

Лесоведение, 2020, № 6, стр. 493-502
Экотипическая обусловленность соотношения желто- и краснопыльниковой форм в южносибирских популяциях сосны обыкновенной
А. В. Пименов a, *, А. Е. Коновалова a, М. Е. Коновалова a, Г. Б. Кофман a, Т. С. Седельникова a, С. П. Ефремов a
a Институт леса им. В.Н. Сукачева СО РАН – обособленное подразделение ФИЦ КНЦ СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
* E-mail: pimenov@ksc.krasn.ru
Поступила в редакцию 06.02.2019
После доработки 05.09.2019
Принята к публикации 06.06.2020
Аннотация
Впервые проанализированы связи соотношения числа деревьев желто- и краснопыльниковой форм сосны обыкновенной (Pinus sylvestris L.) и экотопических условий с использованием взаимной энтропии подсистем и частной информации. Установлено, что структура популяций сосны обыкновенной имеет статистически значимую связь с характеристиками экотопа. Долевое участие деревьев краснопыльниковой формы увеличивается в пессимальных для вида условиях произрастания. Исследования проведены в двух географо-климатических зонах юга Сибири.
Внутривидовое разнообразие сосны обыкновенной (Pinus sylvestris L.) определяется экологически обусловленным внутрипопуляционным соотношением интразональных морфотипов (форм) и уникальных онтогенетических модификаций (Правдин, 1964; Мамаев, Махнев, 1996; Пименов, Седельникова, 2012; и др.).
При этом одним из наиболее четких признаков внутривидовой формовой дифференциации популяций сосны является окраска микростробилов – мужских генеративных структур, диагностируемая на уровне желтопыльниковой (f. sulfuranthera Kozubow) и краснопыльниковой (f. erythranthera Sanio) форм (Козубов, 1962; Правдин, 1964). Поскольку процент участия этих форм P. sylvestris в различных частях ареала, а нередко и в смежных контрастных экотопах неодинаков, предполагается, что данное соотношение отражает физиолого-фенологическую поливариантность вида (Некрасова, 1959, 1960; Козубов, 1962; Черепнин, 1980; Дудник и др., 2006; Седельникова и др., 2007; Пименов и др. 2011, 2014; Новикова, 2012; Тарханов, Бирюков, 2013).
Предыдущими исследованиями выявлялось строение популяций по соотношению деревьев желто- и краснопыльниковой форм (Козубов, 1962; Правдин, 1964; Черепнин, 1980; Особенности формирования …, 1984; и др.) и в лучшем случае оценивалась достоверность различий этих структур в контрастных условиях произрастания. Целью нашего исследования являлась конкретизация статистических связей желто- и краснопыльниковой форм P. sylvestris с условиями произрастания.
ОБЪЕКТЫ И МЕТОДИКА
Исследования охватывают разнообразные местопроизрастания сосны обыкновенной на юге Сибири в двух различных, репрезентативных по своим природно-климатическим условиям для региона в целом, районах: Назаровско-Минусинской межгорной впадине (семигумидный сектор) и на Западносибирской равнине в пределах междуречья Оби и Томи (гумидный сектор).
В семигумидном климатическом секторе Алтае-Саянской горной области ксероморфные сосняки, анализируемые в настоящей работе, занимают типичные для юга Сибири боровые и петрофитные экотопы Назаровско-Минусинской межгорной впадины (53.0°–54.5° с.ш., 89.5°–92.0° в.д.; 400–650 м над ур. моря) (Растительный покров …, 1976).
В гумидном климатическом секторе в южнотаежной подзоне Западно-Сибирской равнины (56.0°–57.0° с.ш., 82.0°–89.5° в.д.) объекты исследования включают древостои в болотных и суходольных экотопах (Седельникова, 2008; Пименов, 2015).
Проанализированные данные по 58 пробным площадям представлены в табл. 1.
Таблица 1.
