

Лесоведение, 2022, № 5, стр. 461-477
Картографирование фитоценотического разнообразия горнотаежных лесов Северо-Восточного Забайкалья
М. В. Бочарников a, *, А. А. Виноградов a
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, д. 1, Россия
* E-mail: maxim-msu-bg@mail.ru
Поступила в редакцию 03.06.2021
После доработки 19.07.2021
Принята к публикации 02.02.2022
- EDN: CECKYF
- DOI: 10.31857/S0024114822040027
Аннотация
Приводится результат картографического анализа пространственной структуры горнотаежного пояса растительности и фитоценотического разнообразия бореальных лесов Северовосточно-Забайкальского оробиома. С использованием оригинальных полевых данных и литературных источников приведена характеристика светлохвойных, темнохвойных и мелколиственных лесов, раскрыто их типологическое разнообразие на уровне преобладающих в растительном покрове классов и групп растительных ассоциаций. На основе сопряженного анализа карт растительности и цифровой модели рельефа с использованием дискриминантного анализа для обширной горной территории составлена актуальная аналитическая карта лесов (по состоянию на 2015 г.), выполненная в мелком масштабе. Она отразила пространственную структуру горнотаежного пояса с выявлением его типологического разнообразия, представленного сообществами лиственничных, сосновых, еловых, пихтово-кедровых и березовых лесов. Характер их пространственной организации определяет региональную специфику фитоценотического разнообразия оробиома, которая находит выражение в дифференциации горнотаежного пояса на высотные подпояса и полосы. Определены ведущие эколого-географические факторы высотно-поясной и внутрипоясной дифференциации разнообразия лесов с учетом связей сообществ с орографической структурой территории. Выявлены различия в оптимумах развития сообществ лесных формаций и комплексе ключевых фитоценотических признаков, которые обуславливают высотную дифференциацию хода восстановительной динамики растительности и характера дестабилизирующих процессов, способствующих ее активизации, а также преобладающей роли выполняемых сообществами экологических функций. Полученные закономерности возможно учитывать при разработке системы мониторинга и охраны лесов малоизученной горной территории, использовании лесных ресурсов в соответствии с природными особенностями сообществ и экотопическим разнообразием.
Оценка типологического разнообразия и современного состояния лесов, выявление закономерностей их пространственно-временной организации являются ключевыми проблемами, решаемыми в рамках фундаментальных биогеографических исследований и востребованными практической сферой в лесоводстве и лесном хозяйстве. При возрастающей роли лесных ресурсов в экономике страны усиливается эксплуатация лесов, понижается уровень биоразнообразия в связи с трансформацией фоновых и уникальных комплексов экосистем, снижается их резистентность к неблагоприятным воздействиям. Это вызывает особую тревогу при наблюдаемых глобальных тенденциях в изменении климата, под воздействием которых прогнозируются существенные изменения в структуре разнообразия растительного покрова (Johnson et al., 2011). Особенно актуальны данные проблемы для зоны Байкало-Амурской магистрали, характеризующейся усилением освоения природных ресурсов и высокими темпами потепления климата, что находит отклик в разнообразии и структуре растительного покрова (Чебакова и др., 2003; Kharuk et al., 2017).
Комплекс негативных последствий на экосистемы ставит под угрозу их устойчивое развитие, воспроизводство лесных ресурсов, что обуславливает необходимость создания единой системы инвентаризации, мониторинга состояния и охраны лесов. Создание и эффективное функционирование данной системы в масштабах обширной территории возможна на природной основе, предполагающей эколого-географический подход к классификации растительных сообществ с учетом региональных особенностей формирования растительного покрова. Они основаны на биогеоценотических принципах разнообразия лесов (Сукачев, 1931) и соподчиненной системе единиц растительного покрова на глобальном, региональном и топологическом уровнях (Сочава, 1979).
Концепция биомной организации биосферы и классификация наземных экосистем (Walter, Breckle, 1991), использованная при картографировании биомного разнообразия (Карта “Биомы …”, 2018), представляет собой надежную основу для выявления региональных закономерностей в структуре биоразнообразия с учетом современных климатических условий и ландшафтной структуры территории. Горные территории занимают особое положение в структуре биомного разнообразия. Выраженный высотный градиент способствует формированию сопряженных в своем разнообразии и пространственной структуре высотно-поясных спектров растительности, рассматриваемых в качестве базового компонента экосистем, которые образуются в рамках одного или нескольких схожих типов высотной поясности (Огуреева, Бочарников, 2017). Горные биомы, или оробиомы, характеризуют региональное единство экосистем, в пределах которого сохраняется определенный уровень разнообразия биоты и ее пространственной (высотно-поясная и внутрипоясная) структуры. Успешное использование оробиомов при анализе флористического и фитоценотического разнообразия различных регионов (Огуреева, 1991; Бочарников, 2018; Bocharnikov et al., 2018) определяет перспективы выявления типологического разнообразия лесов и разработки системы их мониторинга на единой биомной основе.
Горные леса в силу сложной структуры территории, на которой они произрастают, требуют особого подхода к изучению. Он основан, прежде всего, на закономерностях, связанных с универсальным высотным градиентом, который определяет высотно-поясную структуру растительного покрова и отдельных его компонентов (Vetaas, Grytnes, 2002). С изменениями в обеспеченности теплом, влагой, а также с их соотношением связаны переходы между высотно-поясными подразделениями. К ним относятся пояса и подпояса, выделяемые в соответствии с дифференциацией господствующих типов растительности, фитоценотического оптимума фоновых сообществ в пределах типов поясности (Огуреева, 1991). Высотные изменения лесных экосистем, выраженные в типологическом разнообразии сообществ, составе ценофлор, пространственной структуре, лесотаксационных показателях, находящие связь с градиентами в системе экотопических условий, получили интегральное выражение в системе высотно-поясных комплексов типов леса, выделенных в горах Южной Сибири (Поликарпов и др., 1986; Назимова и др., 2005).
Решению задач, связанных с выявлением высотной дифференциации типологического разнообразия горных лесов, эколого-географических факторов их пространственной структуры, динамических явлений в лесном покрове на региональном уровне способствует применение картографического метода. На современном этапе составление инвентаризационных и оценочных карт, рассматриваемых в качестве основы для выявления пространственно-временных закономерностей организации экосистем, проводится с использованием материалов аэро-, фото- и космической съемки (Назимова и др., 2020). В спектр задач, решаемых на основе обработки снимков и картографического анализа составленных тематических продуктов, входят актуальные для науки и практики задачи экосистемного управления лесами и оптимального многоцелевого лесопользования. Их решение возможно с учетом региональной специфики растительного покрова и его свойств, характеризующих их развитие в рамках высотно-поясных спектров на базе спутниковой информации.
Контрастные по комплексу природных условий горные территории обуславливают формирование разнообразия растительности в рамках типологических подразделений разных типов поясности. В настоящем исследовании поставленной целью послужило выявление типологического разнообразия бореальных лесов Северо-Восточного Забайкалья, формирующих горнотаежный пояс и участвующих в сложении подгольцового пояса, с оценкой актуального состояния их пространственной структуры в растительном покрове обширной горной территории. Развитие массивных хребтов и межгорных котловин, значительные перепады абсолютных высот, высокий уровень разнообразия экотопических условий на каждом высотном уровне определяют природные предпосылки для высотной дифференциации лесного покрова и его важнейших характеристик разнообразия и пространственной структуры. В связи с этим в работе поставлены задачи, связанные с инвентаризацией типологического разнообразия, выявлением высотных градиентов в фитоценотическом составе горнотаежного пояса, определением региональной специфики в пространственной структуре разнообразия лесов для Северобайкальского географического варианта Северовосточно-Забайкальского оробиома. Картографический метод послужил основой для пространственного анализа, определив возможности проведения мониторинга на базе высотно-поясной и внутрипоясной структуры растительного покрова.
