

Микробиология, 2022, T. 91, № 5, стр. 547-553
Галактофуранан клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ас-1098D
Н. В. Потехина a, *, А. С. Шашков b, Е. В. Арискина c, Н. В. Присяжная c, Е. М. Тульская a, Ф. М. Хасаева d, Л. И. Евтушенко c
a Московский государственный университет им. М.В. Ломоносова
119234 Москва, Россия
b Институт органической химии им. Н.Д. Зелинского РАН
119991 Москва, Россия
c ФИЦ Пущинский научный центр биологических исследований РАН,
Всероссийская коллекция микроорганизмов
142290 Пущино, Россия
d Кабардино-Балкарский государственный аграрный университет им. В.М. Кокова
360030 Нальчик, Кабардино-Балкарская Республика, Россия
* E-mail: potekhina56@mail.ru
Поступила в редакцию 25.04.2022
После доработки 17.05.2022
Принята к публикации 19.05.2022
- EDN: KMWBGQ
- DOI: 10.31857/S0026365622600365
Аннотация
Установлены структура гликополимера клеточной стенки и таксономическое положение штамма ВKM Ac-1098D, обладающего пиридин-деградирующей активностью. С использованием химических и ЯМР-спектроскопических методов гликополимер был идентифицирован как 1,6-связанный β-D-галактофуранан с боковыми остатками диаминоглюкозы (2,3-диацетамидо-2,3-дидезокси-β-глюкопиранозы, β-GlcpNAc3NAc). Структура полимера впервые описана у прокариот. Близкий по структуре галактофуранан, обнаруженный ранее у некоторых видов Paenarthrobacter, отличался от полимера штамма ВКМ Ас-1098D α-конфигурацией диаминоглюкозы (α-GlcpNAc3NAc) боковой цепи. Анализ последовательностей генов 16S рРНК и MALDI-TOF масс-спектров показал, что штамм ВKM Ac-1098D может быть отнесен к новому виду рода Paenarthrobacter, предварительно названному “Paenarthrobacter pyridinovorans”. Данные, полученные в этом исследовании и ранее, свидетельствуют о том, что галактофуранан с диаминоглюкозой в боковой цепи может служить хемотаксономическим признаком рода Paenarthrobacter, а α-конфигурация гликозидной связи диаминоглюкозы ‒ являться особенностью предполагаемого нового вида.
Клеточные стенки актинобактерий, как и других грамположительных бактерий, наряду с пептидогликаном обычно содержат ряд вторичных гликополимеров (Kohler et al., 2009). К ним относятся тейхоевые кислоты, поли(гликозил-1-фосфаты) и различные бесфосфатные полимеры, ковалентно связанные с пептидогликаном, а также липотейхоевые кислоты, связанные с мембранами (Naumova et al., 2001; Kohler et al., 2009; Потехина и соавт., 2011; Тульская и соавт., 2011). Типы и структурные вариации пептидогликана хорошо известны как важнейшие хемотаксономические признаки таксонов актинобактерий разного уровня (Schumann, 2011).
Вторичные гликополимеры бактерий в сравнении с пептидогликанами являются менее изученными в таксономическом аспекте. Однако имеющиеся в литературе сведения указывают на то, что состав, структуры и отдельные структурные компоненты гликополимеров могут быть также специфичными для видов, родов и высших таксонов актинобактерий (Naumova et al., 2001; Schumann et al., 2009; Evtushenko, Ariskina, 2015; Nouioui et al., 2018; Шашков и соавт., 2020; Потехина и соавт., 2021). В частности, галактофуранан с диаминоглюкозой в боковой цепи был обнаружен у штаммов всех трех изученных видов рода Paenarthrobacter – P. aurescens, P. histidinovorans и P. nicotinovorans (относившихся ранее к роду Arthrobacter) (Busse, 2016). Это указывает на то, что вышеупомянутый полимер может являться хемотаксономическим маркером рода Paenarthrobacter (Потехина и соавт., 2021).
