, ветви, хвоя) от диаметра ствола. По полученным уравнениям зависимости для каждой фракции расчитывалась общая масса древостоя по секциям с использованием перечетных данных всех деревьев на пробной площади. После того, как для каждого слоя была получена фракционная масса, строились общие зависимости вертикального распределения фитомассы древостоя. Оценочные коэффициенты регрессионных уравнений и их погрешности вертикального распределения фитомассы в древостоях сосняков получены при помощи программ Curve Expert 1.4 и Excel 2007. Данные наземной фитомассы сосняков приведены в абсолютно сухом состоянии. Листовой индекс древостоя рассчитан на основе данных массы хвои и коэффициентов, которые представляют поверхность 1 г хвои (Бобкова, 1987). Так, масса 1 г сосновой хвои в свежем состоянии имеет поверхность – 56.97, в абсолютно сухом – 135.6 см2, еловой хвои – 50.41 и 109.6 см2, листья березы – 60.5 и 163.5 см2 соответственно.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
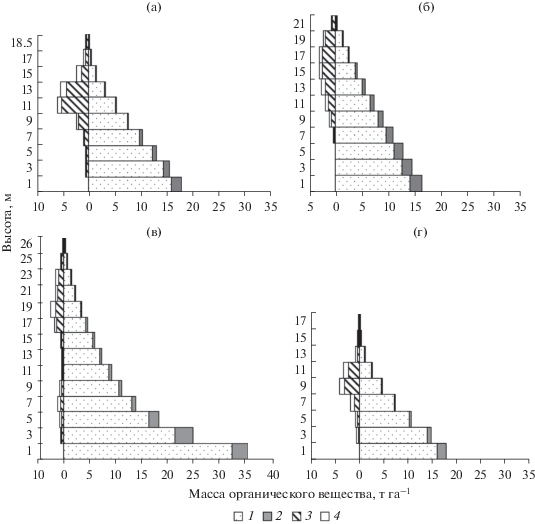
Общие запасы фитомассы ОВ надземных органов древостоев сосняков разных типов изменяются в пределах 56.8 до 146.6 т га–1 (табл. 1). Вертикально-фракционное распределение фитомассы древостоев в спелых и перестойных сосняках представлено в абсолютных единицах массы отдельных фракций. Эти показатели приведены в виде профильных диаграмм (рис. 1). Вертикальный профиль фитомассы древесного яруса среднетаежных сосняков характеризуется следующей протяженностью. Так, деревья в сосняке лишайниковом располагаются в слое высотой 19 м, бруснично-лишайниковом – 21, черничном – 26 м и чернично-сфагновом – 17 м.
Рис. 1.
Вертикальное распределение фитомассы по фракциям в сосняках: а – лишайниковом, б – бруснично-лишайниковом, в – черничном, г – чернично-сфагновом. Фракции фитомассы: 1 – стволовая древесина, 2 – кора стволовая, 3 – ветви, 4 – хвоя.
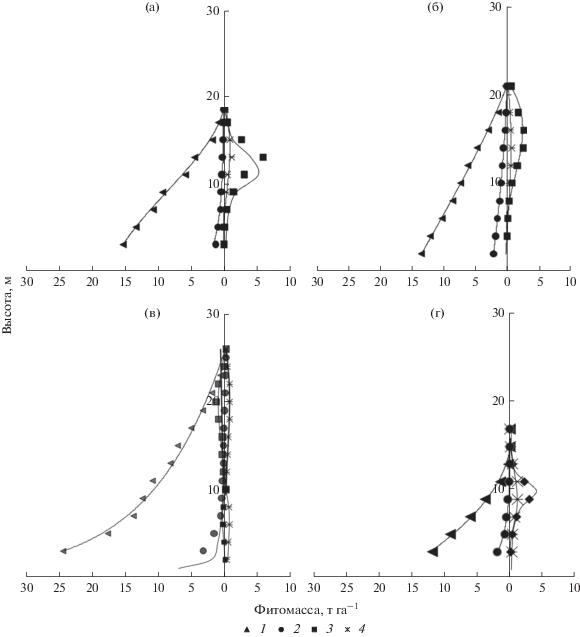
Вертикальное распределение отдельных фракций фитомассы древостоев получено с использованием регрессионных уравнений с аппроксимацией методом наименьших квадратов для каждой из функций (табл. 2). Для каждого уравнения получены коэффициенты a, b, c, d. Выбор уравнения осуществлялся по наименьшему значению стандартной ошибки (SEE) и достоверности аппроксимации тренда (R2). На основе статистического анализа распределение фитомассы отдельных фракций деревьев описывается полиномом третьего порядка y = a + bx + cx2+ dx3 и рациональной функцией – y = (a + bx)/(1 + cx + dx2). В сложном двухъярусном древостое сосняка черничного с целью упрощения кривых вертикального распределения фитомассы первого и второго ярусов были получены отдельные уравнения для хвои и ветвей. Кривые аппроксимации эмпирических данных вертикального распределения надземной фитомассы древостоев по фракциям получены при помощи функций (рис. 2).
