

Лесоведение, 2021, № 3, стр. 250-260
Изменчивость метрических признаков листа тополя черного и лавролистного в зоне естественной гибридизации
Б. В. Прошкин a, *, В. В. Тараканов b, c, А. В. Климов c, d
a ФКОУ ВО Кузбасский институт ФСИН России
654066 Новокузнецк, пр. Октябрьский, 49, Россия
b Новосибирский государственный аграрный университет
630039 Новосибирск, ул. Добролюбова, 160, Россия
c Западно-Сибирское отделение Института леса им. В.Н. Сукачева СО РАН – филиал
Федерального исследовательского центра “Красноярский научный центр СО РАН”
630082 Новосибирск, ул. Жуковского, 100/1, Россия
d ООО ИнЭкА-консалтинг
654027 Новокузнецк, ул. Лазо, 4, Россия
* E-mail: boris.vladimirovich.93@mail.ru
Поступила в редакцию 21.03.2020
После доработки 21.05.2020
Принята к публикации 03.02.2021
Аннотация
Анализ иерархической структуры изменчивости количественных признаков листа является одним из обязательных этапов оценки генетической гетерогенности природных популяций в масштабе ареала вида. В зонах естественного скрещивания таксонов он позволяет не только оценить вклад различных эколого-генетических факторов в изменчивость количественных признаков этих видов, но и пролить дополнительный свет на особенности процессов гибридизации и интрогрессии. Поэтому цель настоящих исследований заключалась в изучении изменчивости метрических признаков листа и ее иерархической структуры в популяциях Populus nigra L., P. laurifolia Ledeb. и их естественного гибрида P. jrtyschensis Ch. Y. Yang в бассейне р. Томь. По результатам изучения морфометрических признаков и их индексов гибридный таксон P. jrtyschensis, как и следовало ожидать, занимает промежуточное положение между родительскими видами P. nigra и P. laurifolia, но отличается от них определенными особенностями структуры изменчивости. Все это в совокупности свидетельствует о преобладании в популяциях P. jrtyschensis гибридов F1, что, возможно, сочетается с действием стабилизирующего отбора. Несмотря на преобладание в составе P. jrtyschensis гибридов F1, для него, в сравнении с родительскими видами, характерен более высокий уровень общей изменчивости, возникающий за счет большого вклада индивидуального (внутрипопуляционного) компонента дисперсии. Это свидетельствует о генетической неоднородности гибридного таксона, которая может создаваться появлением рекомбинантных генотипов от скрещивания гибридов 1-го поколения друг с другом и с родительскими видами. Наряду с этим в очагах гибридизации у P. nigra наблюдается достоверное увеличение размеров листа в сравнении с его популяциями вне очагов. Наиболее простым объяснением этого феномена является интрогрессивная аккумуляция этим видом генов “крупнолистности” от донора P. laurifolia.
Изучение интрогрессивной гибридизации имеет важнейшее значение для понимания механизмов эволюции и адаптации многих видов древесных растений (Коропачинский, Милютин, 2006). В роде Populus L. особенно интересны межсекционные гибриды секций Aigeros Lunell и Tacamahaca Mill. В частности, в Азии в бассейне Черного Иртыша (КНР) и на юге Западной Сибири P. nigra L. и P. laurifolia Ledeb. в зонах перекрывания их ареалов скрещиваются с образованием гибридного таксона P. jrtyschensis Ch. Y. Yang (Yang et al., 1992; Климов, Прошкин, 2016).
Китайскими исследователями с использованием маркеров ДНК показано, что свыше 90% гибридов, образующих P. jrtyschensis, представлено поколением F1 (Jiang et al., 2016; Zeng et al., 2016). При этом в качестве матери чаще выступает P. laurifolia. Гибриды же преимущественно скрещиваются с P. nigra, вследствие чего интрогрессивный поток генов направлен в сторону этого вида. Лучшие вегетативно размножаемые экземпляры P. jrtyschensis используются в Китае для озеленения и в программах селекции (Yang et al., 1992). Данные по фенотипической изменчивости в публикациях китайских коллег практически отсутствуют.
Для сибирской части ареала, напротив, отсутствуют данные по изменчивости P. jrtyschensis на молекулярно-генетическом уровне, но имеются данные об изменчивости на уровне фенотипов. При этом первые сведения о естественной гибридизации P. nigra и P. laurifolia были получены В.Я. Поляковым еще в 1950 г. (Бакулин, 2007). Но обстоятельное изучение изменчивости морфологических и анатомических признаков P. nigra, P. laurifolia и P. jrtyschensis в зоне гибридизации начато лишь недавно (Климов, Прошкин, 2016, 2018, 2019).
В ходе наших исследований по изменчивости качественных морфологических и анатомических признаков родительских видов и гибридного таксона в бассейне р. Томь уточнены критерии идентификации гибридных особей, а также выявлена асимметрия процессов гибридизации и интрогрессии; в принципиальных чертах она схожа с той, которая описана китайскими исследователями по молекулярно-генетическим маркерам (Климов, Прошкин, 2016, 2018, 2019; Klimov, Proshkin, 2019). При этом не было осуществлено детальное изучение изменчивости морфометрических признаков листа, которое представляет несомненный интерес по следующим причинам.
Во-первых, количественные признаки имеют селективную природу, что проявляется, в частности, в способности организмов к адаптивным модификациям; последние контролируются нормой реакции генотипов, разнообразие которых отражается в феномене взаимодействия “генотип–среда” (Glotov, 1992). Без изучения их изменчивости, ограничиваясь изучением только генов и ферментов/белков, невозможно оценить роль отбора в адаптации и эволюции видов (Левонтин, 1978). Значительный интерес при этом представляют морфометрические показатели фотосинтезирующих органов - листьев.