В настоящей работе представлены результаты изучения гликополимеров клеточной стенки штамма ВКМ Ас-1098D, деструктора пиридина (Khasaeva et al., 2011), и приведены данные, с высокой степенью вероятности указывающие на принадлежность этого штамма к новому виду рода Paenarthrobacter. Полученные результаты согласуются с ранее высказанным предположением о специфичности галактофуранана с диаминоглюкозой в боковой цепи для бактерий рода Paenarthrobacter.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Используемый в работе штамм ВКМ Ас-1098D был получен из Всероссийской коллекции микроорганизмов (https://www.vkm.ru). Штамм был помещен в коллекцию под названием Arthrobacter sp. (Khasaeva et al., 2011) и впоследствии реидентифицирован как Paenarthrobacter sp. на основе короткого фрагмента (1200 п.н.) гена 16S рРНК.
Накопление биомассы, выделение клеточных стенок и препаратов гликополимеров, их гидролиз и анализ продуктов гидролиза методами электрофореза и хроматографии на бумаге проводили, как описано ранее (Potekhina et al., 2011).
Выделение ДНК, амплификацию, секвенирование и анализ последовательности гена 16S рРНК штамма ВКМ Ас-1098D проводили, как описано (Ryzhmanova et al., 2017). Процент сходства между последовательностями 16S рРНК гена рассчитывали с помощью программы TaxonDC версии 1.3.1. (Tarlachkov, Starodumova, 2017). MALDI-TOF масс-спектры целых клеток штаммов, выращенных на агаре R2A в течение 3 сут, регистрировали и анализировали, как описано ранее (Присяжная и соавт., 2012).
Спектры ЯМР 1Н и 13С снимали на спектрометре Bruker Avance II 600 (“Bruker”, Германия) для растворов в 99.96% D2O при температуре 323°К, обеспечивающей минимальное перекрытие сигнала остаточной воды с сигналами полимеров. В качестве калибровочного стандарта использовали TSP (натриевая соль 3-(триметилсилил)-2,2,3,3-тетрадейтеропропионовой кислоты) (δН 0.0, δС ‒1.6). Время смешивания в экспериментах 1Н, 1Н TOCSY и ROESY составляло 100 и 150 мс соответственно. Эксперимент 1H, 13C HM-BC был оптимизирован для констант спин-спинового взаимодействия JH, C 8 Гц.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сравнительный анализ почти полной последовательности гена 16S рРНК (1517 п.н.) штамма ВKM Ac-1098D подтвердил его принадлежность к роду Paenarthrobacter (рис. 1) и показал, что он наиболее близок филогенетически к P. nicotinovorans DSM 420 (99.6% сходства), P. ureafaciens DSM 20126 (99.3%) и P. histidinolovorans DSM 20115 (99.3%). Эти значения достаточно высоки, но сопоставимы или ниже значений между типовыми штаммами некоторых близких видов этого рода, например, P. aurescens и P. nitroguaiacolicus (99.9%), P. histidinovorans и P. nicotinovorans (99.7%) (табл. 1). Кроме того, штамм хорошо обособляется от всех вышеперечисленных видов по данным MALDI-TOF масс-спектрометрии (рис. 2) – метода с более высоким таксономическим разрешением, чем секвенирование 16S рРНК, и широко используемого для видовой идентификации бактерий (Mellmann et al., 2008; Seuylemezian et al., 2018). Максимальные показатели сходства, определенные с помощью программного обеспечения Bruker MALDI Biotyper между спектром штамма ВКМ Ас-1098D и спектрами типовых штаммов видов Paenarthrobacter (P. ureafaciens и P. aurescens), были ~1.75, что ниже значений, свойственных штаммам одного вида (≥2.0; Mellmann et al., 2008; рекомендации производителя). Таким образом, изученный штамм, по всей вероятности, является представителем нового вида, предварительно названного “Paenarthrobacter pyridinovorans”.