ОБЪЕКТЫ И МЕТОДИКА
Экосистемное разнообразие Северовосточно-Забайкальского оробиома формируется в континентальной части Евразии в водосборных бассейнах реки Верхняя Ангара и правых притоках реки Лена (Витим, Чара, Большая Чуя, Большой Патом). Исследуемая территория характеризуется выраженным горным рельефом с преобладанием массивных хребтов субширотного простирания, достигающих абсолютных высот 2500–3000 м (Верхнеангарский, Кодар, Калар, Удокан), и обширных межгорных котловин, приуроченных к Байкальской рифтовой зоне (Верхнеангарская, Муйско-Куандинская, Чарская). Географическое положение территории в центральной части обширного материка определяет ограниченное влияние атлантических воздушных масс и крайне незначительное влияние муссонной циркуляции. При господстве резкоконтинентального типа климата выражена его дифференциация под влиянием горного рельефа, которая проявляется в высотном градиенте тепло- и влагообеспеченности, различиях в увлажнении наветренных (северные и западные) и подветренных (южные и восточные) склонов хребтов. Разнообразие растительного покрова связано с формированием трехчленного гольцово-подгольцово-горнотаежного высотного спектра с соответствующей ему высотно-поясной организацией растительного покрова на хребтах Северо-Байкальского, Станового и Патомского нагорий в условиях сложной орографической структуры территории и выраженной дифференциации климатических условий (Зорин, 1971; Пешкова, 1985; Бочарников, 2019). Северобайкальский вариант оробиома формируется в западной части Станового нагорья от северного побережья Байкала до среднего течения реки Витим в районе Муйско-Куандинской котловины. Крупнейшими хребтами являются Верхнеангарский (водораздел бассейнов Байкала и реки Лена), Северо-Муйский, Южно-Муйский, Делюн-Уранский (абсолютные высоты – до 2000–2500 м). К Байкальской рифтовой зоне приурочены обширные межгорные Верхнеангарская, Муйско-Куандинская, Чарская котловины с днищами на высотах 500–600 м.
Бореальные леса Северо-Восточного Забайкалья являются фоновыми для горнотаежного пояса растительности, формирующегося в нижней и средней частях высотных спектров на абсолютных высотах 500–900 м, местами распространяясь выше 1000 м (Bocharnikov et al., 2018). В высотно-поясной структуре растительности в нижней части спектра формируется горнотаежный пояс, на который приходится около половины площади оробиома. В верхней части высотного спектра развиты высокогорные комплексы подгольцового, горнотундрового и гольцового поясов (рис. 1). В соответствии с географо-генетической классификацией растительности (Сочава, 1980) господствующее положение в горнотаежном поясе занимают леса Восточносибирского или Ангаридского комплекса формаций, представленные лиственничной (лиственица Гмелина (Larix gmelinii)) формацией. Сосновые (сосна обыкновенная (Pinus sylvestris)) и темнохвойные (ель сибирская (Picea obovata), сосна сибирская (Pinus sibirica), пихта сибирская (Abies sibirica)) леса Южно-Сибирского комплекса имеют ограниченное распространение, преимущественно в нижней части горнотаежного пояса. Общее представление о фитоценотическом разнообразии, экологии и географии лесов региона дают литературные источники (Пешкова, 1985; Рысин, 2010), а также инвентаризационные (Белов, 1973; Барталев и др., 2011; Владимиров и др., 2014; Софронов, 2015) и оценочные (Лавренко, 1977) карты.
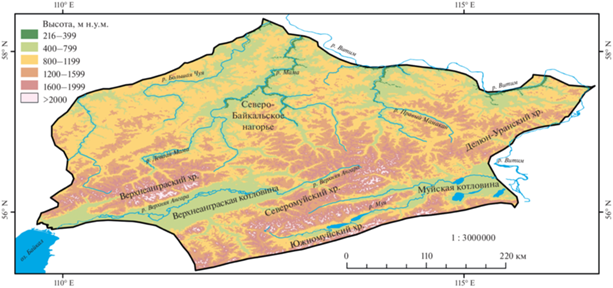
Исследование проведено на основе комплексного анализа оригинальных полевых данных, литературных источников и картографических материалов. Полевые геоботанические работы проведены в ходе экспедиционных исследований на Становом нагорье (Северо-Муйский, Южно-Муйский и Делюн-Уранский хребты; Верхне-Ангарская и Муйско-Куандинская котловины) в 2018 и 2019 гг. (рис. 1). Выполнено 50 полных и кратких геоботанических описаний фоновых для горнотаежного и подгольцового поясов лесных и редколесных сообществ по стандартным методикам (Сукачев, Зонн, 1961). Описания выполнялись по эколого-топологическим профилям для охвата разнообразия экотопических условий, к которым приурочены сообщества на разных уровнях склонов обследованных хребтов. Анализ типологического разнообразия лесов проведен на основе табличной обработки описаний с использованием эколого-фитоценотической классификации растительности (Нешатаев, 1987) и учетом географо-генетических принципов формирования растительности и ее динамического развития (Сочава, 1980). При анализе описаний учитывались литературные данные о типологическом разнообразии светлохвойных и темнохвойных лесов региона (Русяева, 1979; Панарин, 1980; Осипов, 2005). Они послужили основой при эколого-географической оценке лесного покрова и формировании представлений о его разнообразии на региональном уровне. В качестве материалов, характеризующих пространственную структуру растительного покрова, использованы мелкомасштабные карты растительности. Разнообразие лесных сообществ на уровне фоновых формаций и групп формаций определено на основе карты растительности России (Барталев и др., 2011), которая составлена на основе обработки космических снимков низкого пространственного разрешения (MODIS). Актуальное состояние лесного покрова приведено по данным обработки космической съемки за 2015 г. (Барталев и др., 2015). Для оценки разнообразия и структуры в пределах формаций использована карта растительности юга Восточной Сибири (Белов, 1973). Составление карты лесов географического варианта оробиома и проведение анализа их эколого-географической дифференциации выполнено с использованием цифровой модели рельефа ASTER GDEM с пространственным разрешением 30″ (около 1 км2).
Методологической основой исследования послужила концепция экосистемного разнообразия. Обусловленная градиентами климатических показателей высотно-поясная дифференциация растительного покрова отражает комплексные изменения в составе, структуре и функционировании лесных сообществ, необходимость учета которых определена при их изучении в горах (Назимова и др., 1987; Бочарников, 2015). Индивидуальные для высотно-поясных подразделений уровни биоразнообразия, фоновый состав сообществ, характер естественной динамики, маркируемые климатическими показателями, при которых развиваются сообщества, обуславливают решение фундаментальных и практических биогеографических и лесоводственных задач с учетом высотной дифференциации растительного покрова (Назимова и др., 2020).
