

Молекулярная биология, 2023, T. 57, № 2, стр. 197-208
Генотип-специфические особенности регуляции процесса холодового осахаривания клубней картофеля
А. А. Егорова a, b, *, И. А. Сабоиев b, Н. Е. Костина a, Д. Д. Куваева a, А. Б. Щербань a, b, С. М. Ибрагимова a, b, Е. А. Салина b, А. В. Кочетов a
a Федеральный исследовательский центр Институт цитологии и генетики Сибирского отделения
Российской академии наук
630090 Новосибирск, Россия
b Курчатовский геномный центр Институт цитологии и генетики Сибирского отделения Российской академии наук
630090 Новосибирск, Россия
* E-mail: egorova@bionet.nsc.ru
Поступила в редакцию 24.06.2022
После доработки 26.08.2022
Принята к публикации 26.08.2022
- EDN: EFXYKD
- DOI: 10.31857/S0026898423020064
Аннотация
Проанализирована динамика накопления сахаров, а также экспрессия генов вакуолярной инвертазы (Pain-1), сахарозосинтазы (SUS4), ингибитора инвертазы (InvInh2), регулирующих процесс холодового осахаривания, в клубнях картофеля (сорта Никулинский, Симфония, Невский) при хранении в условиях низких температур. Показано, что все три сорта накапливают сахара при низких температурах; при этом максимальное накопление редуцирующих сахаров наблюдается при температуре +4°С. Паттерн экспрессии генов, связанных с холодовым осахариванием, зависит от сорта и длительности хранения. На ранних сроках хранения уровень экспрессии гена вакуолярной инвертазы обратно пропорционален уровню экспрессии ее ингибитора. В клубнях голландского сорта Симфония выявлена высокая экспрессия гена вакуолярной инвертазы и низкая экспрессия гена ингибитора вакуолярной инвертазы; обратная зависимость наблюдается у российских сортов Никулинский и Невский. У всех трех сортов картофеля экспрессия гена сахарозосинтазы наиболее заметно повышена на поздних сроках хранения клубней. Обсуждается участие изученных генов в процессе холодового осахаривания в клубнях картофеля.
ВВЕДЕНИЕ
Картофель (Solanum tuberosum) – одна из важнейших продовольственных, кормовых, технических культур. Так как в зонах с умеренным климатом урожай картофеля убирают раз в году, осенью, то для круглогодичного использования клубни картофеля хранят при низких температурах, что предотвращает прорастание клубней, их усыхание и гниение. Срок хранения клубней в таких условиях не превышает 6 мес. [1]. Однако при хранении на холоде в клубнях картофеля накапливаются редуцирующие сахара, восстановительный потенциал которых обусловлен наличием свободных альдегидной или кетонной групп. Это накопление сопровождается появлением нежелательного сладкого вкуса (cold-induced sweetening, CIS, холодовое осахаривание). К тому же товарные характеристики клубней заметно ухудшаются, поскольку при термической обработке (например, при приготовлении картофельных чипсов) эти сахара реагируют со свободными аминокислотами в реакции Майяра, что приводит к появлению темноокрашенных горьких продуктов [2]. Также образуется акриламид, который считается токсичным и потенциально канцерогенным, хотя его прямая связь с развитием онкологических заболеваний не показана [3].
Накопление сахаров как веществ, обладающих осмопротекторными свойствами, в ответ на воздействие низких температур, видимо, является защитной метаболической реакцией, необходимой для выживания диких предков культурного картофеля, которые происходят из Южной Америки. Однако для современных культурных видов это нежелательный признак, так как из-за высокого круглогодичного спроса на клубни около 70% осеннего урожая картофеля требуется помещать на средне- или долгосрочное хранение [4]. Поэтому предотвращение процесса холодового осахаривания клубней картофеля важно для пищевой промышленности.
В клубнях картофеля сахара представлены в основном сахарозой, глюкозой и фруктозой, которые образуются в результате биохимического разложения крахмала. Эти сахара метаболизируются в процессе дыхания клубней, а также могут участвовать в реакции Майяра во время термической обработки. Хотя сахароза не участвует напрямую в реакции Майяра, она гидролизуется в процессе жарки, что способствует развитию окраски чипсов [5].
В клубнях картофеля, хранящихся в холодных условиях, запускается распад крахмала в амилопластах обусловленный индукцией амилолитических ферментов, а продукты распада выводятся из амилопласта либо в виде гексозофосфатов (гексоза-Р) через переносчик глюкозофосфата, либо в виде свободных сахаров транспортерами глюкозы и/или мальтозы. В цитозоле под действием сахарозофосфатсинтазы и сахарозофосфатазы эти метаболиты превращаются в сахарозу. Затем сахароза гидролизуется на составляющие ее гексозы вакуолярной инвертазой [6].