Во-вторых, большинство количественных признаков контролируется полигенно (Guet et al., 2015). Поэтому при вовлечении в исследования большого числа фенотипических признаков косвенно оценивается значительная часть генома вида, связанная с адаптивной сферой. При этом пока имеются лишь весьма ограниченные данные о том, какие именно гены отвечают за те или иные фенотипические признаки, что делает невозможным изучение адаптивной части генома только молекулярно-генетическими методами.
В-третьих, сложность генетической обусловленности количественных признаков обязана не только большому числу контролирующих их генов. С одной стороны, спектры генов, контролирующих сложные количественные признаки (например, семенную продуктивность), могут меняться под влиянием лимитирующих факторов, что лежит в основе взаимодействий “генотип–среда” (Драгавцев, Драгавцева, 2011). С другой стороны, вследствие недостаточной изученности биохимической основы формирования фенотипических признаков и закона “эмержентности” (Коросов, 2012) большинство количественных признаков лишь косвенно зависят от свойств конкретных белков и ферментов, напрямую контролируемых генами.
В силу рассмотренных особенностей количественных признаков их изучение предложено в качестве одного из обязательных этапов оценки генетической гетерогенности природных популяций в масштабе ареала вида (Глотов, 1983; Семериков, 1986). Анализ изменчивости количественных признаков на различных уровнях – внутрииндивидуальном, внутрипопуляционном и межпопуляционном – позволяет получать представление об относительном вкладе наследственных и ненаследственных факторов в общую изменчивость, особенностях пространственной популяционной структуры вида, а также уровне гомеостаза развития. Анализ иерархической структуры изменчивости количественных признаков листа оказался очень эффективным при изучении видов дубов (Глотов, 1983; Семериков, 1986). С нашей точки зрения, исходя из многочисленных данных по гибридологическому анализу растений, он может оказаться информативным и при оценке эффектов интрогрессивной гибридизации.
Известно, что законы Менделя проявляются в изменчивости количественных признаков следующим образом: 1) по средним значениям количественных признаков гибриды 1-го поколения F1 в сравнении с родителями РР обычно занимают промежуточное положение и характеризуются одновершинными распределениями признаков; 2) расщепления в F2 приводят к существенному увеличению изменчивости в сравнении с F1 и РР; 3) потомство от возвратных скрещиваний Fb сдвигается по средним значениям признаков в сторону соответствующего родителя (Рокицкий, 1978). Из этого можно заключить, что сравнительное изучение структуры изменчивости у родительских видов тополя и их дочернего гибридного таксона может не только оценить вклад различных эколого-генетических факторов в изменчивость количественных признаков этих таксонов, но и пролить дополнительный свет на особенности процессов гибридизации и интрогрессии. Учитывая адаптивную значимость признаков фотосинтезирующих органов и существенные различия в размерах и форме листа у P. nigra и P. laurifolia, сравнительный анализ структуры изменчивости признаков листа у данных видов и у их естественного гибрида P. jrtyschensis представляет очевидный интерес.
В связи с этим основная цель исследований заключалась в изучении изменчивости метрических признаков листа и ее иерархической структуры в популяциях P. nigra, P. laurifolia и их естественного гибрида P. jrtyschensis в бассейне р. Томь.
ОБЪЕКТЫ И МЕТОДИКА
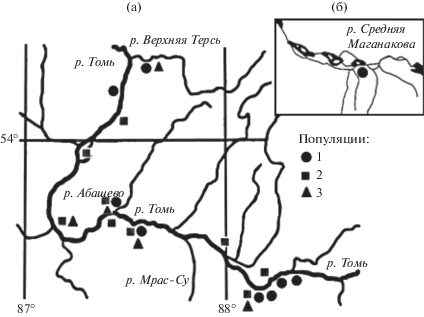
Исследования были выполнены на 684 особях из 23 популяций (пробных площадей внутри ценопопуляций) в бассейне р. Томь, включая девять популяций P. nigra, девять – P. laurifolia и пять – P. jrtyschensis (рис. 1). Название популяции каждого таксона определялось ее географическим положением. Географические координаты популяций в пойме Томи и ее притоков приведены в работе (Климов, Прошкин, 2018).
Рис. 1.
Расположение изученных популяций: а – участок верхнего и среднего течения р. Томь, б – участок бассейна р. Средняя Маганакова. Популяции: 1 – P. laurifolia, 2 – P. nigra, 3 – P. jrtyschensis.
Для исключения сезонной, возрастной и онтогенетической изменчивости сбор гербарного материала проводился со второй половины июля по сентябрь в 2015–2018 гг., и только с укороченных побегов южной стороны средней части кроны репродуктивно зрелых деревьев в возрасте 40–80 лет. С каждой особи отбирали по 15 полностью развитых, неповрежденных листьев.
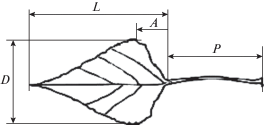
На каждом листе были измерены 4 признака (мм): длина листовой пластинки L, максимальная ширина листовой пластинки D, длина черешка Р, расстояние между самой широкой частью листовой пластинки и ее основанием А (рис. 2) (Бакулин, 2007). Также оценивали отношения трех последних признаков к длине листовой пластинки (P : L, D : L, A : L).
Рис. 2.
Метрические признаки листа: D – максимальная ширина листовой пластинки, L – длина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, Р – длина черешка.
Измерения признаков осуществляли с помощью программы “Axio Vision 4.8.2” (ZEISS, 2018). Статистическую обработку данных проводили с помощью программ Excel и IBM SPSS Statistics 23.0 software (IBM …, 2018). Суммарный объем выборки составил 10 260 листьев, на которых выполнено 41040 измерений.