Картографический метод послужил в исследовании ключевым для выявления пространственной структуры лесного покрова Северобайкальского варианта оробиома. Создание инвентаризационной карты лесов оробиома, на основе которой проведен анализ разнообразия и структуры лесов, предварялось несколькими подготовительными этапами. На первом этапе карта растительности юга Восточной Сибири (Белов, 1973) переведена в цифровой формат. Привязка карты осуществлена на основе опорных точек по элементам гидросети и дорожной сети с использованием полиномиальной модели 6-ой степени в системе координат UTM WGS-84. Картографические выделы векторизованы, в атрибутивную таблицу добавлены поля с нумерацией, приведенной на исходной карте. На втором этапе подготовлен пакет растровых слоев цифровых карт, маскированных по границам в пределах площади, занимаемой географическим вариантом оробиома. В их число вошли оцифрованная карта растительности юга Восточной Сибири, цифровая карта растительного покрова России (Барталев и др., 2015) и цифровая модель рельефа ASTER GDEM. Все они приведены к единой географической проекции и пространственному разрешению опорной единицы анализа (пиксель цифрового изображения) 230 м. Фрагмент карты растительности России использован для отражения актуального состояния светлохвойных (лиственничные и сосновые), темнохвойных (еловые и пихтово-кедровые) и мелколиственных (березовые) лесов. Типологическое разнообразие фоновых сообществ в пределах формаций (уровень классов и групп ассоциаций) определено на основе карты растительности юга Восточной Сибири. Цифровая модель рельефа послужила основой для создания растровых слоев основных морфометрических характеристик (крутизна, экспозиция, кривизна склонов, относительное превышение, глубина заложения речных долин), характеризующих разнообразие экологических условий в соответствии со сложной орографической структурой. Они широко используются в качестве переменных при анализе разнообразия и пространственной структуры растительного покрова горных территорий, которые находят с ними тесные связи (Данилова и др., 2013; Рыжкова и др., 2016; Wiesmair et al., 2017).
Типологическое разнообразие лесов определено с учетом региональной специфики территории, необходимости его картографического отображения в мелком масштабе с оценкой высотно-поясных закономерностей дифференциации сообществ и динамических тенденций в развитии лесов. Всего выделено 11 фоновых типологических подразделений условно-коренных лесов в пределах светлохвойных, темнохвойных и мелколиственных формаций (5 – лиственничных лесов, 2 – сосновых, 2 – еловых, 1 – пихтово-кедровых и 1 – березовых). К условно коренным березовым лесам отнесены сообщества, участвующие в формировании серийных долинных комплексов сообществ в днище Верхнеангарской котловины. Совмещение двух карт растительности позволило отразить актуальное распространение лесов с типологическим разнообразием в пределах лесных формаций. Отдельно для каждой формации отображен неопределенный класс, чья типологическая приуроченность неизвестна в связи с нелесным статусом сообществ по состоянию на период 50-ти лет назад. Березовые леса горных склонов отнесены к длительно производным вариантам коренных светлохвойных и темнохвойных лесов, восстановительные сукцессии которых включают, как правило, стадию березового сообщества.
Созданный набор растровых слоев, представленный лесами, включающими 21 подразделение, и пятью морфометрическими характеристиками рельефа, позволил перейти непосредственно к цифровому картографированию лесов географического варианта оробиома. Интеграция исходных данных через функцию (леса) и определяющие ее переменные (орографические показатели) послужили основой для реализации пошагового дискриминантного анализа. Как метод классификации объектов и определения факторов, вносящих вклад в их разделение на группы, он находит свое применение в картографировании растительности, проводимом на основе точечных полевых материалов (обучающая выборка) и дистанционных данных (Попов, 2016). В настоящей работе в качестве обучающей выборки выступили известные типологические подразделения лесных формаций по данным предварительной цифровой карты. Преимущество метода заключается в возможности выявления оптимального сочетания факторов, объясняющих разграничение исходно определенных типологических подразделений. При этом дается оценка вклада каждого фактора, выявляемого с помощью переменных, и общая оценка анализа, которая является интегральным показателем качества анализа. На основе определения дискриминирующих функций возможна вероятностная оценка отнесения неизвестных подразделений к изначально определенным (максимальные значения вероятностей рассматриваются в качестве основного критерия для выбора соответствующего класса). Исходя из логики анализа, в пределах каждой лесной формации определено соответствие между их типологическими подразделениями и орографическими характеристиками (переменные) на пиксельном уровне, что послужило основой для выявления дискриминирующих функций и отнесения неопределенного класса к одному из известных. На основе сходства экотопических условий условно коренных лесов и развивающихся на их месте производных аналогичным образом выявлено типологическое разнообразие березовых лесов как производных вариантов горнотаежных лесов. В ходе анализа определены статистические показатели (Wilks’ Lambda, F-статистика, уровень значимости), характеризующие качество выделения соответствующих подразделений.
Анализ составленной карты лесов послужил основой для выявления пространственных закономерностей, выраженных в растительном покрове Северо-Восточного Забайкалья. Абсолютная высота послужила интегральным критерием для выявления высотной дифференциации типологического разнообразия лесов и редколесий в рамках горнотаежного пояса и нижней части подгольцового пояса. Для каждого подразделения определены основные статистические показатели, характеризующие связи с занимаемой высотой (медиана, квартили 25 и 75%, максимальные и минимальные значения). Проведенный дисперсионный анализ на основе непараметрического критерия Краскела-Уоллиса позволил определить значимость показателя абсолютной высоты в разделении лесов на группы (использованы H-статистика и уровень значимости). Степень различий между группами при попарном сравнении определена на основе z-критерия.
Пространственная структура лесного покрова в пределах рассматриваемой высотной амплитуды выявлена на основе анализа занимаемых типологическими подразделениями площадей. Определена общая для географического варианта лесистость, доли площадей каждой группы от общей площади территории и от лесопокрытой площади. Для двухсотметровых отрезков на высотном профиле в пределах горнотаежного пояса определено соотношение площадей, занимаемых сообществами, что послужило основой для выявления внутрипоясной организации разнообразия лесов.
Латинские названия сосудистых растений приведены по С.К. Черепанову (1995), лишайников – по списку лихенофлоры России (Список …, 2010). Пространственные операции с данными, в том числе сопряженный анализ растровых слоев, выполнены в геоинформационной среде программы ArcGIS (v. 10.2.2) с помощью набора инструментов анализа растровых поверхностей приложения ArcToolbox (aspect, slope, slope height, valley depth) и программы SAGA GIS (v. 2.1.4). Статистический анализ выполнен в программе Statistica (v. 12.5.192.5).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Создание карты лесов Северобайкальского варианта оробиома. Горнотаежный пояс в растительном покрове Северобайкальского варианта Северовосточно-Забайкальского оробиома образуют формации светлохвойных (лиственица Гмелина, сосна обыкновенная) и темнохвойных (ель сибирская, сосна сибирская, пихта сибирская) лесов с участием мелколиственных (береза плосколистная (Betula platyphylla), тополь душистый (Populus suaveolens), осина (P. tremula), кореянка земляничниколистная (Chosenia arbutifolia)). Их роль в структуре растительного покрова крайне неравнозначна, что обусловлено ботанико-географическими закономерностями, с которыми связана структура фитоценотического разнообразия оробиома. Мелкий масштаб проведенного картографирования определил возможность отображения типологических подразделений лесов (уровни классов и групп ассоциаций), которые оптимальны для характеристики фонового растительного покрова на региональном пространственном уровне организации экосистем. В первую очередь к ним относятся лиственничные леса ангаридского комплекса, доминирующее положение которых определяется занимаемой высотной амплитудой, площадью, разнообразием экотопических условий, в которых произрастают сообщества. При этом обширное географическое распространение, сложная орографическая структура и разнообразие климатопов обуславливают региональную специфику, определяющую возможности развития сообществ других формаций, относящихся к Урало-Южносибирскому комплексу. Особое положение занимают мелколиственные леса. Наиболее распространены на горных склонах березовые (береза плосколистная) с участием хвойных видов длительно производные сообщества, формирующиеся на месте коренных лиственничных лесов. В нижних частях долин рек березовые леса с участием тополя, осины, чозении, видов ив (ива Бебба (Salix bebbiana), ива козья (S. caprea)) выступают коренными сообществами, формируя пойменные серии с лиственнично-еловыми лесами.