Встречаемость деревьев желто- и краснопыльниковой форм в различных экотопах
| Пробная площадь | Группа типов леса | Трофотоп | Гигротоп | Класс бонитета | Долевое участие деревьев различных форм, %* |
|---|---|---|---|---|---|
| Назаровско-Минусинская межгорная впадина | |||||
| 1, 2 | Лишайниковая остепненная | А | 0 | V–Vб | $\frac{{44 \pm 6.0}}{{56 \pm 6.0}}$ |
| 3, 4 | Олиготрофно-разнотравная | В | 1 | V | $\frac{{27 \pm 5.5}}{{73 \pm 5.5}}$ |
| 5, 6 | Ритидиевая | В | 2 | III | $\frac{{22 \pm 2.1}}{{78 \pm 2.1}}$ |
| 7–18 | Остепненно-разнотравная | А–С | 1–2 | III–IV | $\frac{{2 \pm 0.1}}{{98 \pm 0.1}}$ |
| 19, 20 | Разнотравная | В–С | 3 | II | $\frac{{1 \pm 1.0}}{{99 \pm 1.0}}$ |
| 21, 22 | Мезофильно-разнотравная | В–С | 2–3 | II–IV | $\frac{{14 \pm 3.0}}{{86 \pm 3.0}}$ |
| 23–25 | Вейниково-разнотравная | В–С | 1–2 | II–IV | $\frac{{18 \pm 0.7}}{{82 \pm 0.7}}$ |
| 26–29 | Травяно-болотная | В–С | 4–5 | IV–V | $\frac{{25 \pm 2.7}}{{75 \pm 2.7}}$ |
| Западно-Сибирская равнина | |||||
| 30–32 | Брусничная | С | 3 | Iа–II | $\frac{{3 \pm 0.7}}{{97 \pm 0.7}}$ |
| 33 | Кустарничково-осоковая | В | 3 | Vб | $\frac{6}{{94}}$ |
| 34–37 | Лишайниково-зеленомошная | В–С | 2 | III | $\frac{{9 \pm 1.8}}{{91 \pm 1.8}}$ |
| 38–43 | Крапивно-вейниковая | D | 4 | III–V | $\frac{{20 \pm 2.1}}{{80 \pm 2.1}}$ |
| 44–53 | Сфагново-кустарничковая | А–В | 4–5 | Vа–Vб | $\frac{{26 \pm 2.2}}{{74 \pm 2.2}}$ |
| 54–58 | Кустарничково-сфагновая | А | 5 | Vб | $\frac{{46 \pm 1.8}}{{54 \pm 1.8}}$ |
Таксация древостоев проводилась методом сплошного перечета с определением форм сосны по окраске микростробилов. Для характеристики экотопических условий были выбраны относительно стабильные во времени комплексные показатели: класс бонитета древостоя, тип условий местопроизрастания, группа типов леса.
Класс бонитета древостоя использован в работе в качестве интегрального показателя качества условий местопроизрастания, детерминированного комплексом климатических и почвенных условий произрастания (Орлов, 1927). Он определяется по бонитетной шкале М.М. Орлова, созданной на основании высот древостоев в возрасте 100 лет.
Специфика лесорастительных условий оценивалась по методике Алексеева–Погребняка (Погребняк, 1968), принятой в лесоведении и практике лесного хозяйства для характеристики участков по совокупности действующих на растительность почвенно-гидрологических факторов (эдафотопы). В соответствии с ней определены трофотопы (от A до D) и гигротопы (от 0 до 5).
В качестве комплексной характеристики биогеоценоза рассмотрены группы типов леса. Они, в соответствии с эколого-фитоценотической схемой В.Н. Сукачева (1972), объединяют лесные растительные сообщества, близкие по климатическим и эдафическим условиям, производительности, составу всех ярусов растительности и характеру функционирования.
Анализ связи между структурой популяции сосны и выбранными признаками условий проводился теоретико-информационным методом. В отличие от простой оценки разнообразия с помощью энтропии Шеннона, использовалось разложение энтропии системы, позволяющее численно оценить связь подсистем на основе взаимного ограничения их разнообразия (Legendre P., Legendre L., 1998; Orloci et al., 2002; Stone, 2015; Коновалова и др., 2015; и др.). Такой подход хорошо зарекомендовал себя в естественных науках (Yockey, 1992; Pan, 1995, 2005; Hampe et al., 2003; и др.), так как имеет ряд преимуществ: не зависит от распределения данных, применим к номинальным шкалам, нечувствителен к перестановкам переменных и нулевым значениям, численно выражает силу связи, дает возможность анализа структуры связи.