Рис. 1.
Филогенетическое дерево на основе генов 16S рРНК, построенное по методу максимального правдоподобия (Мaximum-likelihood), показывающее положение “Paenarthrobacter pyridinovorans” ВKM Ac-1098D среди видов рода Paenarthrobacter. Числа в точках ветвления указывают на значения “bootstrap” >50% (1000 повторов). В качестве внешней группы использовали штамм Microbacterium lacticum DSM20427T-интекс, как на картинке (X77441). Масштаб ‒ 0.02 замены на одно положение нуклеотидной последовательности.
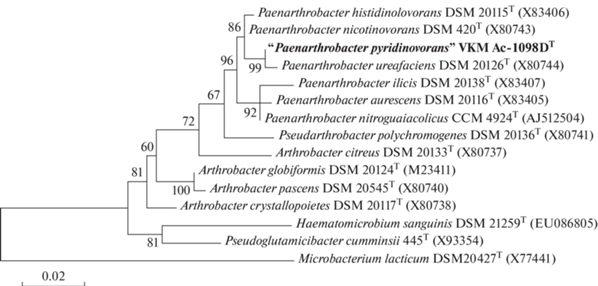
Таблица 1.
Химические сдвиги в 13С и 1H ЯМР-спектрах для галактофуранана из клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ac-1098D
| Остаток | Химические сдвиги (δC TSP ‒1.6 и δHTSP0.00) | |||||
|---|---|---|---|---|---|---|
| C-1 H-1 |
C-2 H-2 |
C-3 H-3 |
C-4 H-4 |
C-5 H-5 |
C-6 H-6,6' |
|
| →6)-β-D-Galf-(1→ (A') | 109.02 5.05 |
82.21 4.12 |
78.07 4.08 |
84.36 3.99 |
71.22 3.95 |
70.28 3.84, 3.62 |
| →6)-β-D-Galf-(1→ 2) (A) |
107.73 5.17 |
89.69 4.24 |
77.08 4.06 |
84.46 3.97 |
71.02 3.86 |
70.28 3.84, 3.60 |
| ↑ β-D-GlcpNAc3NAc-(1 (B) |
101.89 4.79 |
55.14а,б 3.73 |
56.23а,в 3.97 |
68.89 3.53 |
78.30 3.54 |
61.80 3.92, 3.77 |
Рис. 2.
Дендрограмма MALDI-TOF масс-спектров “Paenarthrobacter pyridinovorans” ВКМ Ac-1098D и типовых штаммов близких видов рода Paenarthrobacter.

Галактоза, глюкоза, аминосахара и следовые количества маннозы обнаружены в кислотном гидролизате клеточной стенки (нисходящая хроматография на бумаге), в то время как фосфатсодержащие соединения не были выявлены (электрофорез). Аналогичные моносахариды обнаружены и в гидролизатах препарата гликополимеров, выделенных из клеточной стенки ВКМ Ас-1098D.
Полная структура гликополимера клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ас-1098D установлена методами ЯМР-спектроскопии.
Спектр ЯМР 13С (рис. 3) содержал в области резонанса аномерных атомов углерода два основных сигнала при δС 101.89 и 107.73 м.д. и несколько минорных. В других характеристических областях спектра имелись два основных сигнала атомов углерода, связанных с азотом при δС 55.14 и 56.23 м.д., один сигнал углерода со свободной гидроксильной группой при δС 61.80 м.д. и один – с замещенной гидроксильной группой при δС 70.26 м.д., данные АРТ-спектра. В аномерной области спектра ЯМР 1Н имелись два основных сигнала – уширенный синглет при δН 5.17 м.д. и дублет с КССВ 8 Гц при δН 4.79 м.д.
Рис. 3.
Спектр 13C ЯМР препарата гликополимеров клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ac-1098D. Арабские цифры отнесены к атомам в остатках, обозначенных заглавными латинскими буквами как показано в таблице. Сигналы от разрушенного полимера обозначены звездочками.