Картографирование лесов на основе дискриминантного анализа по морфометрическим показателям рельефа выявило различия в точности отнесения сообществ в составе разных формаций, которая определялась по верно классифицированным подразделениям обучающей выборки (табл. 1). Они связаны с различными амплитудами экотопических условий, свойственных фоновым для высотного спектра лесам и лесам с ограниченным распространением, а также объемом выборки. Наибольшая точность определена для темнохвойных формаций, которые, занимая небольшую площадь, тяготеют к нижней части высотного спектра, вверх по нему распространяясь по речным долинам. Высокое качество дискриминации также имеют сосновые леса, строго приуроченные к нижней части горнотаежного пояса. Общая точность определения типов лиственничных лесов на основе дискриминантного анализа относительно невысока, что характеризует значительную вариабельность условий, в которых они могут произрастать (вся высотная амплитуда горнотаежного пояса и нижняя часть подгольцового пояса). Березовые леса как производные варианты имеют наименьшую точность отнесения к исходным условно коренным сообществам. Средняя точность составленной карты равна 62%.
Таблица 1.
Результаты дискриминантного анализа типологических подразделений лесов Северобайкальского варианта оробиома по морфометрическим показателям. Для типологических подразделений растительности приведены показатели F-статистики, Wilks’ Lambda, качества детерминации (%), число пикселей на карте (N). p-value: ** <0.000; * <0.05; – незначим при p < 0.05
| Морфометрические показатели рельефа | Лесные формации и группы формаций (Wilks’ Lambda) | |||
|---|---|---|---|---|
| Лиственничные N = 392 236 F (20130.1) = 6147 |
Сосновые N = 14 041 F (5.14035) = 379.07 |
Темнохвойные N = 2406 F (10.4798) = 125.52 |
Березовые N = 40 996 F (50.186905) = 437.47 |
|
| Абсолютная высота | 0.851617** | 0.887713** | 0.717650** | 0.707707** |
| Экспозиция склона | 0.743465** | 0.881294* | 0.639142** | 0.610771** |
| Крутизна склона | 0.751203** | 0.886132** | 0.628487– | 0.643187** |
| Относительное превышение | 0.760239** | 0.900170** | 0.671628** | 0.640302** |
| Глубина заложения долин | 0.763892** | 0.904950** | 0.634651** | 0.655485** |
| Точность выделения групп, % | 49 | 74 | 89 | 36 |
Для всех морфометрических показателей определен высокий вклад в разделение большинства типологических групп лесных сообществ (по критерию Wilks’ Lambda). В наибольшей степени это относится к абсолютной высоте, которая является ведущим показателем для разделения лиственничных, темнохвойных и березовых лесов, характеризуя высотные закономерности в их пространственной структуре (табл. 1). Крутизна склонов имеет значение значимости выше предельного уровня (при p < 0.05) при разделении темнохвойных лесов, которые, в целом, избегают крутых денудационных поверхностей.
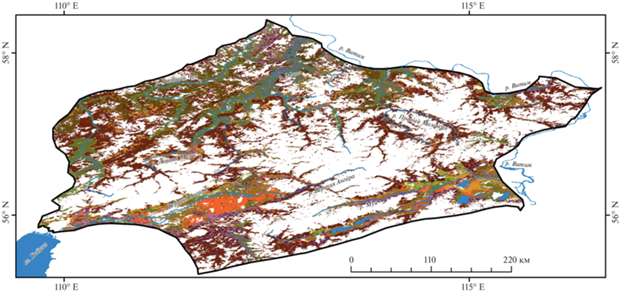
Полученная в результате анализа цифровая инвентаризационная карта лесов Северобайкальского варианта оробиома в мелком масштабе отразила актуальное состояние их типологического разнообразия и ключевые закономерности пространственной структуры, выраженные на горной территории (рис. 2).
Рис. 2.
Карта лесов Северобайкальского варианта Северовосточно-Забайкальского оробиома (м.
1 : 3 000 000).
Легенда (описание лесных сообществ – см. в тексте).
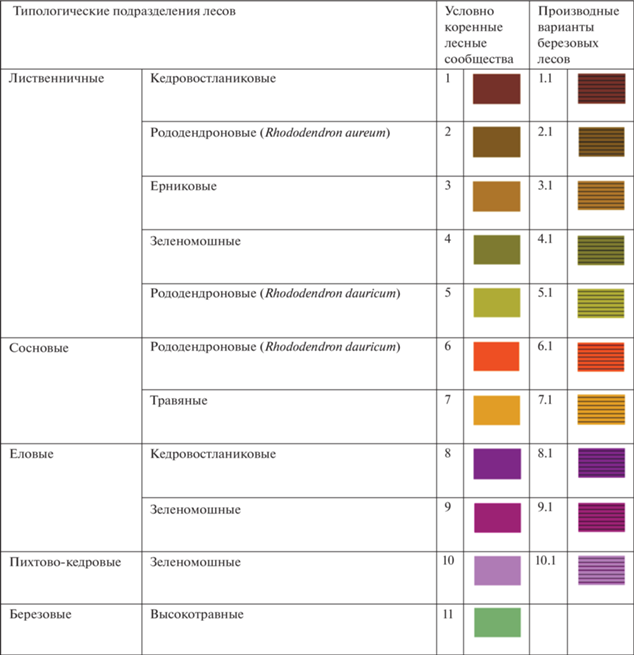
Фитоценотическое разнообразие лесов. Лиственничные леса распространены повсеместно в пределах горнотаежного пояса (300–900 м), а также в нижней части подгольцового пояса (900–1100 м), локально проникая выше 1500 м. Часто они характеризуются монодоминантными древостоями, на верхней границе леса – с примесью ели сибирской и кедра сибирского, в нижней части спектра – с участием пихты сибирской. Для большинства сообществ характерно развитие кустарникового яруса, небольшого по разнообразию травостоя и обычно выраженного мохового покрова. В составе формации лиственничных лесов выделено 5 типологических подразделений.
Лиственничные кедровостланиковые (кедровый стланик (Pinus pumila)) леса отражают тесные связи горнотаежного и подгольцового комплексов, характерных для растительного покрова Северо-Восточного Забайкалья. Постоянным элементом сообществ является кедровый стланик, формирующий более или менее сомкнутый подлесок, в котором он может принимать жизненную форму древовидного кустарника, иногда достаточно высокорослого (Моложников, 1975). Развитие кедровостланиковых лиственничных лесов демонстрирует связи растительного покрова Забайкалья и Дальнего Востока, где в горных районах он характеризуется наибольшим фитоценотическим разнообразием, являясь одним из основных элементов берингийского комплекса (Сочава, Лукичева, 1953), и где он формирует самостоятельную формацию с высоким уровнем разнообразия (Нешатаева, 2011). Достаточно широкая экологическая амплитуда вида при высокой требовательности к освещенности обуславливают развитие лиственничных лесов с его участием в подлеске. Вблизи верхней границы леса характерна примесь ели и кедра в разреженном древостое.
Лиственничные рододендроновые (рододендрон золотистый (Rhododendron aureum)) леса типологически близки к кедровостланиковым. Основу формирует другой компонент берингийского комплекса – рододендрон золотистый, монтанный вид с североазиатским распространением (Малышев, Пешкова, 1984). Он активно участвует в сообществах горнотаежного, подгольцового и горнотундрового поясов в Восточной Сибири и на Дальнем Востоке. Тяготея к верхней части высотных профилей, вид избегает повышенной континентальности климата, находя оптимальные условия для развития в еловых (ель аянская (Picea ajanensis)) лесах (Манько и др., 2020).