Соотношение деревьев желто- и краснопыльниковой форм (подсистема X) и различные признаки условий произрастания – классы бонитета, гигротопы, трофотопы, группы типов леса (подсистема Y) – рассматривались как части единой системы. Каждой пробной площади соответствовало определенное число деревьев желто- и краснопыльниковой форм в пересчете на 100 деревьев и фиксированная характеристика условий, которые группировались в таблицу сопряженности, включающую данные по всем пробным площадям определенной территории. Для такой таблицы вычислялись энтропия подсистем, общая взаимная энтропия системы и полная взаимная информация (Кульбак, 1967; Cover, Thomas, 1991; Stone, 2015). Проверка достоверности связи подсистем проводилась путем сравнения общей взаимной информации с минимальной статистически значимой общей взаимной информацией, рассчитываемой для каждой матрицы с заданным уровнем статистической значимости (Krippendorff, 1986; Pan, 1995).
Сравнение величин связи между системами выполнялось с использованием коэффициента нормированной информации RX|Y, имеющего такой же смысл, как и коэффициент детерминации в регрессионном анализе (Елисеева, 1982; Коновалова и др., 2015). Коэффициент RX|Y определяет часть разнообразия подсистемы X, объясняемую подсистемой Y. Значения RX|Y = 0 и = 1 относятся к независимым и эквивалентным подсистемам, соответственно.
Индивидуальная взаимосвязь доли деревьев желто- и краснопыльниковой форм с заданным значением какого-либо признака условий (состояний xi и yj) оценивалась по величине частной информации ${{I}_{{{{y}_{j}} \leftrightarrow {{x}_{i}}}}}$ о каждом состоянии одной подсистемы (yj), получаемой при известных состояниях другой (xi):
где ${{P}_{{ij}}}$ – вероятность состояния (xi, yj) объединенной системы; rj – вероятность состояния yj подсистемы Y; pi – вероятность состояния xi подсистемы X.Положительное значение частной информации ${{I}_{{{{y}_{j}} \leftrightarrow {{x}_{i}}}}}$ > 0 показывает степень соответствия (связь) состояний xi и yj. В случае, если ${{I}_{{{{y}_{j}} \leftrightarrow {{x}_{i}}}}}$ < 0, можно говорить о нехарактерности сочетания двух состояний, их “уникальности”. Нулевое значение частная информация ${{I}_{{{{x}_{i}} \leftrightarrow {{y}_{j}}}}}$ принимает, когда заданные состояния подсистем xi и yj не связаны (Кофман и др., 2016).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
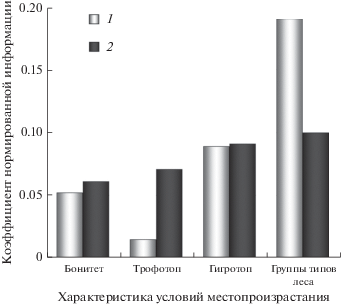
Во всех анализируемых популяциях P. sylvestris в целом преобладает желтопыльниковая форма (табл. 1). Среднее долевое участие деревьев краснопыльниковой формы составляет 22.9 ± 2.7% на Западно-Сибирской равнине и 13.6 ± 2.5% – в Назаровско-Минусинской межгорной впадине. Связь между числом деревьев с различной окраской мужских генеративных структур и рассматриваемыми признаками условий местопроизрастания является статистически достоверной на общепринятом уровне значимости (p = 0.05). Однако влияние различных признаков на структуру популяций существенно различается (рис. 1). В двух рассмотренных районах исследования наименьшую связь с числом деревьев желто- и краснопыльниковой форм в популяциях сосны имеют трофность почв и бонитет древостоя. Наибольшие значения коэффициента нормированной информации соответствуют условиям увлажнения почв (гигротопам) и группам типов леса.
Рис. 1.
Влияние признаков условий местопроизрастания RX|Y на структуру популяций сосны обыкновенной по числу деревьев желто- и краснопыльниковой форм. 1 – Назаровско-Минусинская межгорная впадина; 2 – Западно-Сибирская равнина.
