

Микробиология, 2019, T. 88, № 2, стр. 154-164
Фототрофные сообщества высокоминерализованных мезотермальных сульфидных берикейских источников (Дагестан)
В. М. Горленко a, *, Е. И. Бурганская a, И. А. Брянцева a
a Институт микробиологии им. С.Н. Виноградского, Федеральный исследовательский центр “Фундаментальные основы биотехнологии” Российской академии наук
119071 Москва, Россия
* E-mail: vgorlenko@mail.ru
Поступила в редакцию 07.09.2018
После доработки 21.11.2018
Принята к публикации 30.11.2018
Аннотация
Впервые исследованы фототрофные сообщества, сформированные в ложе мезотермальных высокоминерализованных сульфидных Берикейских источников (Каякентский район Дагестана). Берикейские источники представляют интерес, как пример комплексного воздействия на распространение и структуру фототрофных микробных сообществ таких факторов, как температура, соленость, рН и сульфид. Вода источников хлоридно-натриевая, имела минерализацию 48–97 г/л, рН рассола был близок к нейтральному, концентрация сульфида около 1 мМ. Температура в местах излива источников достигала 60°С. Повышенная температура и высокая минерализация ограничивали развитие фототрофных сообществ. Цианобактериальные маты начинали формироваться ниже 54°С. В микробных матах преобладали Phormidium-подобные цианобактерии и одноклеточные Synechocystis sp. При распреснении до 48–57 г/л количество видов цианобактерий увеличивалось, добавлялись виды морфологически сходные с Spirulina sp., Leptolyngbya sp. и Oscillatoria sp. Аноксигенные фототрофные бактерии были представлены галофильными пурпурными бактериями Ectothiorhodospira sp., Marichromatium sp., Rhodovulum sp., зелеными серобактериями Prosthecochloris sp. и не идентифицированными Chloroflexi. Кислородный фотосинтез в матах не ингибировался сульфидом. Продуктивность цианобактериальных матов достигала 4.7–53.8 мкг/(см2 ч). Вклад аноксигенных фототрофных бактерий в фотосинтезирующую продукцию варьировал от 0 до 100%. Проведено сравнение изученного микробного состава с составом фототрофных сообществ соленых мезотермальных источников побережья Мертвого моря (Израиль), теплого озера Вашингтон (США) и горячих источников на острове Паоха в акватории содового озера Моно Лейк (США).
На территории Дагестана находятся источники разной степени минерализации и температуры. Термальные воды Дагестана очень разнообразны по своему химическому составу. Они приурочены к юрским, меловым и чокракским водоносным горизонтам, представленным трещиноватыми известняками, песчаниками и доломитами. В зависимости от условий образования, источники разделяются на хлоридно-натриевые, хлоридно-кальциевые, гидрокарбонатно-сульфатно-натриевые, сульфатно-гидрокарбонатно-хлоридно-натриевые (Дейнега, 1978; Курбанов, 2001). Ювенильные газы в источниках Дагестана представлены в основном аммиаком, азотом, углекислым газом, сероводородом, водородом и метаном.
Мезотермальные серные источники Дагестана делятся на низкоминерализованные и высокоминерализованные. К низкоминерализованным источникам относится высокосульфидный источник в районе курорта Талги (18 км от г. Махачкала), подробно изученный нами с позиций состава и геохимической активности цианобактериальных матов (Старынин и соавт., 1990). Температура в источнике не превышает 28°C. При общей минерализации 5 г/л концентрация сульфида в данном источнике достигает 218 г/л. Основу микробного мата в источнике Талги составляют цианобактерии рода Oscillatoria. Аноксигенные фототрофные бактерии (АФБ) представлены пресноводными видами: пурпурными серобактериями рода Thiospirillum, несерными пурпурными бактериями рода Rhodobacter, зелеными серобактериями рода Chlorobium и мезофильными нитчатыми бактериями Oscillochloris trichoides.
Берикейские термальные источники (42.3338889° с.ш., 48.0961111° в.д.), существенно отличаются по ряду физико-химических параметров от других гидротерм Дагестана, прежде всего, высоким содержанием солей в изливающейся воде. Промышленные запасы термальных вод Берикейского месторождения, вскрытые скважинами на площади Каякент, составляют 15 тыс. м3/сут, или 5.5 млн м3/г. Из них 1650 м3/сут, или 0.6 млн м3/год самопроизвольно выбрасываются на поверхность. Изливающиеся рассолы принадлежат к хлоридно-натриевому типу с общей минерализацией до 97 г/л. Они также содержат значительное количество ионов кальция, сульфата, бикарбоната и биогенных элементов (бром, йод, бор). Содержание сероводорода варьирует от 30 до 80 мг/л. Температура изливающихся рассолов доходит до 60°C, а рН близок к нейтральному.
Состав фототрофных сообществ мезотермальных источников Берикея ранее не изучался. Это послужило причиной проведения нами комплексных исследований, начатых в 1984 г. Окончательная обработка полевого материала и идентификация выделенных культур АФБ была завершена в 2018 г. Следует отметить, что в литературе опубликовано мало данных о составе и активности фототрофных сообществах мезотермальных минерализованных сульфидных источников (Cohen et al., 1977; Горленко, 2007; Thiel et al., 2017). Берикейские источники представляют интерес как пример комплексного воздействия таких факторов как температура, соленость, рН и сульфид на распространение и структуру фототрофных микробных сообществ.