Следует также отметить, что в спектре ЯМР 31Р препарата обнаружено небольшое количество гликозилфосфатных олигомеров, а в двумерных спектрах 1Н,13С HSQC и 1Н,31P HMBC присутствовали минорные сигналы в аномерной области ЯМР 13С и 1Н спектров. Однако структуру этого полимера, ввиду его малого содержания в препарате и почти полного разрушения при выделении, установить средствами ЯМР-спектроскопии не представлялось возможным.
Сигналы преобладающего полимера были отнесены на основе анализа двумерных гомоядерных 1Н,1Н COSY, TOCSY, ROESY и гетероядерных 1Н,13С HSQC и HMBC экспериментов.
Гомоядерные спектры выявили наличие в полимере остатков β-галактофуранозы (β-Galf, A, Aꞌ) и 2,3-диацетамидо-2,3-дидезокси-β-глюкопиранозы (β-GlcpNAc3NAc, B).
Анализ спектра 1Н,13С HSQC (рис. 4) показал, что большая часть остатков β-Galf (примерно 80%) замещена в положения 2 и 6, меньшая часть – только в положение 6. Другие остатки были не замещены по гидроксилам у С-4 (δС 68.89 м.д.) или С-6 (δС 61.80 м.д.) и, следовательно, являлись терминальными.
Рис. 4.
Части двумерного 1H,13C HSQC спектра галактофуранана клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ac-1098D. Соответствующие части 1H и 13C ЯМР-спектров приведены вверху и слева от двумерного спектра соответственно. Арабские цифры относятся к номерам атомов углерода в остатках, обозначенных заглавными латинскими буквами в соответствии с таблицей.

Последовательность соединения остатков следует из анализа спектра 1Н,13С HMBC (рис. 5). Наличие в спектре корреляционных пиков H-1 (A)/C-6(A, Aꞌ), (5.17/70.28, 70.28); H-1(Aꞌ)/C-6(A, Aꞌ), (5.05/70.28, 70.28); H-6(A)/C-1(A), (3.84 или 3.60/107.73) и H-6(Aꞌ)/C-1(Aꞌ), (3.84 или 3.62/109.02) свидетельствует о наличии полимера, основная цепь которого построена из остатков β-(1→6)-связанных остатков галактофуранозы.
Рис. 5.
Часть двумерного 1H,13C HMBC спектра галактофуранана клеточной стенки “Paenarthrobacter pyridinovorans” ВКМ Ac-1098D. Соответствующие части 1H и 13C ЯМР-спектров приведены вверху и слева от двумерного спектра. Арабские цифры до косой черты относятся к атомам протона, а после – к атомам углерода в остатках, обозначенных заглавными латинскими буквами в соответствии с таблицей.

Корреляционные пики H-1(B)/C-2(A), (4.79/89.69) и H-2(A)/C-1(B), (4.24/101.89) указывают на то, что большая часть остатков основной цени замещена остатками β-GlcpNAc3NAc в положении 2.
Таким образом, структуру основного полимера препарата можно представить формулой:
Таким образом, в результате проведенных исследований получены новые сведения о гликополимерах клеточных стенок бактерий. Как и другие виды рода Paenarthrobacter – P. aurescens, P. histidinovorans и P. nicotinovorans (Потехина и соавт., 2021), штамм “Paenarthrobacter pyridinovorans” ВКМ Ас-1098D характеризуется наличием в клеточной стенке галактофуранана с боковыми остатками диаминоглюкозы и имеет аналогичный набор cахаров (галактоза, глюкоза, аминосахар). Однако галактофуранан штамма ВКМ Ас-1098D с β-конфигурацией гликозидной связи остатка диаминоглюкозы (β-GlcpNAc3NAc) отличается от ранее описанного полисахарида Paenarthrobacter spp., который имеет α-конфигурацию диаминоглюкозы (α-GlcpNAc3NAc). Структуры галактофурананов, обнаруженных у штамма ВКМ Ас-1098D и трех видов Paenarthrobacter, не были описаны у других прокариот (http://csdb.glycoscience.ru/). Полученные в настоящей работе и ранее опубликованные данные свидетельствуют о том, что галактофуранан с боковыми остатками диаминоглюкозы может служить хемотаксономическим маркером рода Paenarthrobacter, а конфигурация гликозидной связи может являться особенностью предполагаемого нового вида.