Таблица 2.
Коэффициенты регрессионных уравнений вертикального распределения фитомассы по фракциям в сосновых фитоценозах*
| Фракция | Уравнение | Коэффициент | SEE | R2 | |||
|---|---|---|---|---|---|---|---|
| a | b | c | d | ||||
| Сосняк бруснично-лишайниковый (пр.пл. 9А) | |||||||
| Древесина стволовая | y = a + bx + cx2 + dx3 | 14.916 | –0.760 | –0.004 | 0.000 | 0.304 | 0.997 |
| Кора стволовая | y = a + bx + cx2 + dx3 | 2.481 | –0.184 | 0.003 | 0.000 | 0.051 | 0.997 |
| Ветви | y = a + bx + cx2 + dx3 | 2.015 | –0.907 | 0.118 | 0.004 | 0.280 | 0.952 |
| Хвоя (листья) | y = a + bx + cx2 + dx3 | –0.356 | 0.075 | 0.004 | 0.000 | 0.082 | 0.993 |
| Сосняк лишайниковый (пр. пл. 11) | |||||||
| Древесина стволовая | y = a + bx + cx2 + dx3 | 16.482 | –0.519 | –0.086 | 0.004 | 0.000 | 0.996 |
| Кора стволовая | y = aebx | 2.419 | –0.237 | – | – | 0.048 | 0.955 |
| Ветви | y = (a + bx)/(1 + cx + dx2) | 0.159 | –0.002 | –0.166 | 0.007 | 0.306 | 0.984 |
| Хвоя (листья) | y = a*exp((–(b – x)2)/(2*c2)) | 1.141 | 12.331 | 2.501 | – | 0.128 | 0.922 |
| Сосняк черничный (пр. пл. 10) | |||||||
| Древесина стволовая | y = a + bLnx | 32.029 | –9.781 | – | – | 0.862 | 0.988 |
| Кора стволовая | y = a + b/x | –0.212 | 7.430 | – | – | 0.100 | 0.990 |
| (I)** Ветви | y = a + bx + cx2 + dx3 | 2.750 | –0.768 | 0.065 | –0.002 | 0.268 | 0.839 |
| (II) Ветви | y = a + bx + cx2 + dx3 | 0.274 | –0.060 | 0.032 | –0.003 | 0.142 | 0.858 |
| (I) Хвоя (листья) | y = a + bx + cx2 + dx3 | 1.690 | –0.428 | 0.034 | –0.001 | 0.078 | 0.922 |
| (II) Хвоя (листья) | y = a + bx + cx2 + dx3 | 0.167 | –0.098 | 0.051 | –0.004 | 0.002 | 0.999 |
| Сосняк чернично-сфагновый (пр.пл. 9) | |||||||
| Древесина стволовая | y = a + bx + cx2 | 18.109 | –2.238 | 0.069 | – | 0.340 | 0.996 |
| Кора стволовая | y = a + bx + cx2 + dx3 | 3.921 | –0.896 | 0.071 | –0.002 | 0.150 | 0.968 |
| Ветви | y = (a + bx)/(1 + cx + dx2) | 0.156 | –0.008 | –0.197 | 0.010 | 0.080 | 0.998 |
| Хвоя (листья) | y = abxxc | 0.00001 | 0.315 | 10.345 | – | 0.203 | 0.922 |
Рис. 2.