В задачи настоящей работы входило: 1) определить физико-химические параметры изливающихся рассолов в местах формирования микробного мата; 2) выявить факторы, ограничивающие развитие цианобактериальных матов и обнаруженных видов цианобактерий и АФБ; 3) определить пигменты фотосинтеза и интенсивность оксигенного и аноксигенного фотосинтеза in situ; 4) определить численность основных групп аноксигенных фототрофов методом предельных разведений на элективных средах; 5) выделить и идентифицировать доминирующие АФБ, участвующие в процессе окисления сульфида.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Характеристика источников. Исследованные Берикейские гидротермы разделены нами на две подгруппы (табл. 1). Первая подгруппа расположена в двухстах метрах от побережья Каспийского моря (номера точек 7, 8 и 9, 11). Это экстремально соленая система с минерализацией воды до 97 г/л. Ко второй подгруппе относятся источники (номера точек 12–16 и 19), находящиеся на мысе Башлы в сбросовом канале гиперсоленого озера Аджи. Вода канала характеризуется меньшей соленостью (48–57 г/л) и более низкими температурами.
Таблица 1.
Общая информация о сульфидных Бирекейских источниках
| Параметры | I подгруппа | II подгруппа | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 7* | 8 | 9 | 11 | 12 | 13 | 14 | 15 | 16 | 19 | |
| Температура, °C | 49 | 44.5 | 50 | 40.5 | 40.5 | 38.5 | 51 | 34 | 44 | 28 |
| H2S, мг/л | 39.4 | 37.7 | 5.78 | 27.9 | 18.7 | 15.6 | 40.8 | 37.4 | 27.2 | НО |
| Соленость, г/л | 97 | 97 | 97 | 97 | 57 | 57 | 68 | 68 | 78 | 48 |
| O2, мг/л | 1.08 | 1.82 | 1.66 | 0.41 | 0 | НО | 0 | 0.83 | 1.2 | НО |
| pH | 6.15 | 6.1 | 5.75 | 6.35 | 7.05 | 7.4 | 6.7 | 7.62 | 6.7 | 6.2 |
| Eh воды, мВ | –85 | –100 | –60 | –160 | –80 | –5 | –20 | +60 | –60 | НО |
| ${\text{HCO}}_{3}^{ - }$ | 732 | 640.5 | 561 | 634 | 2110 | 1961 | 2066 | 2097 | 2268 | НО |
| S/${\text{SO}}_{3}^{{2 - }}$ | 260 | 290 | 400 | 310 | 160 | 160 | 180 | 200 | 190 | НО |
| Хлорофилл a, мкг/см2 | НО** | НО | НО | НО | 275 | 83 | 89 | НО | 335 | НО |
| Бактериохлорофилл c, мкг/см2 | НО | НО | НО | НО | 88 | 28 | 17 | НО | 38 | НО |
| Бактериохлорофилл a, мкг/см2 | НО | НО | НО | НО | 94 | 94 | 94 | НО | 94 | НО |
| Общая продукция фотосинтеза, мкг/см2 ч | 4.7 | 4.8 | 10.8 | 5.6 | 23 | 12.2 | 7.8 | НО | 53.8 | НО |
| % бактериального фотосинтеза | 61.7 | 100 | 17.6 | 14.3 | 24.3 | 0 | 17.9 | НО | 0 | НО |
| Результаты посевов*** | ||||||||||
| Rhodovulum sp. | – | 3 | + | – | 5 | 7 | – | 5 | 6 | 6 |
| Ectothiorhodospira sp. | 4 | 3 | 3 | 4 | 5 | 7 | 5 | 5 | 5 | |
| Marichromatium sp. | – | – | + | – | – | – | – | 5 | ||
| Prosthecochloris sp. | 5 | 4 | – | – | 4 | 5 | 2 | 5 | 5 | 6 |
| Chloroflexus sp. | + | + | ||||||||
В местах отбора проб соленость колебалась от 48 до 97 г/л, содержание сульфида было в пределах 27–40 мг/л, температура рассола варьировала от 27 до 51°C, рН 5.75–7.62. Окислительно-восстановительный потенциал воды во всех исследованных точках – отрицательный, ввиду присутствия сероводорода (–80…–160 мВ). Кислород присутствовал в воде только в дневное время непосредственно над поверхностью мата и, очевидно, был продуктом оксигенного фотосинтеза.
Определение физико-химических условий в местах формирования микробных матов. Общую минерализацию определяли по электропроводности солемером, калиброванным по стандартным растворам хлорида натрия. Содержание карбонатов и гидрокарбонатов определяли титрованием 0.1 н HCl с использованием индикаторов фенолфталеина и метилового оранжевого. Сульфид определяли йодометрическим титрованием (Резников и соавт., 1970). Концентрацию ${\text{SO}}_{4}^{{2 - }}$ определяли нефелометрически: к 0.5 мл пробы прибавляли 2 капли 30%-ой уксусной кислоты и 3.5 мл 0.5%-го раствора BaCl2 в 30%-ом глицерине. Оптическую плотность образовавшейся взвеси измеряли с помощью полевого фотоколориметра ПФ-2 при длине волны 360 нм. Концентрацию сульфата вычисляли по калибровочной кривой, построенной по стандартным растворам сульфата натрия.