Для каждого из признаков вычисляли арифметические средние и их ошибки ($\bar {x}$ ± m), коэффициенты вариации (C.V., %) и парной корреляции, проводили дисперсионный анализ с разложением общей дисперсии признаков на компоненты (Гласс, Стэнли, 1976; Глотов, 1983).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Распределения признаков “внутри дерева”, рассчитанные по нормированным отклонениям их значений от средних, близки к нормальным. Все признаки положительно коррелированны между собой в диапазоне коэффициентов корреляции от 0.257 до 0.703 (n = 450, P < 0.001). Наиболее интегрированным во взаимосвязи с другими признаками является длина листа, наименее – длина черешка.
По средневзвешенным на популяцию оценкам самые крупные листья длиной около 112 и шириной 62 мм имеет тополь лавролистный (табл. 1). Наименьшие размеры длина около 76, ширина 57 мм наблюдаются у P. nigra. В популяциях P. jrtyschensis значения признаков, как и следовало ожидать, промежуточные, с небольшими отклонениями в сторону того или другого родительского вида. То же относится к остальным признакам, за исключением длины черешка (Р), которая у P. jrtyschensis по осредненным оценкам близка к таковой у P. nigra (табл. 1). При этом, если исключить из выборки тополя лавролистного популяцию “Ерунаково” с необычно большой длиной черешка и самыми крупными листьями, то все таксоны по этому признаку окажутся близки между собой. Отметим также, что внутри видов многие популяции достоверно отличаются друг от друга. При этом у P. nigra прослеживается межгрупповая изменчивость: ценопопуляции в очагах гибридизации имеют более крупные листья, чем вне их. Например, по длине листа (L) популяционные средние этих групп варьируют в пределах около 78–85 и 63–79 мм соответственно (табл. 1). К этой проблеме мы вернемся при обсуждении результатов дисперсионного анализа.
Таблица 1.
Средние арифметические и ошибки средних исходных признаков
| Популяция | L | Р | D | A | n |
|---|---|---|---|---|---|
| P. nigra | |||||
| Славино | 73.70 ± 0.57 | 44.10 ± 0.37 | 55.40 ± 0.35 | 21.70 ± 0.18 | 450 |
| Казанково | 71.10 ± 0.54 | 33.20 ± 0.40 | 51.30 ± 0.39 | 22.00 ± 0.16 | 450 |
| Кийзак | 66.90 ± 0.43 | 39.70 ± 0.34 | 53.40 ± 0.30 | 18.90 ± 0.15 | 450 |
| Чистенький | 63.70 ± 0.50 | 36.40 ± 0.36 | 50.30 ± 0.34 | 19.80 ± 0.15 | 450 |
| Швейник | 79.80 ± 0.45 | 40.90 ± 0.33 | 57.60 ± 0.40 | 21.70 ± 0.14 | 450 |
| ГРЭС* | 84.80 ± 0.56 | 46.00 ± 0.37 | 64.90 ± 0.38 | 22.60 ± 0.17 | 450 |
| Карлык* | 78.30 ± 0.46 | 44.70 ± 0.30 | 54.70 ± 0.36 | 20.10 ± 0.16 | 450 |
| Майзас* | 82.70 ± 0.47 | 43.40 ± 0.31 | 63.30 ± 0.30 | 23.60 ± 0.14 | 450 |
| Новокузнецк* | 85.10 ± 0.59 | 49.40 ± 0.33 | 59.40 ± 0.37 | 22.10 ± 0.17 | 450 |
| НСР0.01 | 2.12 | 1.42 | 1.44 | 0.65 | |
| P. laurifolia | |||||
| Бельсинская | 90.20 ± 0.88 | 31.40 ± 0.45 | 46.00 ± 0.49 | 33.30 ± 0.45 | 450 |
| Вороний | 106.80 ± 0.98 | 37.10 ± 0.56 | 54.70 ± 0.61 | 43.30 ± 0.38 | 450 |
| Ерунаково | 138.10 ± 0.65 | 63.30 ± 0.53 | 76.60 ± 0.54 | 47.80 ± 0.29 | 450 |
| Средняя Маганакова | 108.80 ± 0.66 | 41.60 ± 0.45 | 62.40 ± 0.48 | 39.00 ± 0.30 | 450 |
| Студеный плес | 107.60 ± 1.37 | 45.60 ± 0.82 | 60.50 ± 0.90 | 37.20 ± 0.56 | 450 |
| ГРЭС* | 123.70 ± 0.66 | 46.00 ± 0.46 | 71.80 ± 0.44 | 44.50 ± 0.28 | 450 |
| Карлык* | 111.30 ± 0.57 | 42.00 ± 0.39 | 63.30 ± 0.39 | 41.30 ± 0.30 | 450 |
| Майзас* | 105.10 ± 0.79 | 41.80 ± 0.60 | 56.70 ± 0.49 | 40.00 ± 0.32 | 450 |
| Верхняя Терсь* | 117.60 ± 0.66 | 47.00 ± 0.41 | 67.70 ± 0.41 | 41.50 ± 0.27 | 450 |
| НСР0.01 | 4.34 | 2.63 | 2.80 | 1.85 | |
| P. jrtyschensis | |||||
| ГРЭС | 94.20 ± 0.59 | 47.50 ± 0.52 | 63.20 ± 0.48 | 30.50 ± 0.21 | 465 |
| Карлык | 88.70 ± 0.41 | 41.10 ± 0.30 | 54.90 ± 0.27 | 30.10 ± 0.15 | 615 |
| Майзас | 92.60 ± 0.52 | 40.20 ± 0.32 | 62.40 ± 0.36 | 31.80 ± 0.19 | 546 |
| Новокузнецк | 86.90 ± 1.11 | 41.10 ± 0.71 | 59.20 ± 0.83 | 27.10 ± 0.37 | 345 |
| Верхняя Терсь | 80.40 ± 0.63 | 38.60 ± 0.53 | 53.80 ± 0.50 | 24.90 ± 0.27 | 150 |
| НСР0.01 | 3.31 | 2.27 | 2.51 | 1.18 | |
| Среднее на популяцию | |||||
| P. nigra | 76.16 ± 0.206 | 41.96 ± 0.137 | 56.69 ± 0.140 | 21.38 ± 0.057 | 4050 |
| P. laurifolia | 112.15 ± 0.344 | 43.98 ± 0.222 | 62.16 ± 0.229 | 40.89 ± 0.137 | 4050 |
| P. jrtyschensis | 90.11 ± 0.298 | 42.04 ± 0.212 | 59.38 ± 0.225 | 29.82 ± 0.110 | 2121 |
* Популяции P. nigra и P. laurifolia в очагах гибридизации. L – длина листовой пластинки, Р – длина черешка, D – максимальная ширина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, n – объем выборки, НСР – наименее существенная разность при Р < 0.01.