Выровненные по уравнениям регрессионные модели распределения фитомассы по вертикальному профилю в древостоях сосняков: а – лишайниковом, б – бруснично-лишайниковом, в – черничном, г – чернично-сфагновом. Фракции фитомассы: 1 – стволовая древесина, 2 – кора стволовая, 3 – ветви, 4 – хвоя.
Так, с улучшением эдафических условий в сосняках происходит увеличение запасов органической массы стволовой древесины (рис. 1). Основная доля данного компонента располагается в первых двух слоях (на высоте 0–4 м), здесь же концентрируется значительная часть коры. Более высокие показатели массы стволовой древесины в этом слое отмечаются в сосняке черничном и составляют 69.9 т га–1. В сосняках лишайниковом и бруснично-лишайниковом масса стволовой древесины в слое 0–4 м примерно в два раза меньше, чем в сосняке черничном, и составляет 30.1 и 26.8 т га–1, соответственно. В сосняке чернично-сфагновом масса стволовой древесины этого слоя равна 29.1 т га–1.
Кроновое пространство сосняков разных типов характеризуется протяженностью от 15 до 26 м. Большая протяженность крон характерна для древесного яруса, в формировании которого участвуют деревья разных поколений и входящих в состав древостоя сопутствующих пород (кедр, ель, береза). В сосняке черничном древостой образует два яруса, где второй ярус состоит преимущественно из ели. Общая протяженность крон деревьев второго яруса в данном сосняке составляет 10 м. Первый ярус имеет протяженность от 12 до 26 м. Большая масса ветвей в сосняке лишайниковом расположена в слое от 10 до 14 м, бруснично-лишайниковом – от 10 до 20, черничном – от 16 до 22 и чернично-сфагновом – от 8 до 12 м. Следует отметить, что с ухудшением эдафических условий по мере увеличения влажности и сухости почв масса ветвей в кроновом слое возрастает, тогда, как в более продуктивном сосняке черничном основная масса приходится на стволовую древесину (рис. 2).
При оценке вертикально-фракционной структуры древостоя большой интерес представляет биогоризонт, где располагаются ассимилирующие органы древесных растений (Ладанова, Тужилкина, 1992; Stenberg еt al, 1994; Cermak еt al, 1998; Биопродукционный …, 2001; и др.). Биогоризонты с высотной концентрацией массы ассимиляционного аппарата являются основными аккумуляторами органического вещества. Так, преобладающая часть массы хвои в сосняке лишайниковом сосредоточена в слое от 12 до 16 м, бруснично-лишайниковом – от 10 до 18 м, чернично-сфагновом – от 6 до 14 м. В сосняке черничном масса хвои сосредоточена в двух горизонтах – на высоте от 6 до 8 м и от 16 до 24 м.
Листовой индекс – отношение поверхности листьев к единице площади – один из показателей, характеризующих продукционный процесс лесных фитоценозов. По расчетам, проведенным нами, наибольший показатель листового индекса древесных растений старовозрастных сосняков характерен для лишайникового типа – 5.19 га га–1. Остальные типы сосняков показали схожие показатели индексов листовой поверхности, с изменением от 4.88 до 4.92 га га–1 (рис. 3). По данным других исследований в среднетаежных сосняках черничных данный показатель может изменяться от 5.4 до 10.3 га га–1(Бобкова, 1987; Биопродукционный …, 2001).
Рис. 3.
Вертикальное распределение коэффициента листового индекса в сосняках: а – лишайниковом, б – бруснично-лишайниковом, в – черничном, г – чернично-сфагновом. Для каждого типа сообществ указан общий листовой индекс, га га–1.