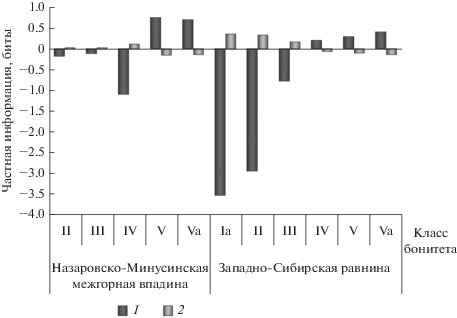
Несмотря на низкие значения RX|Y, для бонитета примечательна структура этой взаимосвязи (рис. 2). Положительная связь числа деревьев краснопыльниковой формы имеется только при низких классах бонитета. Для Назаровско-Минусинской межгорной впадины это V и Vа классы с частной информацией ${{I}_{{{{x}_{i}} \leftrightarrow {{y}_{j}}}}}$ = 0.74 бита и ${{I}_{{{{x}_{i}} \leftrightarrow {{y}_{j}}}}}$ = = 0.69 бита соответственно. На Западно-Сибирской равнине эта величина составляет в насаждениях с IV классом бонитета 0.2 бита, V классом – 0.29 бита, Vа классом – 0.41 бита. Таким образом, деревья краснопыльниковой формы произрастают в насаждениях с любым классом бонитета, но вероятность произрастания дерева краснопыльниковой формы снижается по мере улучшения качества условий местопроизрастания. Эта тенденция более детально прослеживается при анализе связи структуры популяций сосны с эдафическими условиями.
Рис. 2.
Структура связи соотношения числа деревьев красно- и желтопыльниковой форм в популяциях сосны обыкновенной и классов бонитета древостоя. На рис. 2–5: 1 – P. sylvestris f. erythranthera, 2 – P. sylvestris f. sulfuranthera.
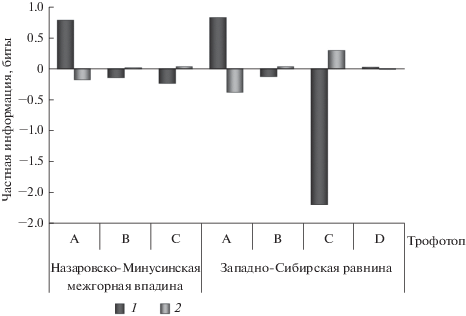
Слабое влияние почвенного плодородия на структуру популяций сосны по соотношению желто- и краснопыльниковой форм (рис. 1), вероятнее всего, связано со значительной толерантностью сосны к данному фактору. В структуре этой связи (рис. 3) значительная положительная сопряженность числа деревьев краснопыльниковой формы имеется только с крайне бедными почвенно-грунтовыми условиями, относящимися к трофотопу А по классификации Алексеева–Погребняка: песчаными или торфяными на олиготрофных болотах. Отрицательная связь наблюдается у деревьев краснопыльниковой формы с относительно богатыми (трофотоп С) и относительно бедными (трофотоп В) эдафическими условиями, соответствующими оптимальным условиям формирования сосняков (на дерновых и серых лесных почвах). Небольшая положительная связь числа деревьев краснопыльниковой формы имеется с богатыми почвами (трофотоп D), обследованными только на объектах Западно-Сибирской равнины на эвтрофных болотах.
Рис. 3.
Структура взаимосвязи соотношения числа деревьев f. erythranthera и f. sulfuranthera в популяциях P. sylvestris и условий почвенного плодородия. Обозначения см. рис. 2.
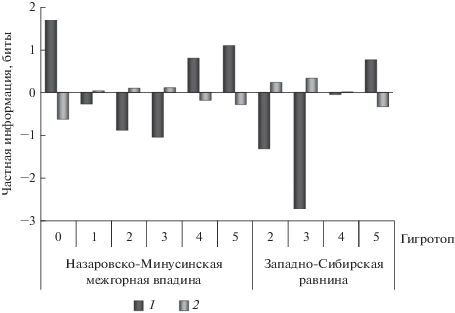
Признаком, более связанным со структурой популяций, является влажность почв (рис. 1). Известно, что сосна обыкновенная произрастает в очень широком диапазоне увлажнения почв – от торфяно-болотных до песчано-сухостепных (Правдин, 1964). Однако оптимум произрастания выражается сравнительно небольшими значениями влажности почвы (Поликарпов и др., 1986). Структура связи соотношения числа деревьев желто- и краснопыльниковой форм с условиями почвенного увлажнения, в целом, хорошо согласуется с характером ее связи с показателем качества условий местопроизрастания: при крайнем недостатке (гигротоп 0 – очень сухие) и избытке влаги (гигротопы 5 – мокрые и 4 – сырые) повышается вероятность участия в популяциях краснопыльниковой формы (рис. 4). И, напротив, в оптимальных для сосны условиях (гигротопы 3 – влажные, 2 – свежие и 1 – сухие) связь носит отрицательный характер. Иными словами, деревья краснопыльниковой формы встречаются в оптимальных условиях увлажнения почв реже, чем при случайном распределении.