Аналогичные данные о промежуточных значениях показателей у гибридного таксона в сравнении с родительскими видами получены и по индексам признаков (табл. 2). В этом случае также сохраняется существенная межпопуляционная изменчивость. В частности, крупнолистная популяция “Ерунаково” P. laurifolia, отличающаяся необычно длинными черешками, сохраняет лидирующее положение и по относительной длине этого признака – P : L.
Таблица 2.
Средние арифметические и ошибки средних индексов признаков
| Популяция | P : L | D : L | A : L | n |
|---|---|---|---|---|
| P. nigra | ||||
| Славино | 0.606 ± 0.0052 | 0.758 ± 0.0038 | 0.295 ± 0.0016 | 450 |
| Казанково | 0.468 ± 0.0044 | 0.729 ± 0.0047 | 0.311 ± 0.0012 | 450 |
| Кийзак | 0.597 ± 0.0046 | 0.802 ± 0.0030 | 0.284 ± 0.0018 | 450 |
| Чистенький | 0.575 ± 0.0047 | 0.797 ± 0.0041 | 0.312 ± 0.0013 | 450 |
| Швейник | 0.515 ± 0.0038 | 0.724 ± 0.0037 | 0.274 ± 0.0017 | 450 |
| ГРЭС* | 0.546 ± 0.0039 | 0.772 ± 0.0043 | 0.268 ± 0.0015 | 450 |
| Карлык* | 0.574 ± 0.0036 | 0.701 ± 0.0032 | 0.257 ± 0.0017 | 450 |
| Майзас* | 0.528 ± 0.0036 | 0.771 ± 0.0034 | 0.288 ± 0.0016 | 450 |
| Новокузнецк* | 0.587 ± 0.0040 | 0.704 ± 0.0036 | 0.262 ± 0.0017 | 450 |
| P. laurifolia | ||||
| Бельсинская | 0.346 ± 0.0034 | 0.511 ± 0.0028 | 0.370 ± 0.0031 | 450 |
| Вороний | 0.343 ± 0.0038 | 0.512 ± 0.0033 | 0.410 ± 0.0028 | 450 |
| Ерунаково | 0.458 ± 0.0029 | 0.553 ± 0.0022 | 0.347 ± 0.0019 | 450 |
| Средняя Маганакова | 0.382 ± 0.0036 | 0.575 ± 0.0033 | 0.358 ± 0.0016 | 450 |
| Студеный плес | 0.407 ± 0.0035 | 0.559 ± 0.0030 | 0.345 ± 0.0026 | 450 |
| ГРЭС* | 0.372 ± 0.0031 | 0.583 ± 0.0030 | 0.361 ± 0.0017 | 450 |
| Карлык* | 0.380 ± 0.0034 | 0.571 ± 0.0031 | 0.370 ± 0.0017 | 450 |
| Майзас* | 0.394 ± 0.0045 | 0.540 ± 0.0026 | 0.384 ± 0.0025 | 450 |
| Верхняя Терсь* | 0.400 ± 0.0028 | 0.578 ± 0.0025 | 0.354 ± 0.0016 | 450 |
| P. jrtyschensis | ||||
| ГРЭС | 0.503 ± 0.0044 | 0.673 ± 0.0038 | 0.326 ± 0.0018 | 465 |
| Карлык | 0.463 ± 0.0027 | 0.622 ± 0.0025 | 0.340 ± 0.0013 | 615 |
| Майзас | 0.435 ± 0.0028 | 0.677 ± 0.0027 | 0.345 ± 0.0014 | 546 |
| Новокузнецк | 0.473 ± 0.0048 | 0.687 ± 0.0055 | 0.312 ± 0.0020 | 345 |
| Верхняя Терсь | 0.481 ± 0.0054 | 0.672 ± 0.0052 | 0.310 ± 0.0028 | 150 |
| Среднее на популяцию | ||||
| P. nigra | 0.555 ± 0.0016 | 0.752 ± 0.0014 | 0.285 ± 0.0009 | 4050 |
| P. laurifolia | 0.386 ± 0.0013 | 0.553 ± 0.0011 | 0.367 ± 0.0008 | 4050 |
| P. jrtyschensis | 0.467 ± 0.0017 | 0.662 ± 0.0017 | 0.332 ± 0.0008 | 2121 |
* Популяции P. nigra и P. laurifolia в очагах гибридизации. P : L – отношение длины черешка к длине листовой пластинки, D : L – отношение максимальной ширины листовой пластинки к длине листовой пластинки, A : L – отношение расстояния между самой широкой частью листовой пластинки и ее основанием к длине листовой пластинки, n – объем выборки.
Если исходить из предположения о подавляющем преобладании в составе популяций P. jrtyschensis гибридов первого поколения, то их изменчивость по всем признакам должна быть приблизительно такой же или даже ниже, чем у родительских видов. Для исключения эффекта шкалы при проверке этого предположения использовали внутрипопуляционные коэффициенты вариации признаков. Их осредненные значения находятся в пределах 11–26% (табл. 3). При этом предположение о меньшей изменчивости гибридного таксона не подтверждается. По признакам P и D P. jrtyschensis занимает промежуточное положение между родительскими таксонами, по остальным он приближается к P. nigra.