Температуру измеряли максимальным ртутным термометром. Концентрацию кислорода измеряли методом Винклера. Значения pH и Eh определяли при помощи полевого иономера И-102.
Микроскопия. Морфологию клеток в природных образцах и в выделенных культурах изучали в световом микроскопе с фазовым контрастом Olympus BX 41 (“Olympus Corp.”, Япония).
Определение пигментов. Пробы для определения пигментов и для опытов с радиоизотопами отбирали из мест формирования видимых обрастаний, характерных для микробных матов, с помощью пробочного сверла площадью 1 см2.
Фиксацию проб для определения состава пигментов проводили с помощью 50%-го глицерина. Пробы хранили в темноте при комнатной температуре, а затем в холодильнике.
Пигменты микробных матов определяли из проб, фиксированных глицерином (1 : 1). В лаборатории пробы обрабатывали 2 мин ультразвуком (14.5 кГц) на приборе УЗД 1 и снимали развернутые спектры фрагментов мембран (так называемый спектр пигментов “in vivo”) на спектрофотометре Unicam-1800 в интервале длин волн 400–850 нм.
Из отдельно отобранной пробы проводили экстракцию пигментов ацетоном в течение 1 сут в темноте при 4°С, после чего снимали спектры поглощения растворенных пигментов на спектрофотометре. Для расчетов содержания хлорофиллов использовались следующие формулы:
Для измерения спектров поглощения пигментов выделенных культур АФБ были приготовлены тотальные препараты фрагментов клеточных мембран, полученные при разрушении клеток ультразвуком частотой 14.5 кГц с использованием Bandelin electronic UW 2070 (Германия). Кроме того, исследовали спектральные характеристики ацетон-метанольных (7 : 2) экстрактов клеток изолированных штаммов. Для анализа состава пигментов в клетках выделенных культур бактерий использовали спектрофотометр СФ 56А (“ЛОМО”, Россия). Спектры поглощения пигментов регистрировали в диапазоне длин волн 350–1100 нм.
Определение световой фиксации углекислоты. Для определения продукции оксигенного и аноксигенного фотосинтеза отобранные пробы матов с площади 1 см2 помещали в пенициллиновые флаконы объемом 20 мл, которые заполняли полностью водой источника, отобранной шприцем над поверхностью мата. Флаконы закрывали резиновыми пробками без пузырька воздуха. В качестве селективного ингибитора оксигенного фотосинтеза использовали диурон в конечной концентрации 10–7 мМ. В экспериментах в трех повторностях использовали 2 светлые (с диуроном и без диурона), 1 темную склянку и одну контрольную, фиксированную формалином (0.2 мл 40% формалина на флакон). Во все склянки шприцем добавляли по 0.2 мл (20 мкКи) раствора NaH14CO3. Склянки экспонировали в ложе источников в температурных и световых условиях in situ. По завершении экспозиции (4 ч) содержимое светлых и темных флаконов фиксировали 0.2 мл 40% формалина. На завершающем этапе фиксированные пробы матов подкисляли фосфорной кислотой, продували азотом для удаления меченой углекислоты, затем гомогенизировали ультразвуком, после чего 0.1 мл пробы помещали в сцинтилляционную жидкость ЖС-8. Таким образом, учитывалась как клеточная, так и внеклеточная продукция. Измерение радиоактивности продуктов проводили на сцинтилляционном счетчике Rack-Betta 1219 (“LKB”, Швеция).
Бактериальную продукцию рассчитывали по формуле Стимана–Нельсона (Кузнецов, Дубинина, 1989). Скорость оксигенного фотосинтеза вычисляли по разнице между суммарным фотосинтезом (без диурона) и аноксигенным фотосинтезом (с диуроном). В расчет брали среднеарифметическое значение данных из двух повторностей каждого варианта опыта:
где: мг С – суммарный фотосинтез, Ck – концентрация всех форм углерода, rсв – радиоактивность пробы из светлой склянки без диурона, rтемн – радиоактивность пробы из темной склянки, t – время экспозиции. Контрольная проба, убитая формалином, не фиксировала меченый гидрокарбонат и в расчет не бралась.Посевы и очистка культур. Посевы природных проб и выделенных культур проводили на универсальную среду Пфеннига (Pfennig, 1965), содержащую (г/л): NH4Cl – 0.33, КН2РO4 – 0.33, КСl – 0.33, MgCl2 · 6Н2O – 0.33, NaHCO3 – 0.5, Na2SO4 – 0.33, NaСl – 50 г/л. Среду готовили на дистилированной воде с добавлением раствора микроэлементов и витаминов (Pfennig, Lippert, 1966). После стерилизации в среду добавляли (г/л): Na2S · 9Н2О – 0.5, Na2S2O3 – 0.5, CH3COONa – 0.5 и агар (“Difco”) – 1.8%. Конечный рН среды был близок к 7.0.
Для определения относительной численности АФБ производили посев проб в пробирки с агаризованной (0.5% агара) питательной средой с соленостью 50 г/л методом предельных серийных разведений. В качестве посевного материала использовались пробы микробных матов объемом 1 мл, собранных шприцем с поверхности ложа источников. Перед посевом пробы разбавляли водой источника (1 : 1), затем гомогенизировали встряхиванием со стеклянными бусами. Культивирование проводили анаэробно при естественном освещении в полевых условиях. По приезде в Москву посевы доращивали в лабораторных условиях под лампами дневного света при освещенности 2000 люкс и температуре 20–25°С. Относительную численность пурпурных и зеленых бактерий определяли микроскопированием красных и зеленых колоний, выросших в последних разведениях. На этом этапе родовую принадлежность АФБ определяли по морфологическим признакам. Идентификацию выделенных чистых культур проводили методами молекулярной биологии.