Лиственничные ерниковые (береза растопыренная (Betula divaricata), береза тощая (B. exilis)) леса имеют фоновое участие в растительном покрове горнотаежного пояса. Обычны чистые древостои сомкнутостью 0.3–0.6, местами – на зандровых равнинах, с участием сосны, на верхней границе леса – ели и кедра. Подлесок сообществ, высотой до 1.5 м и сомкнутостью до 0.8, разнообразен по составу. Наиболее часто встречается береза растопыренная, иногда совместно с березой шерстистой (Betula lanata), представляя широко распространенный в горах (и на равнинах в тундровой зоне) Восточной Сибири и Дальнего Востока монтанный элемент (Малышев, Пешкова, 1984). В высокогорьях этот вид проявляет высокую активность и формирует самостоятельную формацию.
Лиственничные зеленомошные леса имеют относительно небольшое распространение. При активном развитии мохового покрова в сообществах лиственничных лесов группа зеленомошных лесов характеризуется отсутствием кустарникового яруса или единичным участием ольхи кустарниковой (Duschekia fruticosa), кедрового стланика.
Лиственничные рододендроновые (рододендрон даурский (Rhododendron dauricum)) леса являются характерным компонентом растительного покрова для всего Забайкалья (Малышев, 1984; Сочава, 1980). В составе их древостоя преобладает лиственница с небольшой примесью сосны. Длительно производные варианты представлены березовыми сообществами, которые встречаются часто в связи с возникающими с высокой периодичностью пожарами. Отмечаемая для Чарской котловины конвергентность рододендроновых лиственничных лесов с сосновыми, выраженная в сходстве видового состава с обилием ксерофитов, развитии древостоя и подроста из лиственницы и сосны (Гаращенко, 1993), прослеживается и в других областях формирования оробиома, характеризуя контакт между подпоясами горнотаежного пояса, который приходится на высоты 500–800 м (Bocharnikov et al., 2018).
Сосновые леса имеют гораздо более ограниченное распространение, однако в нижнем подпоясе горнотаежного пояса, который развит в межгорных котловинах Байкальской рифтовой зоны на высотах 500–600 м, являются одними из фоновых (Bocharnikov et al., 2018). Наиболее конкурентоспособными сосновые леса по сравнению с лиственничными становятся в условиях песчаного состава грунтов со слабым промерзанием или его полным отсутствием, что в ландшафтной структуре оробиома имеет ограниченное распространение. Сообщества характеризуются монодоминантными древостоями, хорошо развитым подлеском и травяно-кустарничковым ярусом из бореальных и боровых видов, а также видов берингийского комплекса, произрастающих по всему высотному спектру.
Сосновые рододендроновые (рододендрон даурский) леса занимают доминирующие позиции в ценотическом разнообразии формации. Древесный ярус сообществ, образованный сосной, иногда с небольшой примесью лиственницы, обычно разреженный (сомкнутость – 0.3–0.5). Высота наиболее старовозрастных и продуктивных древостоев не превышает 25 м. Восстановление лесов после нарушений происходит без смены основной породы или с участием березы и лиственницы в зависимости от условий произрастания сообществ. Сосновые леса развиваются в условиях относительно высоких температур вегетационного периода (сумма активных температур выше 10°C – более 1400), которые обуславливают возможность их произрастания в нижних частях высотных спектров (Бочарников, 2019). Только за счет специфических условий, например, развития карбонатных пород, сосновые леса достигают абсолютных высот 1000 м и более, например, на Северо-Муйском хребте в верховьях долины реки Верхняя Ангара (оригинальные данные) и в истоках реки Келяны (Курнаев, 1973). Основные массивы рододендроновых сосновых лесов приурочены к зандровым поверхностям, вершинам основных и конечных морен в Верхнеангарской и Муйско-Куандинской котловинах, песчаным аллювиальным террасам рек (Гаращенко, 1993).
Сосновые травяные леса значительно уступают сообществам рододендроновой группы по роли в растительном покрове горнотаежного пояса. Для них характерно относительно высокое разнообразие травяно-кустарничкового яруса. Увеличение видового богатства связано с развитием лесостепных псаммофильных видов (прострел Турчанинова (Pulsatilla turczaninovii), осока аргунская (Carex argunensis), полынь пижмолистная (Artemisia tanacetifolia)). Травяные сосновые леса участвуют в формировании реликтовых лесостепных комплексов, встречающихся локально только на западе Станового нагорья – в низкогорьях Верхнеангарского хребта (Осипов, 2005).
Еловые, кедровые и пихтовые леса не являются фоновыми для оробиома, однако их развитие характеризует важные региональные черты, находящие отражение в структуре высотной поясности Северобайкальского географического варианта оробиома (Bocharnikov et al., 2018). При небольшой площади, занимаемой сообществами темнохвойных лесов, они встречаются на всей амплитуде горнотаежного пояса, в целом характеризуя повышенное увлажнение и сниженную континентальность климата (Бочарников, 2019). Наиболее распространенными являются еловые кедровостланиковые, еловые зеленомошные и кедровые зеленомошные леса.
Мелколиственные леса распространены по всей высотной амплитуде горнотаежного пояса, но не принимают фонового участия в какой-либо его части. Условно коренной характер имеют березовые высокотравные леса в долинах рек, наиболее характерные для днища Верхнеангарской котловины, где они формируют пойменные серии с участием разнообразных по типологическому составу сообществ (помимо лесов, это ерниковые заросли, гигрофильные луга). В составе ценофлор мелколиственных лесов долинных комплексов снижена роль берингийского комплекса, а наиболее характерными являются гигрофильные виды с широким распространением, находящие наибольшую ценотическую активность в лесах гор Южной Сибири (осока большехвостая (Carex macroura), дягиль низбегающий (Archangelica decurrens), таволга вязолистная (Filipendula ulmaria), какалия копьевидная (Cacalia hastata)). На склонах хребтов вплоть до верхней границы леса распространены березовые леса, представляющие длительно производные варианты сообществ, фоновых для горнотаежного пояса лиственничных лесов. Восстановительные сукцессии в них могут происходить без смены основной лесообразующей породы, но чаще с прохождением стадии березового древостоя, что определяется природой инвариантов.
Пространственная структура лесов горнотаежного пояса. Леса исследуемой территории распространены на площади около 39 тыс. км2, что составляет почти половину от общей площади, занимаемой географическим вариантом. Фоновое участие в высотно-поясном спектре занимают лиственничные леса (более 80% от лесопокрытой площади с учетом производных березовых лесов, формирующихся на их месте), представленные 5 типологическими подразделениями (табл. 2). Они распространены по всей высотной амплитуде горнотаежного пояса, заходя также в нижнюю часть подгольцового пояса. В пределах данных поясов пространственная структура лесных сообществ определяется комплексом экологических факторов, которые связаны с теплообеспеченностью, увлажнением, мощностью мерзлоты, составом горных пород, что не может быть в полной мере интерпретировано через орографическую структуру территории. В структуре разнообразия лесов находят выражение также региональные черты. Характерным примером выступают лиственничные рододендроновые с рододендроном золотистым (Rhododendron aureum) леса, приуроченные к наиболее увлажненной северо-западной части Северо-Байкальского нагорья, где они связаны с верхней частью горнотаежного пояса (верховья долин рек Верхняя Чуя, Левая Мама). Климатопы этих лесов характеризуются сниженной континентальностью климата (Bocharnikov, Stas’ko, 2018).
Таблица 2.