Таблица 3.
Внутрипопуляционные коэффициенты вариации признаков (%)
| Вид тополя | L | Р | D | A | P : L | D : L | A : L |
|---|---|---|---|---|---|---|---|
| P. nigra | 14.20 | 17.60 | 13.30 | 15.70 | 16.20 | 10.70 | 11.80 |
| P. laurifolia | 16.10 | 26.20 | 19.00 | 19.10 | 19.30 | 11.20 | 13.00 |
| P. jrtyschensis | 14.90 | 22.50 | 16.50 | 15.80 | 16.60 | 11.30 | 10.40 |
L – длина листовой пластинки, Р – длина черешка, D – максимальная ширина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, P : L – отношение длины черешка к длине листовой пластинки, D : L – отношение максимальной ширины листовой пластинки к длине листовой пластинки, A : L – отношение расстояния между самой широкой частью листовой пластинки и ее основанием к длине листовой пластинки.
Одним из информативных аспектов изучения структуры изменчивости является ее разложение на компоненты методами дисперсионного анализа (Глотов, 1983; Efimov et al., 2019). При использованной методике сбора образцов возможно вычленение из общей дисперсии каждого признака эндогенного (внутрикронного), индивидуального (внутрипопуляционного) и межпопуляционного компонентов. Эндогенная изменчивость листьев внутри дерева имеет паратипическую (экологическую) природу, отражая влияние микроусловий внутри кроны. Индивидуальная изменчивость имеет смешанную эколого-генетическую природу, в суммарном виде отражая экологическую мозаичность среды обитания и генотипические различия между деревьями. Межпопуляционная изменчивость суммирует эффекты средовых (биотопных) и генетических межпопуляционных различий.
Вначале рассмотрим соотношение внутрикронного и индивидуального компонентов изменчивости различных признаков. Они получены по данным однофакторного дисперсионного анализа признаков. Несмотря на варьирование оцениваемого показателя по популяциям, усредненные данные выявляют совершенно определенную и неожиданную картину: по всем признакам наибольшими значениями долей влияния деревьев характеризуется P. jrtyschensis (табл. 4) (значение статистики критерия Фридмана $\chi _{R}^{2}$ = = 10.571; k = 3; n = 7; P < 0.01). Этот результат, так же, как и анализ коэффициентов вариации, но еще более наглядно подтверждает предположение о генетической неоднородности P. jrtyschensis, в составе которого, очевидно, присутствуют не только гибриды 1-го поколения.
Таблица 4.
Коэффициенты внутриклассовой корреляции (доли влияния деревьев в общей дисперсии, %), по данным однофакторного дисперсионного анализа
| Популяция | L | Р | D | A | P : L | D : L | A : L |
|---|---|---|---|---|---|---|---|
| P. nigra | |||||||
| ГРЭС | 30.60 | 28.50 | 26.80 | 46.90 | 29.70 | 45.00 | 9.10 |
| Казанково | 63.50 | 67.50 | 69.50 | 48.90 | 58.60 | 38.70 | 33.30 |
| Карлык | 60.90 | 48.60 | 65.90 | 59.70 | 39.20 | 54.30 | 21.20 |
| Кийзак | 37.80 | 24.30 | 50.60 | 14.90 | 18.90 | 23.80 | 33.40 |
| Майзас | 45.70 | 13.80 | 40.70 | 35.00 | 9.30 | 25.00 | 30.20 |
| Новокузнецк | 64.70 | 51.10 | 61.40 | 59.50 | 48.80 | 57.20 | 44.30 |
| Славино | 62.20 | 56.60 | 46.10 | 48.70 | 34.00 | 42.10 | 17.80 |
| Чистенький | 73.30 | 62.10 | 51.50 | 66.60 | 56.40 | 49.20 | 25.70 |
| Швейник | 49.90 | 55.30 | 61.10 | 48.10 | 58.90 | 37.80 | 47.30 |
| P. laurifolia | |||||||
| Бельсинская | 62.80 | 30.80 | 39.90 | 55.00 | 27.30 | 12.50 | 37.10 |
| Вороний | 63.80 | 60.50 | 46.50 | 40.90 | 55.50 | 36.40 | 36.70 |
| ГРЭС | 33.90 | 19.10 | 30.80 | 35.90 | 15.10 | 31.80 | 20.70 |
| Ерунаково | 42.80 | 26.20 | 35.70 | 42.50 | 29.20 | 14.60 | 33.20 |
| Карлык | 36.40 | 39.20 | 56.10 | 55.60 | 54.80 | 51.20 | 48.20 |
| Майзас | 69.30 | 58.80 | 65.10 | 64.70 | 55.20 | 42.90 | 58.60 |
| Средняя Маганакова | 27.30 | 37.90 | 44.10 | 39.90 | 49.30 | 37.20 | 25.00 |
| Студеный плес | 87.30 | 74.70 | 83.70 | 73.90 | 49.80 | 44.50 | 17.70 |
| Верхняя Терсь | 62.00 | 53.80 | 66.40 | 47.70 | 51.20 | 43.10 | 29.30 |
| P. jrtyschensis | |||||||
| ГРЭС | 56.80 | 50.90 | 67.00 | 38.60 | 49.80 | 66.30 | 50.30 |
| Карлык | 60.90 | 59.00 | 62.60 | 64.10 | 56.70 | 48.60 | 45.10 |
| Майзас | 67.20 | 58.80 | 68.20 | 56.80 | 61.50 | 50.30 | 47.20 |
| Новокузнецк | 92.50 | 81.80 | 91.80 | 87.60 | 61.90 | 82.00 | 37.20 |
| Верхняя Терсь | 51.10 | 36.00 | 67.20 | 49.40 | 32.30 | 58.80 | 39.30 |
| Среднее на популяцию | |||||||
| P. nigra | 54.30 | 45.30 | 52.60 | 47.60 | 39.30 | 41.50 | 29.10 |
| P. laurifolia | 54.00 | 44.50 | 52.00 | 50.70 | 43.00 | 34.90 | 34.10 |
| P. jrtyschensis | 65.70 | 57.30 | 71.30 | 59.30 | 52.40 | 61.20 | 43.80 |
Примечание. Влияние фактора “деревья” статистически значимо при P < 0.001 во всех случаях. L – длина листовой пластинки, Р – длина черешка, D – максимальная ширина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, P : L – отношение длины черешка к длине листовой пластинки, D : L – отношение максимальной ширины листовой пластинки к длине листовой пластинки, A : L – отношение расстояния между самой широкой частью листовой пластинки и ее основанием к длине листовой пластинки.