Молекулярно-генетическая идентификация бактерий. ДНК изолированных фототрофных бактерий выделяли с помощью набора PowerMax Soil DNA Isolation Kit (“MO BIO Laboratories, Inc.”, США) согласно инструкции производителя.
Фрагменты участка оперона pufLM амплифицировали и секвенировали с использованием праймерных систем, специфичных для аноксигенных фототрофных пурпурных бактерий (Béjà et al., 2002). Для зеленой серобактерии использовали специфичный для зеленых серобактерий праймер F-99-GSB и бактериальный R-1369 (Alexander et al., 2002).
Продукты амплификации очищали с использованием набора Wizard SV Gel and PCR Clean-Up System (“Promega”, США), согласно инструкции производителя. Секвенирование продуктов амплификации проводили по методу Сэнгера с использованием набора реактивов BigDye Terminator v3.1 Cycle Sequencing Kit на автоматическом секвенаторе DNA Analyzer 3730 (“Applied Biosystems”, США), согласно инструкции производителя.
Редактирование полученных последовательностей проводили в программе BioEdit [http://jwbrown.mbio.ncsu.edu/BioEdit/bioedit.html]. Сравнительный анализ с последовательностями базы данных GenBank проводили с помощью программы BLAST [http://www.ncbi.nlm.nih.gov/blast]. Полученные de novo нуклеотидные последовательности были проверены на наличие химер с помощью программы DECIPHER (Wright et al., 2012). Дендрограммы были построены по методу Maximum Likelihood с помощью программного пакета MEGA 6.1 (Tamura et al., 2013).
Полученные нуклеотидные последовательности гена pufLM были депонированы в базу данных GenBank под номерами: MH800851–MH800854. Данные о нуклеотидном составе гена 16S рРНК Ectothiorhodospira sp. Dg9-1 предоставлены Й. Имхоффом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИE
Основные группы фототрофных бактерий в составе микробных матов. Температура источников на выходе составляла 57–60°С, однако обрастания фототрофов наблюдались только при температурах ниже 54°С. Основными матобразующими микроорганизмами были нитчатые цианобактерии. Предварительную диагностику цианобактерий проводили на основании их морфологических особенностей. Количество видов цианобактерий в микробных матах первой подгруппы гидротерм было ограничено, очевидно, из-за высокой солености изливающегося рассола. Обнаружены два вида Phormidium и один вид морфологически сходных с Synechosystis одноклеточных сине-зеленых водорослей (рис. 1).
Рис. 1.
Морфотипы фототрофных бактерий в природных образцах: а – Phormidium sp., б – Leptolyngbya sp. (Lep.), Chloroflexus-подобные микроорганизмы (Cfl.), Spirulina sp. (Spir.), в – Phormidium sp., г – Chloroflexus-подобные микроорганизмы, д – Ectothiorhodospira sp., е – Synechocystis sp., ж – Phormidium sp. S – включение элементарной серы. Масштабная метка – 10 мкм.
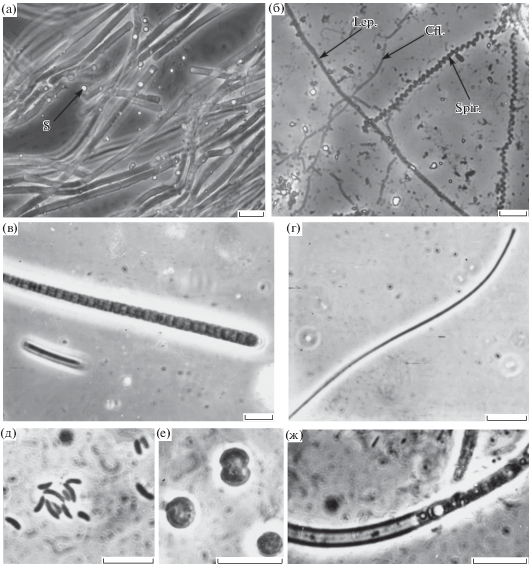
Посев на элективные среды показал, что мезотермальные условия, повышенная соленость, присутствие сульфида и нейтральный рН обусловливали развитие в матах Берикейских источников морских и умеренно галофильных АФБ, участвующих в окислении сульфида: зеленых серобактерий (ЗСБ) Prosthecochloris sp. и пурпурных серобактерий (ПСБ) рода Ectothiorhodospira sp., Chromatium-подобных бактерий, а также несерных пурпурных бактерий (НПБ) Rhodovulum sp. (табл. 1). Кроме того, в выросших культурах и при прямом микрокопировании природного материала обнаружены бактерии, морфологически сходные с зелеными аноксигенными нитчатыми фототрофными бактериями (АНФБ) рода Chloroflexus (рис. 1б). Численность пурпурных и зеленых АФБ достигала около 104–105 клеток на мл (табл. 1).