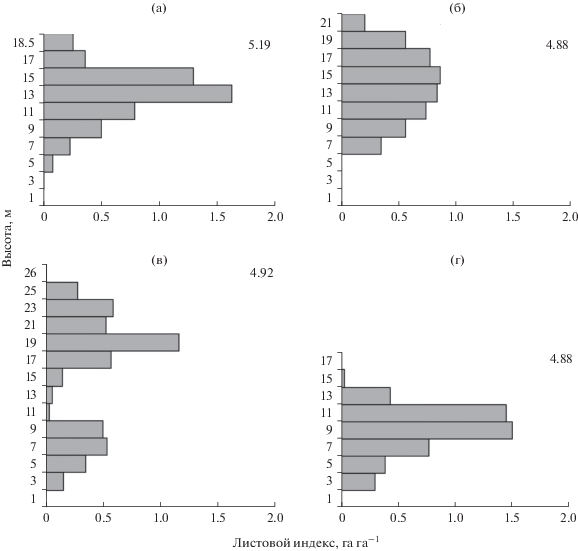
Как показывают данные рис. 3, большая часть хвои (листьев) в сосняке лишайниковом расположена на высоте от 12 до 16 м, с общей площадью поверхности листьев 2.9 га га–1. В бруснично-лишайниковом древостое ассимиляционный аппарат деревьев (4.9 га га–1) располагается по высоте кроны равномерно (без скачков) на протяжении всего кронового полога с максимальной концентрацией в слое 10–18 м. В двухъярусном сосняке черничном концентрация листовой поверхности размещена крайне неравномерно. Основная площадь ее поверхности располагается в слое от 16 до 24 м с суммарным индексом 3.1 га га–1. Во втором ярусе, состоящем преимущественно из ели и березы, основная площадь листовой поверхности концентрируется на высоте от 4 до 10 м. Чернично-сфагновый сосняк аккумулирует основную площадь листовой поверхности в слое от 6 до 12 м с суммарным индексом 3.7 га га–1, что составляет 76% от суммарной площади листовой поверхности. Такое неравномерное распределение ассимиляционных органов в заболоченном сосняке объясняется участием в составе соснового древостоя ели, которая в свою очередь обладает относительно большой массой и высокой концентрацией хвои в пологе.
ВЫВОДЫ
1. Вертикальный профиль спелых и перестойных сосняков разных типов различен. Так, в сосняке лишайниковом фитомасса располагается в слое 19 м, бруснично-лишайниковом – 21, черничном – 26, чернично-сфагновом – 17 м.
2. Протяженность крон по вертикали в сосняках разных типов составляет от 15 до 24 м, где основная масса ветвей и хвои сосредоточена в слое 8–24 м. Индекс листовой поверхности в перестойных среднетаежных сосняках Северного Приуралья изменяется от 4.9 до 5.2 га га–1. Большая площадь листовой поверхности характерна для древостоя сосняка черничного, с наличием в его составе темнохвойных древесных видов второго яруса, где древостой представлен двумя максимумами листовой поверхности на высотах 8–10 и 18–20 м.
3. Вертикальное распределение массы фракций отдельных частей дерева не имеет четкой выраженности в определенной функции, но наиболее часто описывается полиномом третьего порядка и рациональной функцией.
Список литературы
Алексеев В.А. Световой режим леса. Л.: Наука, 1975. 227 с.
Биопродукционный процесс в лесных экосистемах Севера / Под ред. Бобковой К.С., Галенко Э.П. СПб.: Наука, 2001. 278 с.
Бобкова К.С. Биологическая продуктивность хвойных лесов европейского Северо-Востока. Л.: Наука, 1987. 156 с.
Бобкова К.С., Урнышев А.П., Урнышев В.А. Вертикальное распределение фитомассы в еловых лесах европейского Северо-Востока // Лесоведение. 2000. № 3. С. 49–54.
Вертикально-фракционное распределение фитомассы в лесах / Под. ред. Вомперского С.Э., А.И. Уткина. М.: Наука, 1986. 260 с.
Гульбе Я.И., Ермолова Л.С., Рождественский С.Г., Уткин А.И., Цельникер Ю.Л. Вертикальное распределение поверхности листьев и световой режим в лиственных молодняках южной тайги // Лесоведение. 1983. № 2. С. 21–29.
Данилин И.М., Фаворская М.Н. Трехмерное моделирование лесных ландшафтных сцен на основе данных дистанционного зондирования // География и природные ресурсы. 2013. № 2. С. 151–159.