На следующем этапе осуществляли наиболее полный анализ структуры изменчивости, используя для этой цели двухфакторную иерархическую схему (модель компонент дисперсии (Гласс, Стэнли, 1976)) (табл. 5). Из общей изменчивости признаков вычленяли не только внутрикронный и индивидуальный, но также и межпопуляционный компоненты. В среднем соотношение межпопуляционного, внутрипопуляционного (индивидуального) и внутрикронного (эндогенного) компонентов дисперсии длины и ширины листа, а также черешка листовой пластинки у родительских видов P. nigra и P. laurifolia составляет соответственно около 30–35, 30–40 и 25–40% (табл. 6). Структура изменчивости индексных показателей и признака “А” менее стабильна, и отличается понижением вклада межпопуляционной дисперсии до уровня около 15–20%.
Таблица 5.
Характеристика дисперсионного комплекса, используемого для оценки долей влияния факторов в двухфакторной иерархической схеме, на примере P. nigra
| Источник изменчивости | Число наблюдений | N = IJn | Структура среднего квадрата |
|---|---|---|---|
| Популяции (I) Деревья (J) Побеги внутри кроны (К) |
9 30 15 |
4050 | $\sigma _{K}^{2}$ + 15×$\sigma _{J}^{2}$ + 15×30×$\sigma _{I}^{2}$ $\sigma _{K}^{2}$ + 15×$\sigma _{J}^{2}$ $\sigma _{K}^{2}$ |
Таблица 6.
Доли влияния факторов (%), вычисленные по данным двухфакторного иерархического дисперсионного анализа
| Признак | Вид тополя | Источник изменчивости | ||
|---|---|---|---|---|
| популяции | деревья | побеги внутри крон | ||
| L | P. nigra | 32.70 | 37.30 | 30.00 |
| P. laurifolia | 34.20 | 41.90 | 24.00 | |
| P. jrtyschensis | 7.00 | 68.30 | 24.70 | |
| Р | P. nigra | 29.60 | 30.90 | 39.50 |
| P. laurifolia | 35.10 | 33.70 | 31.20 | |
| P. jrtyschensis | 8.90 | 57.30 | 33.80 | |
| D | P. nigra | 29.80 | 37.50 | 32.70 |
| P. laurifolia | 37.20 | 36.00 | 26.70 | |
| P. × jrtyschensis | 13.30 | 66.30 | 20.40 | |
| A | P. nigra | 15.10 | 40.90 | 44.00 |
| P. laurifolia | 21.50 | 43.40 | 35.10 | |
| P. jrtyschensis | 17.80 | 53.50 | 28.70 | |
| Р : L | P. nigra | 18.50 | 34.90 | 46.70 |
| P. laurifolia | 16.00 | 38.50 | 45.50 | |
| P. jrtyschensis | 9.40 | 49.80 | 40.70 | |
| D : L | P. nigra | 17.10 | 28.40 | 54.50 |
| P. laurifolia | 14.80 | 30.90 | 54.30 | |
| P. jrtyschensis | 11.00 | 57.30 | 31.60 | |
| A : L | P. nigra | 13.80 | 16.30 | 69.90 |
| P. laurifolia | 14.20 | 29.80 | 55.90 | |
| P. jrtyschensis | 14.10 | 38.70 | 47.30 | |
Примечание. Влияние факторов достоверно при Р < 0.001. L – длина листовой пластинки, Р – длина черешка, D – максимальная ширина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, P : L – отношение длины черешка к длине листовой пластинки, D : L – отношение максимальной ширины листовой пластинки к длине листовой пластинки, A : L – отношение расстояния между самой широкой частью листовой пластинки и ее основанием к длине листовой пластинки.
Особенности иерархической структуры изменчивости в интегральном виде отражают специфику пространственной популяционной структуры вида, которая складывается в ее длительном историческом взаимодействии со средой обитания (Глотов, 1983; Семериков, 1986). Существенное отклонение структуры изменчивости у гибридных таксонов от таковой у родительских видов может маркировать те или иные особенности их генофондов, обусловленные спецификой процессов гибридизации.
Поскольку соотношение первых двух компонентов рассмотрено выше, в данном случае при сравнении гибридов с родительскими видами наиболее интересен вклад в общую дисперсию межпопуляционных различий. Он в большинстве случаев оказался наименьшим для P. jrtyschensis, варьируя в пределах 7–17% (табл. 6) (значение статистики критерия Фридмана $\chi _{R}^{2}$ = 7.143; k = 3; n = 7; P < 0.05). Очевидно, этот результат может быть обусловлен преобладанием в составе всех популяций данного вида относительно “единообразных” гибридов первого поколения и/или действием стабилизирующего отбора.