Пространственная структура лесов на высотном градиенте спектра Северобайкальского варианта оробиома. В ячейках для каждой высотной ступени приведена доля площади, занимаемая типологическими подразделениями лесов от лесопокрытой площади. Типологические подразделения лесов – см. рис. 2
| Абсолютная высота, м | Типологические подразделения лесных сообществ | Лесистость, % | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 1600–1800 | 1 | |||||||||||
| 1400–1600 | 5 | |||||||||||
| 1200–1400 | 18 | |||||||||||
| 1000–1200 | 40 | |||||||||||
| 800–1000 | 65 | |||||||||||
| 600–800 | 85 | |||||||||||
| 400–600 | 87 | |||||||||||
| 200–400 | 78 | |||||||||||
| Площадь, тыс. км2 | 15.5 | 4.4 | 3.9 | 7.2 | 1.1 | 2.0 | 2.2 | 0.8 | 0.003 | 1.6 | 0.6 | 39 |
| Доля от лесопокрытой площади, % | 40.1 | 11.2 | 10.0 | 18.6 | 2.8 | 5.1 | 5.6 | 2.2 | 0.007 | 4.2 | 0.2 | 100 |
| Доля от общей площади, % | 18.6 | 5.2 | 4.6 | 8.7 | 1.3 | 2.4 | 2.6 | 1.0 | 0.003 | 2.0 | 0.08 | 47 |
| Доля площади лесов от лесопокрытой площади на высотных уровнях, % | |
|---|---|
| 80 < 100 | |
| 60 < 80 | |
| 40 < 60 | |
| 20 < 40 | |
| 10 < 20 | |
| 1 < 10 | |
| 0 < 1 | |
| 0 |
Второстепенное участие в сложении высотно-поясной структуры растительного покрова, характерное для сосновых (более 10% от общей площади лесов), пихтово-кедровых (более 2%) и еловых (более 4%) лесов, определяет их строгую экотопическую приуроченность, включая связи с высотным уровнем, склонами определенной экспозиции и крутизны. Все сообщества тяготеют к более теплообеспеченным условиям нижней части горнотаежного пояса (табл. 2). Распространение вплоть до верхней границы леса связано преимущественно с речными долинами, а также с развитием горных пород, прежде всего, карбонатов, способствующих развитию сообществ за пределами климатического оптимума их распространения. Менее 1% площади приходится на долинные комплексы березовых лесов.
Характер распределения лесных сообществ по высотному градиенту характеризует неоднородность пространственной структуры фитоценотического разнообразия, выраженного в пределах поясов. Выделенные типологические подразделения существенно отличаются в соответствии с приуроченностью к определенной части на высотном спектре (рис. 3). Абсолютная высота, взятая в качестве интегрального критерия, показала высокую значимость при разделении лесов (H (10, N = 731312) = 229998.8, p = 0.000). За исключением еловых зеленомошных лесов, распространенных по долинам рек более равномерно по всему спектру, все типы достоверно различаются между собой при попарном сравнении занимаемых высот, обосновывая возможность выявления внутрипоясной структуры горнотаежного пояса.
Рис. 3.
Высотное распределение типологических подразделений лесов. Типологические подразделения – см. рис. 2.
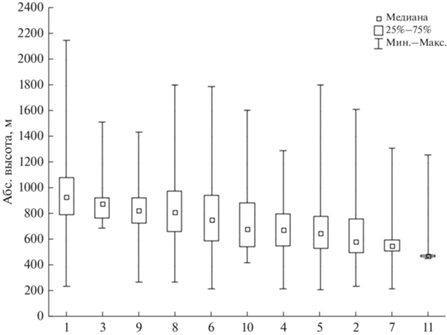
В соответствии с оптимальным развитием типологических подразделений лесов на соответствующих высотах возможна дифференциация горнотаежного пояса. В качестве косвенного показателя оптимального либо ограниченного участия сообществ взята занимаемая площадь, которая изменяется в связи с высотным положением (табл. 2). На всех высотных уровнях доминирующие позиции занимают лиственничные леса. До высоты 800 м преобладают лиственничные зеленомошные, ерниковые и рододендроновые (рододендрон даурский) леса, а на высотах 800–1000 м – лиственничные кедровостланиковые. Последние формируют верхнюю границу леса, проникая отдельными фрагментами в подгольцовый пояс на высоты более 1400 м, где общая лесистость не превышает 5%. Оптимум развития сосновых лесов приходится на 400–600 м (преобладают сосновые рододендроновые леса), где лесистость превышает 80%. Наибольшие площади они занимают в межгорных котловинах и низкогорьях хребтов на мощных аллювиальных отложениях. Сосновые травяные леса, несмотря на значительную высотную амплитуду, находят экотопически пригодные местообитания на небольшой площади, преимущественно в нижней части спектра. Их ограниченное развитие характеризует региональные особенности оробиома, определяя его отличия от окружающих равнин и горных территорий (Приангарье, Южное Забайкалье) с фоновой ролью этих сообществ в растительном покрове. Темнохвойные леса при общем небольшом распространении достаточно равномерно распределены по спектру, занимая всю амплитуду горнотаежного и подгольцового поясов. На каждом высотном уровне на них приходится до 5–10% от лесопокрытой площади.
Высотная дифференциация структуры лесного покрова, дающая интегральную оценку изменения биотического и абиотического компонентов экосистем в рамках горнотаежного пояса, обуславливает возможность выделения высотных подпоясов и полос. На высотах 300–600 м формируется нижний подпояс сосновых, сосново-лиственничных и темнохвойных с участием березовых лесов. На высотах 600–1000 м развит верхний подпояс лиственничных лесов. Подпояс лиственничных лесов включает две высотные полосы: зеленомошных и рододендроновых (рододендрон даурский) лиственничных лесов (600–800 м) и кедровостланиковых и ерниковых лиственничных лесов (800–1000 м). Фоновые типы лиственничных сообществ распространены повсеместно и характерны для всех горных хребтов, имея варьирование в оптимуме развития на высотном спектре. Верхняя полоса характеризуется преобладанием кедровостланиковых лиственничных лесов с участием ерниковых. В формировании верхней границы леса (около 800–1000 м и выше) участвуют кедровостланиковые лиственничные леса. В нижней полосе (600–800 м) преобладают лиственничные рододендроновые леса с участием сосновых лесов. В нижнем подпоясе горнотаежного пояса сохраняется фоновая роль лиственничных лесов, однако увеличивается общий уровень фитоценотического разнообразия, увеличивается роль сосновых лесов, особенно рододендроновой группы. В рамках подпояса выражена региональная специфика. В соответствии с ней в низкогорьях хребтов Северо-Байкальского нагорья, расположенного в северной части географического варианта оробиома, он занимает более низкие отметки высот (300–500 м). В пределах Байкальской рифтовой зоны он приурочен к нижним частям межгорных котловин (500–600 м).
Высотно-поясная организация фитоценотического разнообразия в горнотаежном поясе Северо-Восточного Забайкалья отражается в комплексе фитоценотических показателей лесных сообществ с оптимальным развитием в пределах подпоясов и полос. Интегральное выражение, высотная дифференциация фитоценотического разнообразия находятся в комплексе показателей. В их число входят структурно-динамические характеристики растительности, функциональная роль сообществ и абиотические факторы, которые определяют пространственные закономерности их развития (табл. 3). Нижний подпояс сосновых и сосново-лиственничных лесов не имеет повсеместного распространения и выражен фрагментарно в днищах Верхнеангарской и Муйско-Куандинской котловин и в низкогорных частях хребтов, расположенных ниже 600 м, занимая около 15% от площади горнотаежного пояса. Фоновую роль в его разнообразии играют сосновые рододендроновые леса с небольшой примесью сосновых травяных лесов. Для нижнего подпояса характерно развитие склоновых комбинаций сосновых и лиственничных лесов, в речных долинах формируются серийные сообщества с участием березовых, осиновых и еловых лесов. В поймах рек бассейна Верхней Ангары формируются долинные комплексы с участием березовых и осиновых высокотравных лесов. Сосновые леса, сильно подверженные пожарам, восстанавливаются без смены основной лесообразующей породы (Гаращенко, 1993).