Рис. 4.
Структура взаимосвязи соотношения числа деревьев f. erythranthera и f. sulfuranthera в популяциях P. sylvestris и условий почвенного увлажнения. Обозначения см. рис. 2.
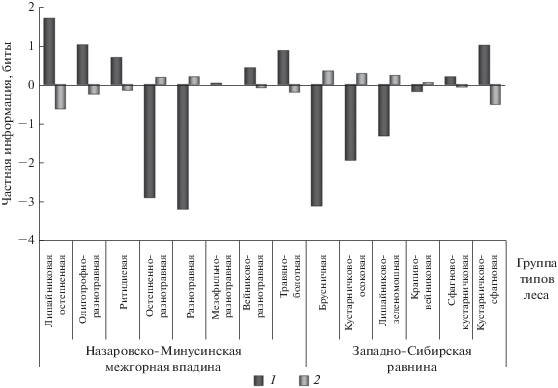
Максимальная сопряженность соотношения желто- и краснопыльниковой форм обнаружена с группами типов леса (рис. 1). Последние являются наиболее детальным из рассмотренных показателем условий местопроизрастания, учитывающим сходство климатических, орографических, эдафических факторов и определяемым на основании индикаторных свойств растительности. Соответственно, наибольшая взаимосвязь соотношения числа деревьев желто- и краснопыльниковой форм в популяциях сосны с этим признаком доказывает зависимость структуры популяции от всего комплекса экологических факторов.
Увеличение числа деревьев краснопыльниковой формы сопряжено с ухудшением условий произрастания. В частности, повышается вероятность присутствия деревьев краснопыльниковой формы в кустарничково-сфагновой и травяно-болотной группах типов леса в условиях избыточного увлажнения и в лишайниковой остепненной, олиготрофно-разнотравной и ритидиевой группах в экстремально сухих и олиготрофных условиях (рис. 5). Напротив, наименьшая вероятность участия в составе древостоев краснопыльниковой формы соответствует группам типов леса, характеризующим условия, приближающиеся к оптимальным для произрастания сосны: разнотравной, остепненно-разнотравной и брусничной, а также кустарничково-осоковой и лишайниково-зеленомошной. Встречаемость деревьев краснопыльниковой формы, близкая к случайной, наблюдается в мезофильно-разнотравной, сфагново-кустарничковой, вейниково-разнотравной и крапивно-вейниковой группах типов леса на довольно богатых влагообеспеченных почвах.
Рис. 5.
Структура взаимосвязи соотношения числа деревьев f. erythranthera и f. sulfuranthera в популяциях P. sylvestris и групп типов леса. Обозначения см. рис. 2.
Таким образом, различия в структуре популяций по соотношению желто- и краснопыльниковой и форм обусловлены широким спектром экологических условий. Для Назаровско-Минусинской межгорной впадины и Западносибирской равнины в пределах междуречья Томи и Оби соотношение в популяциях сосны обыкновенной числа деревьев желто- и краснопыльниковой форм имеет статистически значимую связь с условиями местопроизрастания на уровне экотопа. Это совпадает с результатами исследований, проведенными в других регионах – бассейне Северная Двина (Дудник и др., 2006; Тарханов, Бирюков, 2013), Карелии, на Кольском полуострове (Козубов, 1962) и Центральном Кавказе (Темботова и др., 2017). Численность деревьев краснопыльниковой формы в популяциях сосны обыкновенной возрастает в неблагоприятных эдафических условиях. Это означает, что деревья краснопыльниковой формы проявляют повышенную выживаемость в экстремальных условиях. Этот результат хорошо согласуется с тем, что физиологическая природа разнокачественности желто- и краснопыльниковой форм во многом определяется высоким содержанием антоцианов в мужских шишках краснопыльниковой формы, адаптивно значимым для растений в экстремальных условиях их произрастания (Некрасова, 1959; Козубов, 1962; Наквасина, Бедрицкая, 2002). Это также подтверждается результатами исследований сезонной динамики биохимических показателей у желто- и краснопыльниковой форм сосны, обнаружившими относительное увеличение концентрации стрессовых белков в хвое деревьев краснопыльниковой формы (Аганина, Тарханов, 2016), и подтверждает предположение об определяющей роли лимитирующих факторов в организации формовой структуры популяций сосны (Новикова, 2012). Для обследованной нами на фоне общих климатических и зональных особенностей территории юга Сибири таким фактором выступает влажность почв.