Тем не менее, несмотря на близость оценок структуры изменчивости родительских видов P. nigra и P. laurifolia, природа межпопуляционной изменчивости их может быть различна. В этой связи напомним, что популяции P. nigra в зонах гибридизации характеризуются более крупными листьями в сравнении с таковыми вне зон (табл. 1). Вклад этого фактора в общую дисперсию размерных признаков оценивали в 3-факторной иерархической схеме дисперсионного анализа. Для облегчения восприятия результатов приводим соотношение компонентов дисперсий лишь по факторам “очаги гибридизации” и “популяции”, на которые у родительских видов приходится около 15–35% общей дисперсии. Если сумму этих двух компонентов принять за 100%, то доля влияния очагов гибридизации в межпопуляционной изменчивости трех признаков (L, P и D) и одного индекса (A : L) может достигать 56–71% (табл. 7). При этом влияние зон гибридизации на межпопуляционную дифференциацию у P. laurifolia совершенно не прослеживается.
Таблица 7.
Доли влияния факторов “Зоны гибридизации” и “Популяции” в сумме дисперсий по этим факторам, вычисленные по данным иерархического дисперсионного анализа признаков в популяциях P. nigra и P. laurifolia
| Признак | Вид тополя | Доли влияния, % | |
|---|---|---|---|
| зоны гибридизации | популяции | ||
| L | P. nigra | 70.90** | 29.10*** |
| P. laurifolia | 0.00 | 100.00*** | |
| Р | P. nigra | 63.50** | 36.50*** |
| P. laurifolia | 0.00 | 100.00*** | |
| D | P. nigra | 61.30* | 38.70*** |
| P. laurifolia | 0.00 | 100.00*** | |
| A | P. nigra | 17.20 | 82.80*** |
| P. laurifolia | 0.00 | 100.00*** | |
| Р:L | P. nigra | 0.00 | 100.00*** |
| P. laurifolia | 0.00 | 100.00*** | |
| D:L | P. nigra | 0.10 | 99.90*** |
| P. laurifolia | 22.60 | 77.40*** | |
| A:L | P. nigra | 56.10* | 43.90*** |
| P. laurifolia | 0.00 | 100.00*** | |
* P < 0.05,** P < 0.01, *** P < 0.001, L – длина листовой пластинки, Р – длина черешка, D – максимальная ширина листовой пластинки, А – расстояние между самой широкой частью листовой пластинки и ее основанием, P : L – отношение длины черешка к длине листовой пластинки, D : L – отношение максимальной ширины листовой пластинки к длине листовой пластинки, A : L – отношение расстояния между самой широкой частью листовой пластинки и ее основанием к длине листовой пластинки.
Суммируем и обсудим основные результаты исследований.
Первый из них заключается в том, что по средним значениям большинства морфометрических признаков гибридный таксон P. jrtyschensis занимает промежуточное положение по отношению к родительским видам P. nigra и P. laurifolia. Этот вполне ожидаемый результат дополняет впервые обнаруженный нами феномен пониженного уровня межпопуляционной изменчивости P. jrtyschensis. В совокупности это свидетельствует о преобладании в его популяциях гибридов F1, что, возможно, сочетается с действием стабилизирующего отбора. Это заключение полностью согласуется с ранее полученными данными по китайской части зоны гибридизации и с нашими результатами (Климов, Прошкин, 2016, 2018, 2019; Jiang et al., 2016). При этом у гибридов не отмечается выраженного доминирования количественных признаков крупного яйцевидного листа, свойственных P. laurifolia, несмотря на наличие дискобластов, характерных только для P. laurifolia.
Второй интересный результат заключается в том, что, несмотря на преобладание в составе P. jrtyschensis гибридов F1, для него, в сравнении с родительскими видами, характерен более высокий уровень общей изменчивости, возникающий за счет большого вклада индивидуального (внутрипопуляционного) компонента дисперсии. Это свидетельствует о генетической неоднородности гибридного таксона, которая может создаваться появлением рекомбинантных генотипов от скрещивания гибридов 1-го поколения друг с другом и с родительскими видами. В этой связи напомним, что внутрикронный компонент отражает преимущественно вклад в изменчивость листьев микрофлуктуаций экологических условий внутри кроны дерева и, возможно, некоторую онтогенетическую разнокачественность листьев в связи с особенностями их размещения на побеге. Индивидуальный компонент изменчивости между деревьями имеет смешанную эколого-генетическую природу и отражает как влияние микроусловий произрастания отдельных деревьев, так и их генотипические различия.
В целом рассмотренный выше результат также хорошо согласуется с ранее полученными данными по изменчивости качественных анатомо-морфологических признаков в исследуемых популяциях (Климов, Прошкин, 2018; Klimov, Proshkin, 2019).
Третья и наиболее важная особенность полученных данных состоит в том, что в очагах гибридизации, в которых на ограниченной площади произрастают все исследуемые виды, именно у P. nigra наблюдается достоверное увеличение размеров листа в сравнении с его популяциями вне очагов гибридизации. Наиболее простым объяснением этого феномена является интрогрессивная аккумуляция этим видом генов “крупнолистности” от донора P. laurifolia. Этот результат, с одной стороны, дает дополнительное обоснование выводу об асимметрии интрогрессии, который мы сделали на основании анализа изменчивости признаков петиолярной анатомии. С другой, он усиливает аргументы в пользу адаптивной природы этого явления, поскольку оно проявляется не только в увеличении потенциала вегетативного размножения осокоря (Klimov, Proshkin, 2019), но и в увеличении биомассы его фотосинтезирующих органов.
ЗАКЛЮЧЕНИЕ
Анализ изменчивости морфометрических признаков листа у P. nigra, P. laurifolia и гибридного таксона P. jrtyschensis в значительной мере подтверждает выводы, которые были получены нами и другими исследователями при изучении изменчивости этих видов по качественным признакам. Прежде всего, это относится к вполне ожидаемому факту промежуточного положения средних значений большинства морфометрических признаков гибридного таксона P. jrtyschensis по отношению к родительским видам P. nigra и P. laurifolia. Однако в очагах гибридизации, в которых на ограниченной площади произрастают все исследуемые виды, у P. nigra было выявлено достоверное увеличение размеров листа в сравнении с его популяциями вне очагов. В этой связи выдвинуто предположение об интрогрессивной аккумуляции этим видом генов “крупнолистности” от донора P. laurifolia.