Список литературы
Потехина Н.В., Шашков А.С., Тульская Е.М, Арискина Е.В., Дорофеева Л.В., Евтушенко Л.И. Галактофуранан клеточной стенки актинобактерий рода Paenarthrobacter // Микробиология. 2021. Т. 90. С. 122–128.
Potekhina N.V., Shashkov A.S., Ariskina E.V., Tul’skaya E.M., Dorofeeva L.V., Evtushenko L.I. Cell wall galactofuranan of the Paenarthrobacter actinobacteria // Microbiology (Moscow). 2021. V. 90. P. 106‒111. https://doi.org/10.1134/S0026261720060156
Присяжная Н.В., Плотникова Е.Г., Буева О.В., Корсакова Е.С., Дорофеева Л.В., Ильина Е.Н., Лебедев А.Т., Евтушенко Л.И. Применение MALDI-TOF масс-спектрометрии для разграничения близкородственных видов филогенетической группы “Arthrobacter crystallopoietes” // Микробиология. 2012. Т. 81. С. 696‒701.
Prisyazhnaya N.V., Plotnikova E.G., Bueva O.V., Korsakova E.S., Dorofeeva L.V., Il’ina E.N., Lebedev A.T., Evtushenko L.I. Application of MALDI-TOF mass spectrometry for differentiation of closely related species of the “Arthrobacter crystallopoietes” phylogenetic group // Microbiology (Moscow). 2012. V. 81. P. 754–759.
Шашков А.С., Тульская Е.М., Стрешинская Г.М., Дмитренок А.С., Потехина Н.В., Сенченкова С.Н., Пискункова Н.Ф., Дорофеева Л.В., Евтушенко Л.И. Рамноманнаны и тейхуроновая кислота из клеточной стенки Rathayibacter tritici ВКМ Ас-1603Т // Биохимия. 2020. Т. 85. С. 428–437.
Shashkov A.S., Tul’skaya E.M., Streshinskaya G.M., Dmitrenok A.S., Potekhina N.V., Senchenkova S.N., Piskunkova N.F., Dorofeeva L.V., Evtushenko L.I. Rhamnomannans and teichuronic acid from the cell wall of Rathayibacter tritici VKM Ac-1603T // Biochemistry (Moscow). 2020. V. 85. P. 369–377.
Тульская Е.М., Шашков А.С., Стрешинская Г.М., Сенченкова С.Н., Потехина Н.В., Козлова Ю.И., Евтушенко Л.И. Тейхуроновые и тейхулозоновые кислоты актиномицетов // Биохимия. 2011. Т. 76. С. 904–913.
Tul’skaya E.M., Shashkov A.S., Streshinskaya G.M., Senchenkova S.N., Potekhina N.V., Kozlova Yu.I., Evtushenko L.I. Teichuronic and teichulosonic acids of Actinomycetes // Biochemistry (Moscow). 2011. V. 76. P. 736–744.
Busse H.J. Review of the taxonomy of the genus Arthrobacter, emendation of the genus Arthrobacter sensu lato, proposal to reclassify selected species of the genus Arthrobacter in the novel genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov., Pseudoglutamicibacter gen. nov., Paenarthrobacter gen. nov. and Pseudarthrobacter gen. nov., and emended description of Arthrobacter roseus // Int. J. Syst. Evol. Microbiol. 2016. V. 66. P. 9–37.