Казимиров Н.И., Волков А.Д., Зябченко С.С., Иванчиков А.А., Морозова Р.М. Обмен веществ и энергии в сосновых лесах Европейского Севера. Л.: Наука, 1977. 304 с.
Кутявин И.Н., Бобкова К.С. Биологическая продуктивность сосновых фитоценозов Северного Приуралья (Республика Коми) // Лесоведение. 2017. № 1. С. 3–16.
Кутявин И.Н., Торлопова Н.В., Осипов А.Ф., Кузьмина Е.С., Бобкова К.С. Биологическая продуктивность коренного среднетаежного сосняка бруснично-лишайникового (средняя Печора) // Растительные ресурсы. 2016. Вып. 4. С. 484–500.
Ладанова Н.В., Тужилкина В.В. Структурная организация и фотосинтетическая активность хвои ели сибирской. Сыктывкар: изд-во Коми НЦ УрО РАН, 1992. 100 с.
Лесной фонд России. М.: ВНИИЦлесресурс, 1999. 649 с.
Лукина Н.В., Никонов В.В. Биогеохимические циклы в лесах Севера в условиях аэротехногенного загрязнения. Апатиты: Изд-во Кольского НЦ РАН, 1996. Ч. 1. 213 с; Ч. 2. 192 с.
Молчанов А.А. Продуктивность органической массы в лесах различных зон. М.: Наука, 1971. 257 с.
Пулы и потоки углерода в наземных экосистемах России / Под ред. Заварзина Г.А. М.: Наука, 2007. 315 с.
Смирнов В.В. Органическая масса в некоторых фитоценозах европейской части СССР. М.: Наука, 1971. 362 с.
Усольцев В.А. Вертикально-фракционная структура фитомассы деревьев. Исследование закономерностей. Екатеринбург: Уральский государственный лесотехнический университет, 2013. 603 с.
Уткин А.И. Биологическая продуктивность лесов // Лесоведение и лесоводство. М.: Изд-во ВИНИТИ, 1975. Т. 1. С. 9–190.
Уткин А.И., Дылис Н.В. Изучение вертикального распределения фитомассы в лесных биогеоценозах // Бюл. МОИП. Отд. биол. 1986. Т. 69. Вып. 6. С. 79–91.
Феклистов П.А., Клевцов Д.Н., Кунников Ф.А., Хабарова Е.П., Амомова И.Б. Динамика продуктивности сосновых древостоев разного происхождения // Лесной журн. 2015. № 4. С. 55–60с.
Cermak J., Riguzzi F., Ceulemans R. Scaling up from the individual tree to the stand level in Scots pine. I. Needle distribution, overall crown and root geometry // Annals of Forest Science. 1998. 55. P. 63–88.
Korpela I., Tuomola T., Välimäki E. Mapping forest plots: an efficient method combining photogrammetry and field triangulation // Silva Fennica. 2007. V. 41. № 3. P. 457–469.
Repola J. Models for vertical wood density of Scots pine, Norway spruce and birch stems, and their application to determine average wood density // Silva Fennica. 2006. V. 40. № 4. P. 673–685.
Shanin V., Komarov A., Mäkipää R. Tree species composition affects productivity and carbon dynamics of different site types in boreal forests // European J. Forest Research. 2014. V. 133. P. 273–286.
Stenberg P., Kuuluvainen T., Kellomaki S., Grace J.C., Jokela E.J., Gholz H.L. Crown structure, light interception and productivity of pine trees and stands // Ecological Bulletins. 1994. (Copenhagen). V. 43. P. 63–88.
Stinson G., Kurz W.A., Smyth C.E., Neilson E.T., Dymond C.C., Metsaranta J.M., Boisvenue C., Rampley G.J., Li Q., Wite T.M., Blain D. An inventory-based analysis of Canada’s managed forest carbon dynamics, 1990 to 2008 // Global Change Biology. 2011. V. 17. P. 2227–2244.
Vauhkonen J. Mehtätalo L. Matching remotely sensed and field-measured tree size distributions // Canadian J. Forestry Research. 2014. V. 45. P. 353–363.
Дополнительные материалы отсутствуют.