Новые результаты получены при изучении структуры изменчивости исследуемых признаков. Они сводятся к тому, что в сравнении с родительскими видами гибридный таксон P. jrtyschensis характеризуется сниженным уровнем межпопуляционной дифференциации, но высокой индивидуальной изменчивостью. Вероятными причинами этого являются: 1) преобладание в популяциях P. jrtyschensis гибридов F1 и действие стабилизирующего отбора, которые сглаживают межпопуляционные различия; 2) выщепление рекомбинантных генотипов, возникающее от скрещивания гибридов 1-го поколения друг с другом и с родительскими видами, которое увеличивает внутрипопуляционную дисперсию.
Для проверки выдвинутых гипотез необходимы дополнительные исследования с применением ДНК–маркеров, а также проведение искусственных скрещиваний родительских видов и их гибридов и анализом расщеплений в контролируемых условиях.
Список литературы
Бакулин В.Т. Тополь черный в Западной Сибири. Новосибирск: Гео, 2007. 121 с.
Гласс Д., Стэнли Д. Статистические методы в педагогике и психологии. М.: Прогресс, 1976. 496 с.
Глотов Н.В. Оценка генетической гетерогенности природных популяций: количественные признаки // Экология. 1983. № 1. С. 3−10.
Драгавцев В.А., Драгавцева Е.В. Механизмы сдвигов доминирования количественных признаков яровой пшеницы в разных географических точках // Генетика. 2011. Т. 47. № 5. С. 691–696.
Климов А.В., Прошкин Б.В. Морфологическая идентификация естественных гибридов P. nigra × P. laurifolia в пойме р. Томи // Сибирский лесной журн. 2016. № 5. С. 55−62. https://doi.org/10.15372/SJFS20160506
Климов А.В., Прошкин Б.В. Фенотипическое разнообразие качественных признаков Populus nigra L., P. laurifolia Ledeb. и P. × jrtyschensis Ch. Y. Yang. в зоне естественной гибридизации // Вавиловский журн. генетики и селекции. 2018. Т. 22. № 4. С. 468−475. https://doi.org/10.18699/VJ18.384
Климов А.В., Прошкин Б.В. Использование морфо-анатомических признаков для выявления гибридных растений в зоне естественной гибридизации Populus laurifolia и P. nigra в Сибири, Россия // Вестник томского государственного университета. Биология. 2019. № 46. С. 64−81. https://doi.org/10.17223/19988591/46/4
Коропачинский И.Ю., Милютин Л.И. Естественная гибридизация древесных растений. Новосибирск: Гео, 2006. 223 с.
Коросов А.В. Принцип эмерджентности в экологии // Принципы экологии. 2012. № 3. С. 48–66. https://doi.org/10.15393/j1.art.2012.1481
Левонтин Р. Генетические основы эволюции. М.: Мир. 1978. 352 с.
Рокицкий П.Ф. Введение в статистическую генетику: Учеб. пособие. И.: Высш. школа. 1978. 448 с.
Семериков Л.Ф. Популяционная структура древесных растений. М.: Наука, 1986. 140 с.
Efimov V.M., Tarakanov V.V., Naumova N.B., Kovaleva V.Y., Kutsenogiy K.P. Highly inheritable variables-components in the clonal plantation of Scots Pine // Sibirskij Lesnoj Zurnal. 2019. V. 6. P. 82−88. https://doi.org/10.15372/SJFS20190609
Glotov N.V. Ana1ysis of the genotype-environment interaction in natural populations // Acta Zoo1ogy Fennica. 1992. V. 5. P. 45−53.
Guet J., Fabbrini F., Fichot R., Sabatti M., Bastien C., Brignolas F., Thomas S. Genetic variation for leaf morphology, leaf structure and leaf carbon isotope discrimination: European populations of black poplar (Populus nigra L.) // Tree Physiology. 2015. V. 35. № 8. P. 850−863. https://doi.org/10.1093/treephys/tpv056
IBM SPSS Statistics 23. 2018. http://www-01.ibm.com/ support/ docview.wss?uid=swg24038592.
Jiang D., Feng J., Dong M., Wu G., Mao K., Liu J. Genetic origin and composition of a natural hybrid poplar Populus × × jrtyschensis from two distantly related species // Plant Biology. 2016. V. 16(1). P. 88−99. https://doi.org/10.1186/s12870-016-0776-6
Klimov A.V., Proshkin B.V. Identification of Populus nigra, P. laurifolia and P. × jrtyschensis by leaf petiole anatomy // Sibirskij Lesnoj Zurnal. 2019. V. 6. P. 89−99. https://doi.org/10.15372/SJFS20190610
Yang C.Y., Shen K.M., Mao Z.M. Flora Xinjianggensis. Populus L. Urumqi: Technology & Hygiene Publishing House, 1992. T. 1. V. 2. P. 122−158.
ZEISS AxioVision – Microscope software for materials microscopy & industrial applications. Carl Zeiss Microscopy. Jena, Germany: Zeiss Int. TM, 2018. https://www.zeiss.com/ microscopy/int/products/microscope-software/axiovision.html.
Zeng Y.F., Zhang J.G., Duan A.G., Abuduhamiti B. Genetic structure of Populus hybrid zone along the Irtysh River provides insight into plastid-nuclear incompatibility // Scientific Reports. 2016. V. 6. P. 377−389. https://doi.org/10.1038/srep28043
Дополнительные материалы отсутствуют.