Видовой состав сообщества фототрофов в источниках второй подгруппы был расширен вследствие понижения общей минерализации (табл. 1) по сравнению с источниками первой подгруппы. Разнообразие фототрофов расширялось, прежде всего, за счет дополнительных видов цианобактерий. При микроскопировании обнаружено 3 вида нитчатых цианобактерий родов Phormidium и Oscillatoria, Spirulina (рис. 1б). Кроме того, в мате встречались отдельные нити бесцветных серобактерий Beggiatoa sp.
Видовой состав АФБ в альгобактериальных матах был такой же, как и в источниках первой подгруппы. В обрастании встречались ПСБ Ectothiorhodospira sp., морфотип Chromatium sp., НПБ Rhodovulum sp., ЗСБ Prosthecochloris sp. и АНФБ Chloroflexus sp. (табл. 1). Численность пурпурных и зеленых АФБ в источниках второй подгруппы была выше, чем в первой подгруппе, до 107 клеток в 1 мл (табл. 1). Ограничение развития фототрофов высокой температурой и соленостью было обнаружено и здесь. Формирование мата наблюдалось при температуре ниже 54°C.
Физико-химические условия в местах формирования микробных матов. В точках 7 и 9 при низком значении окислительно-восстановительного потенциала воды, равном –85 мВ, значение Eh на поверхности мата было положительным и составляло, соответственно, +5 и +40 мВ. Обнаруженные матобразующие цианобактерии содержали фикобилиновые пигменты и осуществляли оксигенный фотосинтез в присутствии сульфида. Содержание кислорода в верхней части мата варьировало от 0.4 до 1.8 мг/л.
Для системы Берикейских источников в целом был характерен высокий уровень первичной продукции микробных матов, до 53.8 мкг С/(см2 ч). За сутки это составляло 4.3 г С/(м2 сут). Эта величина хорошо коррелирует с цифрами, приводимыми для сходных систем Крумбейном – 1–5 г С/(м2 сут) (Krumbein et al., 1977). На выходе источников концентрация сероводорода повышена, и значительная часть органического вещества (около 20%) образовывалась за счет аноксигенного фотосинтеза. Особо отметим, что в Берикейских источниках преобладали виды цианобактерий, не способные к аноксигенному фотосинтезу. Тем не менее, при микроскопировании видно, что элементная сера откладывается на поверхности трихомов цианобактерий (рис. 1а). Окисление сероводорода, видимо, происходит химическим путем за счет кислорода, продуцированного при фотосинтезе цианобактериями (Castenholz, 1973).
Характеристика выделенных культур аноксигенных фототрофных бактерий. Из типичных для Берикейских источников микробных матов были выделены: зеленая серобактерия – штамм Dg19, пурпурные серобактерии – штаммы Dg9-1 и Dg9-2, и несерная пурпурная бактерия – штамм Dg9-4.
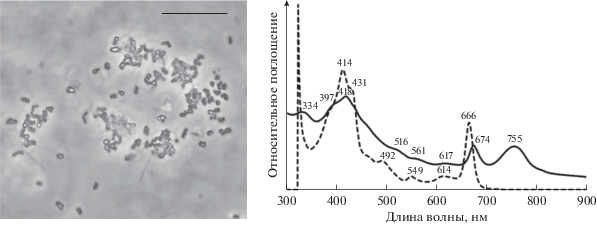
Штамм Dg19. Морфология клеток бактерий штамма DG19 типична для бактерий рода Prosthecochloris (рис. 2). Округлые клетки размером около 1 мкм имели короткие выросты – простеки, газовые вакуоли отсутствовали. Капли серы откладывались вне клеток в процессе фототрофного окисления сульфида. Суспензия клеток молодой культуры Dg19 имела ярко-зеленый цвет. Спектр поглощения in vivo показывал максимумы 334, (397), 418, 516, 561, 617, 674, 755 нм. Спектр поглощения ацетон-метанольного экстракта клеток бактерии показывал максимумы при (397), 414, (431), 492, 549, 614, 666 нм. Преобладающим пигментом у штамма Dg19 являлся бактериохлорофилл c, имеющий на спектрах in vivo основной пик в целых клетках в длинноволновой области 755 нм, а в ацетон-метанольном экстракте – при 666 нм (рис. 2).
Рис. 2.
Морфология и спектры поглощения целых клеток (сплошная линия) и ацетон-метанолового экстракта (пунктирная линия) зеленой серной бактерии штамма Prosthechochloris sp. Dg19. Масштабная метка – 10 мкм.
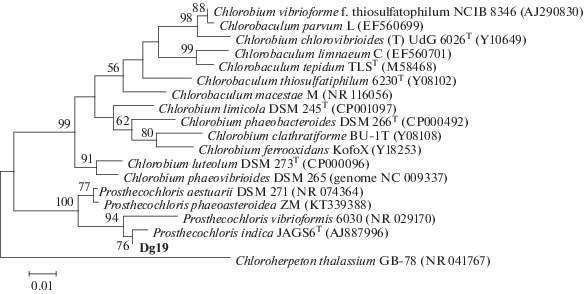
Таксономическая принадлежность штамма Dg19 была определена из накопительной культуры, не содержащей других АФБ. Бактерия относится к виду Prosthecochloris indica (этот вид не валидирован), с которым имеет 99% сходства (рис. 3).
Рис. 3.
Дендрограмма филогенетического сходства последовательностей генов 16S рРНК нового штамма Dg19 и изученных представителей ЗСБ. Дендрограмма построена с использованием алгоритма максимального подобия (Maximum Likelihood) с применением модели Тамуры–Нея. Достоверность ветвления рассчитана по результатам анализа 500 альтернативных дендрограмм. Длина сравниваемых последовательностей составляла 1192 пар нуклеотидов.