Таблица 3.
Интегральная схема высотной дифференциации фитоценотического разнообразия лесов горнотаежного пояса Северобайкальского варианта оробиома
| Высотный подпояс (абсолютные высоты, м) | Высотная полоса (абсолютные высоты, м) | Фоновые и второстепенные лесные сообщества | Преобладающие функции сообществ; основные дестабилизирующие факторы | Преобладающий ход естественной динамики сообществ |
|---|---|---|---|---|
| Верхний
лиственничных лесов (600–1000 м) |
Верхняя лиственничных кедровостланиковых и ерниковых лесов (800–1000 м) | Лиственничные кедровостланиковые, ерниковые, рододендроновые (Rhododendron aureum), зеленомошные | Водосборная и противоденудационная функции; пожары | Смены на коротко производные лиственничные леса, длительно производные ерниковые сообщества и березовые леса |
| Нижняя лиственничных зеленомошных и рододендроновых лесов (600–800 м) | Лиственничные зеленомошные, рододендроновые (Rhododendron dauricum), кедровостланиковые | Противоденудационная функция; пожары, рубки | Смены на длительно производные березовые леса и ерниковые сообщества | |
| Нижний
сосновых,
сосново-лиственничных и темнохвойных лесов (300–600 м) |
Сосновых и сосново-лиственничных рододендроновых и темнохвойных зеленомошных лесов (300–600 м) | Сосновые рододендроновые (Rhododendron dauricum), лиственничные зеленомошные, пихтово-кедровые зеленомошные, березовые высокотравные | Противоэрозионная и водорегулирующая функции; пожары, рубки, водная эрозия | Смены на коротко производные сосновые леса, длительно производные березовые леса и луга |
Верхний подпояс лиственничных лесов является фоновым для горнотаежного пояса и всей высотно-поясной структуры растительного покрова в целом. Он выражен на высотах 600–1000 м, характеризуясь преобладанием кедровостланиковых и ерниковых сообществ. Для нижней полосы характерны склоновые комбинации с сосновыми лесами, для верхней – с зарослями кедрового стланика, а на наиболее увлажненных хребтах – с березовыми (береза шерстистая) криволесьями. Произрастая в широком спектре экотопических условий, лиственничные леса занимают разные уровни фитокатен на горных склонах. Восстановление сообществ после рубок и пожаров идет через стадии длительно производных березовых лесов.
По высотному градиенту в пределах горнотаежного пояса прослеживается изменение не только типологического состава лесных сообществ, но и комплекса характеристик экосистем, обнаруживающих высотно-поясную организацию. Среди них выделяется функциональная роль сообществ, характеризующихся высоким разнообразием выполняемых функций в связи со сложной экотопической структурой горной территории и высотной дифференциацией преобладающих функций (Огуреева, Бочарников, 2020). В верхнем подпоясе ключевую роль играют водосборная и противоденудационная функции. В нижнем подпоясе важнейшее значение приобретают противоэрозионная и водорегулирующая функции. Высотная дифференциация прослеживается и в отношении влияющих на леса дестабилизирующих факторов, которая связана как с природными, так и антропогенными процессами. Наиболее обширное и практически повсеместное влияние на лесные экосистемы оказывают пожары. По состоянию на 2015 г. около 2% территории Северо-Восточного Забайкалья была занята свежими гарями (Барталев и др., 2015). Основной объем рубок леса связан с низкогорьями хребтов и межгорными котловинами, наиболее доступными в связи с орографической структурой территории и в отношении развития транспортной инфраструктуры. На лесные сообщества нижнего подпояса горнотаежного пояса важное влияние оказывает также водная эрозия, характерная для долинных комплексов.
ЗАКЛЮЧЕНИЕ
Интенсификация хозяйственной деятельности в отдаленных горных районах Восточной Сибири и Дальнего Востока, особенно вдоль трассы Байкало-Амурской магистрали, трансформация природных экосистем вследствие, прежде всего, рубок леса и возрастание рисков негативных явлений катастрофического характера (прежде всего, лесные пожары) происходят возрастающими темпами. В соответствии с этим требуются новые знания о растительном покрове и лесах как фоновом компоненте в высотно-поясной структуре типов поясности бореального класса. Имеющиеся данные о фитоценотическом разнообразии и пространственной структуре лесов горных территорий вдоль БАМа не дают целостного представления о современном состоянии разнообразия лесного покрова. Положенная в основу настоящего исследования концепция экосистемного разнообразия на региональном уровне позволила провести оценку фитоценотического разнообразия и пространственной организации лесов с учетом высотно-поясной структуры растительного покрова Северо-Восточного Забайкалья. С помощью картографического метода получены новые данные об эколого-географических закономерностях дифференциации лесов, отражающих историю формирования растительного покрова и современное разнообразие условий в системе сложной орографической обстановки горной территории.
Впервые составленная на территорию инвентаризационная мелкомасштабная карта лесов (м. 1 : 3 000 000) отразила их актуальное разнообразие в системе фоновых для горнотаежного пояса типологических подразделений. Фитоценотическое разнообразие бореальных лесов Северобайкальского варианта Северовосточно-Забайкальского оробиома представлено условно-коренными сообществами 11 типологических подразделений светлохвойных (лиственничных и сосновых), темнохвойных (пихтово-кедровых и еловых) и мелколиственных (березовых) лесов на уровне классов и групп ассоциаций и их производными вариантами. Выделенные группы существенно различаются по занимаемой площади (при общей лесистости около 50%), высотной амплитуде и интегральной роли в структуре растительного покрова, характеризуя выраженную региональную специфику всего высотно-поясного спектра.
Проведенное исследование представляет собой важный этап в решении фундаментальных проблем географии биоразнообразия горных территорий, а также практических вопросов ботанической географии и лесоведения, связанных с мониторингом состояния лесов, их охраной и оптимизацией использования лесных ресурсов Северо-Восточного Забайкалья.
Список литературы
Барталев С.А., Егоров В.А., Ершов Д.В., Исаев А.С., Лупян Е.А., Плотников Д.Е., Уваров И.А. Спутниковое картографирование растительного покрова России по данным спектрорадиометра MODIS // Современные проблемы дистанционного зондирования Земли из космоса. 2011. Т. 8. № 4. С. 285–302.
Барталев С.А., Егоров В.А., Жарко В.О., Лупян Е.А., Плотников Д.Е., Хвостиков С.А. Состояние и перспективы развития методов спутникового картографирования растительного покрова России // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 5. С. 203–221.
Белов А.В. Карта растительности юга Восточной Сибири. Принципы и методы составления // Геоботаническое картографирование. Л.: Наука, 1973. С. 16–30.
Бочарников М.В. Эколого-фитоценотическая структура лесного покрова северного макросклона Западного Саяна // Лесоведение. 2015. № 1. С. 10–19.
Бочарников М.В. Региональные черты в географии флористического разнообразия Станового нагорья // Проблемы ботаники Южной Сибири и Монголии. Барнаул: изд-во АлтГУ. 2018. С. 21–25.
Бочарников М.В. Роль климата в пространственной организации растительного покрова Кодаро-Каларского оробиома // Сибирский экологический журн. 2019. Т. 26. № 3. С. 239–252.
Владимиров И.Н., Софронов А.П., Сороковой А.А., Кобылкин Д.В., Фролов А.А. Структура растительного покрова западной части Верхнеангарской котловины // География и природные ресурсы. 2014. Вып. 35. № 2. С. 44–53.
Гаращенко А.В. Флора и растительность Верхнечарской котловины (Северное Забайкалье). Новосибирск: ВО “Наука”. Сибирская издательская фирма, 1993. 280 с.