ЗАКЛЮЧЕНИЕ
Полученные результаты конкретизируют связь желто- и краснопыльниковой форм сосны обыкновенной с признаками условий местопроизрастания, раскрывая функциональную роль поливариантности вида в разнообразных природно-климатических условиях юга Сибири.
Анализ структуры связей в популяциях сосны обыкновенной позволяет рассматривать желто- и краснопыльниковую формы в качестве генетически обусловленных, “маркирующих” соответственно пессимальные и оптимальные условия произрастания. Данные формы сосны обыкновенной являются генетически обособленными компонентами внутривидового разнообразия, обеспечивающими устойчивость вида в широком диапазоне факторов среды.
На юге Сибири экотопические условия оказывают существенное влияние на соотношение числа деревьев желто- и краснопыльниковой форм в популяциях сосны обыкновенной. Максимальное влияние на структуру популяций оказывают признаки, в большей степени характеризующие лимитирующий фактор почвенного увлажнения – гигротоп и группа типов леса.
В контрастных биоклиматических секторах юга Сибири – гумидном и семигумидном – участие деревьев желтопыльниковой формы максимально в популяциях сосны обыкновенной из оптимальных условий произрастания. Напротив, краснопыльниковая форма увеличивает свое присутствие в сосновых древостоях пессимальных почвенных условий – экстремально сухих или избыточно увлажненных.
Список литературы
Аганина Ю.Е., Тарханов С.Н. Изменчивость биохимических показателей и адаптация краснопыльниковой и желтопыльниковой форм сосны (Pinus sylvestris L.) в условиях избыточного увлажнения // Известия Самарского НЦ РАН. 2016. Т. 18. № 2. С. 10–14.
Дудник С.В., Тарханов С.Н., Щекалёв Р.В. Фенотипическая изменчивость сосны на территории Северо-Двинского бассейна // Актуальные проблемы лесного комплекса. Брянск: БГИТА, 2006. Вып. 15. С. 16–18.
Елисеева И.И. Статистические методы измерения связей. Л.: Изд-во Ленингр. университета, 1982. 136 с.
Козубов Г.М. О краснопыльниковой форме сосны обыкновенной // Ботанический журн. 1962. Т. 47. № 2. С. 276–280.
Коновалова М.Е., Кофман Г.Б., Коновалова А.Е. Сопряженность признаков рельефа и типов леса в горных условиях // География и природные ресурсы. 2015. № 2. С. 177–182.
Кофман Г.Б., Коновалова М.Е., Коновалова А.Е. Интегральная и парциальная сопряженность растительности и элементов рельефа // Интенсификация лесного хозяйства России: проблемы и инновационные пути решения: Матер. Всеросс. науч.-практич. конф. с междунар. участием. Красноярск, 19–23 сентября 2016 г. Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 2016. С. 117–119.
Кульбак С. Теория информации и статистика. М.: Наука, 1967. 408 с.
Мамаев С.А., Махнев А.К. Проблемы биологического разнообразия и его поддержания в лесных экосистемах // Лесоведение. 1996. № 5. С. 3–10.
Наквасина Е.Н., Бедрицкая Т.В. Изменчивость потомства сосны обыкновенной различного географического происхождения по цвету пыльников // Там же. 2002. № 3. С. 18–22.
Некрасова Т.П. О значении желтой и розовой окраски мужских шишек у видов Pinus // Ботанический журнал. 1959. Т. 44. № 7. С. 975–978.
Некрасова Т.П. Плодоношение сосны в Западной Сибири. Новосибирск: Изд-во СО АН СССР, 1960. 132 с.
Новикова Т.Н. Сибирские климатипы сосны в географических культурах Западного Забайкалья // Хвойные бореальной зоны. 2012. Т. 30. № 1–2. С. 140–144.