Численность ЗСБ в Берикейских источниках возрастала при относительно низких соленостях. Все известные виды бактерий рода Prosthecochloris являются умеренными галофилами с пределом развития обычно около 60 г/л (Gorlenko, 1988).
Среди АФБ, особенно во второй подгруппе источников, доминировали ПСБ рода Ectothiorhodospira, для которых характерно отложение серы вне клеток. Было выделено два фенотипически близких вида бактерий Dg-16 (Kuntikov, Gorlenko, 1998) и Dg9-1.
Штамм Dg9-1. Клетки штамма Dg9-1 представляли собой слабо извитые спириллы размером 1.2 × 3 мкм (рис. 4). Бактерии использовали на свету сульфид или тиосульфат, капли серы откладывали вне клеток. Культура штамма Dg9-1 имела красно-коричневый цвет.
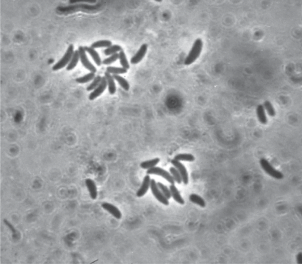
Спектр поглощения живых клеток (in vivo) штамма Dg9-1 свидетельствовал о присутствии пигментов фотосинтеза бхл a с максимумами при 378, 591, 797, 860, (879) нм и каротиноидов спириллоксантиновой серии – 398, 493, 527 нм.
Штамм Dg9-1 был выделен в чистую культуру и идентифицирован нами как новый штамм вида Ectothiorhodospira marismortui с уровнем сходства последовательностей гена 16S рРНК 97%. Этот вид выделен из соленого серного источника на берегу озера Солар Лейк (Синайский полуостров) (Oren et al., 1989). В результате детальных молекулярно-генетических исследований Ectothiorhodospira marismortui был признан штаммом ранее описанного Пельшем вида Ectothiorhodospira mobilis (Пельш, 1937; Imhoff, Süling, 1996).
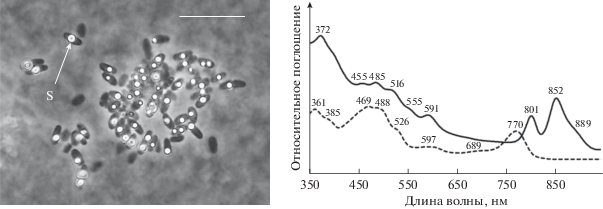
Штамм Dg9-2. В менее соленых матах первой и второй подгрупп источников обнаружены пурпурные бактерии семейства Chromatiaceae. В матах источника первой подгруппы при солености 97 г/л была изолирована и идентифицирована пурпурная серобактерия Marichromatium gracile (рис. 5). ПСБ рода Marichromatium могут расти при солености до 130 г/л и температуре до 35°C (Imhoff, 2001). Бактерия Marichromatium Dg9-2 имела клетки размером 1.2 × 2.5 мкм, окисляла сульфид, откладывая серу внутриклеточно (рис. 6). Спектр поглощения живых клеток (in vivo) штамма Dg9-2 свидетельствовал о присутствии пигментов фотосинтеза бхл a с максимумами при 372, 591, 801, 852, (889) нм и каротиноидов серии спириллоксантина с максимумами 455, 485, 516, (555) нм. Спектр поглощения ацетон-метанольного экстракта клеток бактерии показывал максимумы поглощения при 361, (385), 469, 488, 526, 597, 689, 770 нм.
Рис. 5.
Дендрограмма филогенетического сходства последовательностей генов PufLM новых штаммов Marichromatium sp. Dg9-2 и Rhodovulum sp. Dg9-3 с другими представителями пурпурных бактерий. Достоверность ветвления рассчитана по результатам анализа 500 альтернативных дендрограмм. Длина сравниваемых последовательностей составляла 430 аминокислот.
Рис. 6.
Морфология и спектры поглощения целых клеток (сплошная линия) и ацетон-метанолового экстракта (пунктирная линия) пурпурной серной бактерии штамма Marichromatium sp. Dg9-2. S – включение элементарной серы. Масштабная метка – 10 мкм.
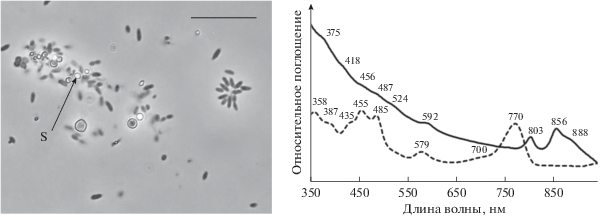
Штамм Dg9-3. В микробных матах мезотермальных Берикейских источников в значительном количестве присутствовали несерные пурпурные бактерии, идентифицированные как Rhodovulum Dg9-3 (рис. 5). По данным анализа гена, кодирующего фотосинтезирующий комплекс pufLM, эта бактерия близка к видам Rhodovulum visakhapatnamense и R. sulfidophilum. Rhodovulum Dg9-3, а по физиологии – к Rhodovulum sulfidophilum. Клетки бактерии – короткие палочки (1.2 × × 1.8 мкм), иногда образующие характерные звездообразные микроколонии (рис. 7). Основной пигмент – бактериохлорофилл а, а также каротиноид сфероиден (альтернативная спириллоксантиновая серия). Спектр поглощения живых клеток (in vivo) штамма Dg9-3 свидетельствовал о присутствии пигментов фотосинтеза бхл a с максимумами при (375), 418, 592, 803, 856, 888 нм и каротиноидов с максимумами (456), 487, 524 нм. Спектр поглощения ацетон-метанольного экстракта клеток бактерии имел максимумы поглощения при 358, (387), (435), 455, 485, 579, 700, 770 нм. Бактерия являлась умеренным галофилом, в процессе фотосинтеза окисляла сульфид, откладывая серу вне клеток. По этому свойству выделенные несерные пурпурные бактерии конкурируют с пурпурными серобактериями. В наших экспериментах они хорошо росли при концентрации сульфида 500 мг/л. В то же время, бактерии рода Rhodovulum способны к фотогетеротрофному росту, используя широкий спектр органических соединений (Imhoff, 2001).