Данилова И.В., Рыжкова В.А., Корец М.А. Алгоритм автоматизированного картографирования современного состояния и динамики лесов на основе ГИС // Вестник Новосибирского государственного университета. Серия: Информационные технологии. 2010. Т. 8. № 4. С. 15–24.
Зорин Ю.А. Изостазия и новейшая структура Байкальской рифтовой зоны и сопредельных территорий. М.: Наука, 1971. 168 с.
Курнаев C.Ф. Лесорастительное районирование СССР / АН СССР. Лаб. лесоведения. М.: Наука, 1973. 203 с.
Лавренко Н.Н. Опыт составления карты ландшафтнозащитных и ресурсных функций растительного покрова зоны Байкало-Амурской магистрали // Геоботаническое картографирование. Л.: Наука, 1977. С. 20–32.
Малышев Л.И., Пешкова Г.А. Особенности и генезис флоры Сибири (Предбайкалье и Забайкалье). Новосибирск: Наука, 1984. 264 с.
Манько Ю.И., Гладкова Г.А., Сибирина Л.А. Леса из ели аянской с подлеском из рододендрона золотистого // Лесоведение. 2020. № 5. С. 412–423.
Моложников В.Н. Кедровый стланик горных ландшафтов Северного Прибайкалья. М.: Наука, 1975. 203 с.
Назимова Д.И., Коротков И.А., Чередникова Ю.С. Основные высотно-поясные подразделения лесного покрова в горах Южной Сибири и их диагностические признаки // Чтения памяти В.Н. Сукачева. М.: Наука, 1987. С. 30–64.
Назимова Д.И., Пономарев Е.И., Степанов H.B., Федотова Е.В. Черневые темнохвойные леса на юге Красноярского края и проблемы их обзорного картографирования // Лесоведение. 2005. № 1. С. 12–18.
Назимова Д.И., Пономарев Е.И., Коновалова М.Е. Роль высотно-поясной основы и дистанционных данных в задачах устойчивого управления горными лесами // Лесоведение. 2020. № 1. С. 3–16.
Нешатаев Ю.Н. Методы анализа геоботанических материалов. Л.: изд-во ЛГУ, 1987. 188 с.
Нешатаева В.Ю. Сообщества кедрового стланика (Pinus pumila (Pall.) Regel) полуострова Камчатка // Растительность России. 2011. № 19. С. 71–100.
Огуреева Г.Н. Ботанико-географическое районирование СССР. М.: МГУ, 1991. 78 с.
Огуреева Г.Н., Бочарников М.В. Оробиомы как базовые единицы региональной оценки биоразнообразия горных территорий // Экосистемы: экология и динамика. 2017. Т. 1. № 2. С. 52–81.
Огуреева Г.Н., Бочарников М.В. Экологические функции современного растительного покрова Станового нагорья // Вестник СПбГУ. Науки о Земле. 2020. Т. 65. № 2. С. 377–394.
Огуреева Г.Н., Леонова Н.Б., Емельянова Л.Г. и др. Карта “Биомы России” (М. 1 : 7 500 000) в серии карт природы для высшей школы. М.: WWF. 2018. 1 л.
Осипов К.И. Флора Витимского плоскогорья (Северное Забайкалье). Улан-Удэ: БНЦ СО РАН, 2005. 216 с.
Панарин И.И., Митрофанов Д.П., Исаева Л.Н. Горные леса зоны БАМ. Новосибирск: Наука, 1980. 214 с.
Пешкова Г.А. Растительность Сибири (Предбайкалье и Забайкалье). Новосибирск: Наука, 1985. 145 с.
Поликарпов Н.П., Чебакова Н.М., Назимова Д.И. Климат и горные леса Южной Сибири. Новосибирск: Наука, 1986. 225 с.
Попов С.Ю. Опыт создания геоботанической карты методом дискриминантного анализа полевых и дистанционных данных // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 13. № 1. С. 25–35.
Русяева Г.Г. Краткий очерк растительности центральной части Байкало-Патомского нагорья // Флора и растительность высокогорий. Новосибирск: Наука, 1979. Т. XIV. Вып. 1. С. 110–117.
Рыжкова В.А., Данилова И.В., Корец М.А. Классификация лесного покрова и геоинформационное моделирование его восстановительной динамики (на примере южной части приенисейской Сибири) // Сибирский экологический журн. 2016. Т. 23. № 6. С. 817–831.
Рысин Л.П. Лиственничные леса России М.: Изд-во КМК, 2010. 344 с.
Софронов А.П. Геоботаническое картографирование растительного покрова котловин Северо-Восточного Прибайкалья // Геоботаническое картографирование. 2015. С. 62–77.
Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 190 с.
Сочава В.Б. Географические аспекты сибирской тайги. Новосибирск: Наука, 1980. 256 с.
Сочава В.Б., Лукичева А.Н. К географии кедрового стланика // Докл. АН СССР. 1953. Т. 90. № 6. С. 1163–1166.
Список лихенофлоры России / Сост. Урбанавичюс. СПб.: Наука, 2010. 194 с.
Сукачев В.Н. Руководство к исследованию типов леса. М.–Л.: Гос. изд-во с.-х. и колхоз.-кооп. лит-ры, 1931. 328 с.
Сукачëв В.Н., Зонн С.В. Методические указания к изучению типов леса. М.: АН СССР, 1961. 144 с.
Типы лесов Сибири / Под ред. Смагина В.Н. Вып. 2. Красноярск: Красноярский рабочий, 1969. 280 с.
Чебакова H.M., Рейфельдт Дж., Парфенова Е.И. Перераспределение растительных зон и популяций лиственницы сибирской и сосны обыкновенной в Средней Сибири при потеплении климата. Сибирский экологический журн. 2003. № 6. С. 677–686.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
Bocharnikov M.V., Ogureeva G.N., Luvsandorj J. Regional features of the altitudinal gradients in Northern Transbaikalia vegetation cover // Geography, Environment, Sustainability. 2018. V. 11. № 4. P. 67–84.
Bocharnikov M.V., Stas’ko A.A. Spatial structure of the Kodar-Kalar orobiome botanical diversity on bioclimatic basis // BIO Web of Conferences. EDP Sciences. 2018. P. 1–4.
Johnson K.G., Brooks S.J., Fenberg P.B., Glover A.G., James K.E., Lister A.M., Michel E., Spencer M., Todd J.A., Valsami-Jones E., Young J.R., Stewart J.R. Climate change and biosphere response: Unlocking the collections vault // BioScience. 2011. V. 61. № 2. P. 147–153.
Kharuk V.I., Im S.T., Petrov I.A., Golyukov A.S., Ranson K.J., Yagunov M.N. Climate-induced mortality of Siberian pine and fir in the Lake Baikal Watershed, Siberia // Forest Ecology and Management. 2017. V. 384. P. 191–199.
Puzachenko M.Yu., Chernenkova T.V. Assessment of the vegetation cover conditions for the central part of the Murmansk region based on field and remote sensing data // Geography, Environment, Sustainability. 2012. V. 5. № 3. P. 4–13.
Vetaas O.R., Grytnes J.A. Distribution of vascular plants species richness and endemic richness along the Himalayan elevation gradient in Nepal. Global Ecology and Biogeography. 2002. № 11. P. 291–301.
Walter H., Breckle S.-W. Okologishe Grundlagen in global sicht. Stuttgart: G. Fischer, 1991. 586 p.
Wiesmair M., Otte A., Waldhardt R. Relationships between plant diversity, vegetation cover, and site conditions: implications for grassland conservation in the Greater Caucasus // Biodiversity and Conservation. 2017. V. 26. № 2. P. 273–291.
Дополнительные материалы отсутствуют.