Evtushenko L.I., Ariskina E.V. Nocardioidaceae // Bergey’s Manual of Systematics of Archaea and Bacteria / Ed. Whitman W.B. 2015. P. 1–18. https://doi.org/10.1002/9781118960608.fbm00042
Khasaeva F., Vasilyuk N., Terentyev P., Troshina M., Lebedev A.T. A novel soil bacterial strain degrading pyridines // Environ. Chem. Lett. 2011. V. 9. P. 439–445.
Kohler T., Xia G., Kulauzovic E., Peschel A. Teichoic acids, lipoteichoic acids, and related cell wall glycopolymers of Gram-positive bacteria // Microbial Glycobiology: Structures, Relevance and Applications / Eds. Moran A., Holst O., Brennan P., von Itzstein M. Amsterdam: Elsevier, 2009. P. 75–91.
Mellmann A., Cloud J., Maier T., Keckevoet U., Ramminger I., Iwen P., Dunn J., Hall G., Wilson D., Lasala P., Kostrzewa M., Harmsen D. Evaluation of matrix-assisted laser desorption ionization-time-of-flight mass spectrometry in comparison to 16S rRNA gene sequencing for species identification of non-fermenting bacteria // J. Clin. Microbiol. 2008. V. 46. P. 1946–1954. https://doi.org/10.1128/JCM.00157-08
Naumova I.B., Shashkov A.S., Tul’skaya E.M., Streshin-skaya G.M., Kozlova Yu.I., Potekhina N.V., Evtushenko L.I., Stackebrandt E. Cell wall teichoic acids: structural diversity, species-specificity in the genus Nocardiopsis, and chemotaxonomic perspective // FEMS Microbiol Rev. 2001. V. 25. P. 269–284.
Nouioui I., Carro L., Garcia-Lopez M., Meier-Kolthoff J.P., Woyke T., Kyrpides N.C., Pukall R., Klenk H.P., Goodfellow M., Goker M. Genome-based taxonomic classification of the phylum Actinobacteria // Front. Microbiol. 2018. V. 9. Art. 2007. https://doi.org/10.3389/fmicb.2018.02007
Potekhina N.V., Streshinskaya G.M., Tul’skaya E.M., Shashkov A.S. Cell wall teichoic acids in the taxonomy and characterization of Gram-positive bacteria // Taxonomy of Prokaryotes. Methods in Microbiology / Eds. Rainey F.A., Oren A. London: Academic Press, 2011. V. 38. Ch. 6. P. 132–164.
Ryzhmanova Y., Oshurkova V., Troshina O., Abashina T., Ariskina E., Avtukh A., Shcherbakova V. Anoxynatronum buryatiense sp. nov., an anaerobic alkaliphilic bacterium from a low mineralization soda lake in Buryatia, Russia // Int. J. Syst. Evol. Microbiol. 2017. V. 67. P. 4704–4709. https://doi.org/10.1099/ijsem.0.002365
Schumann P. Peptidoglycan structure // Taxonomy of Prokaryotes. Methods in Microbiology / Eds. Rainey F.A., Oren A. London: Academic Press, 2011. V. 38. Ch. 6. P. 101–129.
Schumann P., Kampfer P., Busse H.J., Evtushenko L.I. Proposed minimal standards for describing new genera and species of the suborder Micrococcineae // Int. J. Syst. Evol. Microbiol. 2009. V. 59. P. 1823–1849.
Seuylemezian A., Aronson H.S., Tan J., Lin M., Schubert W., Vaishampayan P. Development of a custom MALDI-TOF MS database for species-level identification of bacterial isolates collected from spacecraft and associated surfaces // Front. Microbiol. 2018. V. 9. Art. 780.
Tarlachkov S.V., Starodumova I.P. TaxonDC: calculating the similarity value of the 16S rRNA gene sequences of prokaryotes or ITS regions of fungi // J. Bioinform. Genom. 2017. V. 3(5). https://doi.org/10.18454/jbg.2017.3.5.1
Дополнительные материалы отсутствуют.