Рис. 7.
Морфология и спектры поглощения целых клеток (сплошная линия) и ацетон-метанолового экстракта (пунктирная линия) пурпурной несерной бактерии штамма Rhodovulum sp. Dg9-3. S – включение элементарной серы. Масштабная метка – 10 мкм.
Следует отметить присутствие в микробных матах АНФБ, которые не удалось идентифицировать до вида. Однако сам факт нахождения АНФБ при высоких соленостях до 95 г/л предполагает его принадлежность к известному экстремальному (в отношении солености) роду Chlorothrix (Klappenbach, Pierson, 2004).
В заключение можно отметить, что в пресных гидротермах граница распространения цианобактериального мата находится около 70°С. Это предельная температура нахождения как оксигенных, так и аноксигенных фототрофов. В высокоминерализованных термальных источниках, как нейтральных, так и щелочных, развития истинно термофильных ФБ не происходит, а появление фототрофного мата начинается ниже 47–54°С. Примерами служат исследованные нами нейтральные Берикейские источники (Дагестан, Северный Кавказ), гиперсоленые гидротермы на побережье Мертвого моря (Израиль), теплое озеро Вашингтон (США) и щелочные соленые источники на острове Паоха в центре содового озера Моно Лейк (США) (Oren et al., 1989; Горленко, 2007; Thiel et al., 2017). В микробных матах этих соленоводных гидротерм не обнаружено термофильных фототрофных сообществ.
Однако некоторые соленоводные АФБ проявляют толерантность к повышенным температурам. В литературе описаны случаи развития Prosthecochloris sp. в соленом озере Солар Лейк, вода которого прогревается до 60°C (Cohen et al., 1977). Выделенный из теплого соленого озера Вашингтон штамм Prosthecochloris sp. имеет пределы развития до 45°С (Thiel et al., 2017). Известны умеренно термофильные формы Halorhodospira, обитающие в гиперсоленых и содовых водоемах и имеющие температурный оптимум 37–42°С (Imhoff, Truper, 1977; Imhoff, 2001). Приведенные примеры не противоречат парадигме, что среди соленоводных АФБ нет истинно термофильных микроорганизмов.
В гиперсоленых Берикейских источниках обнаружены галофильные формы АФБ: зеленые бактерии рода Prosthecochloris, пурпурные серобактерии Ectothiorhodospira sp., Marichromatium sp., Rhodovulum sp. и неизвестные мезофильные Chloroflexi. Примечательно, что эти виды находились в функционально активном состоянии в микробном мате при повышенных температурах до 54°С.
Соленость среды влияет на температурный оптимум развития АФБ (Kuntikov, Gorlenko, 1998). Ранее было установлено на примере штаммов Prosthecochloris sp. Dg19 и Ectothiorhodospira sp. Dg16, что при более высокой солености температурный оптимум АФБ смещается в более высокую область. Так, ЗСБ при солености 20 г/л лучше развивались при температуре 30°C, а при солености 80 г/л штамм Dg19 развивался слабо, оптимум развития имел при 35°C. Штамм Ectothiorhodospira sp. Dg16 хорошо развивался в широком интервале солености (от 20 до 120 г/л), при солености 80 г/л имел температурный оптимум 40–42°C. Нами показано, что штамм Ectothiorhodospira sp. Dg9-1 обладал теми же физиологическими характеристиками, что и штамм Dg16.
Таким образом, исследованные галофильные штаммы ЗСБ и преобладающие виды ПСБ являются галотолерантами и приспособлены к жизни в гиперсоленых Берикейских источниках в мезотермальных условиях с нейтральным рН.
Авторы приносят благодарность сотрудникам и студентам, принимавшим участие в полевых работах по изучению микробных матов Берийских источников.
Работа выполнена при финансовой поддержке программы Президиума РАН “Эволюция органического мира и планетарных процессов” (подпрограмма 2), РФФИ (проекты № 16-04-00830 и 19-04-00423) и Министерства науки и высшего образования РФ. Новые фототрофные бактерии описаны в рамках госзадания № 0104-2018-0033. Секвенирование ДНК частично проводили с использованием оборудования ЦКП “Биоинженерия” ФИЦ Биотехнологии РАН.
Список литературы
Горленко В.М. Аноксигенные фототрофные бактерии содовых озер // Труды Института микробиологии имени С.Н. Виноградского. Вып. XIV. Алкалофильные микробные сообщества / Под. ред. Гальченко В.Ф. М.: Наука, 2007. С. 225–257.