Орлов М.М. Лесоустройство. Л.: Изд-во журн. “Лесное хозяйство, лесопромышленность и топливо”, 1927. 1120 с.
Особенности формирования популяции сосны обыкновенной / Отв. ред. Некрасов В.И. М.: Наука, 1984. 128 с.
Пименов А.В. Биоразнообразие сосны обыкновенной (Pinus sylvestris L.) в контрастных экотопах юга Сибири: Дис. … д-ра биол. наук (03.02.01). Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 2015. 346 с.
Пименов А.В., Седельникова Т.С. Качественная оценка формового разнообразия сосны обыкновенной в лесоболотных комплексах Западной Сибири // Хвойные бореальной зоны. 2012. Т. 30. № 1–2. С. 157–161.
Пименов А.В., Седельникова Т.С., Ефремов С.П. Морфология и качество пыльцы желто- и краснопыльниковой форм Pinus sylvestris в болотных и суходольных условиях произрастания (Томская область) // Ботанический журн. 2011. Т. 96. № 3. С. 367–376.
Пименов А.В., Седельникова Т.С., Ефремов С.П. Морфология и качество пыльцы сосны обыкновенной в контрастных экотопах Хакасии // Лесоведение. 2014. № 1. С. 57–64.
Погребняк П.С. Общее лесоводство. М.: Колос, 1968. 440 с.
Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. Климат и горные леса Южной Сибири. Новосибирск: Наука, 1986. 226 с.
Правдин Л.Ф. Сосна обыкновенная (изменчивость, внутривидовая систематика и селекция). М.: Наука, 1964. 190 с.
Растительный покров Хакасии / Под ред. Куминовой А.В. Новосибирск: Наука, 1976. 421 с.
Седельникова Т.С. Дифференциация болотных и суходольных популяций видов семейства Pinaceae Lindl. (репродуктивные и кариотипические особенности): Дис. … д-ра биол. наук (03.00.05). Красноярск: Институт леса им. В.Н. Сукачева СО РАН, 2008. 323 с.
Седельникова Т.С., Пименов А.В, Ефремов С.П., Муратова Е.Н. Особенности генеративной сферы сосны обыкновенной болотных и суходольных популяций // Лесоведение. 2007. № 4. С. 44–50.
Сукачев В.Н. Основы лесной типологии и биогеоценологии. Л.: Наука, 1972. Т. 1. 420 с.
Тарханов С.Н., Бирюков С.Ю. Формовое разнообразие Pinus sylvestris (Pinaceae) в бассейне Северной Двины // Растительные ресурсы. 2013. Т. 49. Вып. 4. С. 481–490.
Темботова Ф.А., Моллаева М.З., Пшегусов Р.Х. Изменчивость пыльцы желто- и краснопыльниковой форм сосны обыкновенной (Pinus sylvestris L.) на территории национального парка “Приэльбрусье” (Центральный Кавказ) // Известия Дагестанского государственного педагогического университета. Естественные и точные науки. 2017. Т. 11. № 4. С. 55–61.
Черепнин В.Л. Изменчивость семян сосны обыкновенной. Новосибирск: Наука, 1980. 183 с.
Cover T.M., Thomas J.A. Elements of information theory. N.Y.: John Wiley&Sons, 1991. 542 p.
Hampe J., Schreiber S., Krawczak M. Entropy-based SNP selection for genetic association studies // Human Genetics. 2003. № 114. P. 36–43.
Krippendorff K. Information theory: structural models for qualitative data. Newbury Park: SAGE Publications, 1986. 96 p.
Legendre P., Legendre L. Numerical ecology. Amsterdam: Elsevier, 1998. № 20. 853 p.
Orloci L., Anand M., Pillar V.D. Biodiversity analysis: issues, concepts, techniques // Community Ecology. 2002. V. 3. № 2. P. 217–236.
Pan G.C. Related information measures for the associations of earth-science variables // Mathematical geology. V. 27. № 5. 1995. P. 609–632.
Stone J.V. Information theory: A tutorial introduction. Sheffield: Sebtel Press, 2015. 243 p.
Yockey H.P. Information theory and molecular biology. Cambridge: Cambridge University Press, 1992. 428 p.
Yockey H.P. Information theory, evolution and the origin of life. Cambridge: Cambridge University Press, 2005. 259 p.
Дополнительные материалы отсутствуют.