Дейнега Г.И. Зависимость динамики и химизма подземных вод от сейсмической активности недр. Труды Института геологии, Дагестан. Филиал АН СССР, 1978. С. 10.
Курбанов М.К. Геотермальные и гидроминеральные ресурсы Восточного Кавказа и Предкавказья. М.: Наука, 2001. 260 с.
Кузнецов С.И., Дубинина Г.А. Методы изучения водных микроорганизмов. М.: Наука, 1989. 285 с.
Пельш А.Д. О фотосинтезирующих серобактериях восточного бассейна Сакского озера // Микробиология. 1937. Т. 6. № 8. С. 1092–1100.
Резников А.А., Муликовская Е.П, Соколов И.Ю. Методы анализа природных вод. М.: Недра, 1970. 488 с.
Старынин Д.А., Горленко В.М., Компанцева Е.И., Качалкин В.И., Чеботарев Е.Н. Особенности круговорота углерода и серы в высокосульфатном источнике Талги-W Дагестанской АССР // Известия АН СССР. Сер. Биол. 1990. № 5. С. 749–758.
Федоров В.Д. О методах изучения фитопланктона и его активности. М.: МГУ, 1979. 168 с.
Alexander B., Andersen J., Cox R., Imhoff J. Phylogeny of green sulfur bacteria on the basis of gene sequences of 16S rRNA and of the Fenna–Matthews–Olson protein // Arch. Microbiol. 2002. V. 178. P. 131–140.
Béjà O., Suzuki M.T., Heidelberg J.F., Nelson W.C., Preston C.M., Hamada T., Eisen J.A., Fraser C.M., De Long E.F. Unsuspected diversity among marine aerobic anoxygenic phototrophs // Nature. 2002. V. 415. № 6872. P. 630–633.
Castenholz R.W. The possible photosynthetic use of sulfide by the filamentous phototrophic bacteria of hot springs // Limnol. Oceanogr. 1973. V. 18. P. 863–876.
Cohen Y., Krumbein W.E., Shilo M. Solar Lake (Sinai). 2. Distribution of photosynthetic microorganisms and primary production1 // Limnol. Oceanogr. 1977. V. 22. P. 609–620.
Gorlenko V.M. Ecological niches of green sulfur and gliding bacteria // Green Photosynthetic Bacteria / Eds. Olson J.M., Ormerod J.G., Amesz J., et al. New York: Plenum Press, 1988. P. 257–268.
Imhoff J.F. True marine and halophilic anoxygenic phototrophic bacteria // Arch. Microbiol. 2001. V. 176. P. 243–254.
Imhoff J.F., Süling J. The phylogenetic relationship among Ectothiorhodospiraceae: a reevaluation of their taxonomy on the basis of 16S rDNA analyses // Arch. Microbiol. 2001. V. 165. P. 106–113.
Imhoff J.F., Truper H.G. Ectothiorhodospira halochloris sp. nov., a new extremely halophilic phototrophic bacterium containing bacteriochlorophyll b // Arch. Microbiol. 1977. V. 114. P. 115–121.
Klappenbach J.A., Pierson B.K. Phylogenetic and physiological characterization of a filamentous anoxygenic photoautotrophic bacterium “Candidatus Chlorothrix halophila” gen. nov., sp. nov., recovered from hypersaline microbial mats // Arch. Microbiol. 2004. V. 181. P. 17–25.
Krumbein W.E., Cohen Y., Shilo M. Solar Lake (Sinai). 4. Stromatolitic cyanobacterial mats // Limnol. Oceanogr. 1977. V. 22. P. 635–656.
Kuntikov E.I., Gorlenko V.M. Interrelation between halo- and thermotolerance in anoxygenic phototrophic bacteria // Microbiology. 1998. V. 67. № 3. P. 245–250.
Oren A., Kessel M., Stackebrandt E. Ectothiorhodospira marismortui sp. nov., an obligately anaerobic, moderately halophilic purple sulfur bacterium from a hypersaline sulfur spring on the shore of the Dead Sea // Arch. Microbiol. 1989. V. 151. P. 524–529.
Pfennig N. Anreicherungskulturen für rote und grüne Schwefelbakterien // Zb. Bakt., 1. Abt. Orig. Suppl. 1965. № 1. P. 503–504.
Pfennig N., Lippert K.D. Über das vitamin B12-bedürfnis phototropher Schwefelbakterien // Arch. Microbiol. 1966. V. 55. P. 245–256.
Takahashi M., Ichimura S. Vertical distribution and organic matter production of photosynthetic sulfur bacteria in Japanese lakes // Limnol. Oceanogr. 1968. V. 13. P. 644–655.
Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0 // Mol. Biol. Evol. 2013. V. 30. P. 2725–2729.
Thiel V., Drautz-Moses D.I., Purbojati R.W., Schuster S.C., Lindemann S., Bryant D.A. Genome sequence of Prosthecochloris sp. strain HL-130-GSB from the phylum Chlorobi // Genome Announc. 2017. V. 5. № 24. P. 1–2.
Wright E.S., Yilmaz L.S., Noguera D.R. DECIPHER, a search-based approach to chimera identification for 16S rRNA sequences // Appl. Environ. Microbiol. 2012. V. 78. P. 717–725.
Дополнительные материалы отсутствуют.