Функциональное значение и точный механизм накопления редуцирующих сахаров при хранении клубней картофеля на холоде до конца не изучены. В настоящее время процесс холодового осахаривания считается одной из самых серьезных проблем в производстве и переработке картофеля. При этом получение картофеля, устойчивого к условиям холодового осахаривания, становится приоритетным в ряде селекционных программ [7, 8].
Вакуолярная инвертаза (Pain-1, potato acid invertase 1; VacInv, beta-fructofuranosidase, [Е.С. 3.2.1.26]), локализованная в вакуолях, вносит основной вклад в холодовое осахаривание клубней картофеля. Показано, что экспрессия гена Pain-1 повышается при воздействии холода, а снижение экспрессии гена вакуолярной инвертазы методом РНК-интерференции [9, 10] и его нокаут методом TALEN [11] приводят к уменьшению накопления редуцирующих сахаров на холоде, вследствие чего полученные из мутантных растений чипсы имеют более светлый цвет, нежели чипсы из немутантных растений.
Идентифицирован ген вакуолярной инвертазы (Pain-1) картофеля, изучена его структура и экспрессия, а также найдены однонуклеотидные замены (SNP, single nucleotide polymorphism), предположительно влияющие на активность фермента [12, 13]. Показано, что активность промотора гена вакуолярной инвертазы регулируется сахарами, гормонами, холодом [14], а гиперметилирование промотора приводит к снижению экспрессии гена и накопления редуцирующих сахаров [15]. Регуляция инвертазы происходит не только на уровне транскрипции, но и посттрансляционно, с участием белков-ингибиторов. Найдены два ингибитора вакуолярной инвертазы, St-Inh (Inh1) и StInvInh2 (Inh2), локализованные, соответственно, в апопласте и вакуолях [16]. Показано, что активность вакуолярной инвертазы регулируется ингибиторами, но не влияет на уровень ее мРНК. Оба ингибитора играют роль в холодовом осахаривании, но показано, что Inh2 более эффективно ингибирует вакуолярную инвертазу [17, 18]. Уровень экспрессии Inh2 в клубнях картофеля с устойчивым к холодовому осахариванию генотипом также существенно выше, чем у сортов с чувствительным к осахариванию генотипом [16].
Сахарозосинтаза (SuSy, sucrose synthase, SUS, [E.C. 2.4.1.13]) катализирует превращение сахарозы и нуклеозиддифосфата (NDP) в нуклеозиддифосфатглюкозу (NDP-глюкоза) и фруктозу, а также обратный процесс синтеза сахарозы. Но в основном SUS расщепляет сахарозу и именно в этом качестве играет важную роль в метаболизме сахаров в клубнях картофеля. SUS4 – это изоформа сахарозосинтазы, которая чаще всего экспрессируется в клубнях [19]. Показана возможная связь сахарозосинтазы с холодовым осахариванием, а также повышение экспрессии этого фермента в ответ на воздействие холода [20–23]. Сахарозосинтаза, наряду с вакуолярной инвертазой и ее ингибиторами, вносит большой вклад в холодовое осахаривание [24]. Относительный вклад генов, кодирующих эти ферменты, и их регуляция в процессе холодового осахаривания остаются неясными. Вполне вероятно, что наряду с общими особенностями генетической регуляции холодового осахаривания существуют и генотипзависимые особенности, связанные с дифференцированным вкладом того или иного гена в этот процесс. Соответственно, для выработки оптимальной стратегии искусственного ингибирования процесса холодового осахаривания в селекционно-ценных сортах картофеля, необходимо детально исследовать динамику накопления сахаров в клубнях разных генотипов на холоде, параллельно оценивая уровень экспрессии, в первую очередь, структурных и метаболических генов, а также генов, вовлеченных в их регуляцию на разных уровнях.
Сорта российской селекции Никулинский, Невский, а также сорт голландской селекции Симфония популярны в России, имеют хорошие вкусовые качества, высокую урожайность, устойчивы ко многим заболеваниям и неблагоприятным факторам внешней среды (http://reestr.gossort.com). Но эти сорта генетически достаточно разнородны, что подтверждается их фенотипическими различиями – окраской цветков и клубней, массой клубня, содержанием крахмала, сроками созревания (http://reestr.gossort.com), опушением [25], спектральными характеристиками клубней [26]. Анализ маркеров этих сортов выявляет их генетическую разнородность по SSR-маркерам, маркерам устойчивости к заболеваниям [27, 28], SNP-маркерам, ассоциированным со свойствами крахмала [29, 30]. По набору хозяйственно-ценных признаков и уровню регенерационной способности in vitro эти сорта представляют интерес для работ по геномному редактированию [31]. Поэтому анализ процесса холодового осахаривания именно у этих сортов особенно актуален. В нашей работе проанализирована динамика накопления сахарозы, глюкозы и фруктозы в клубнях трех сортов картофеля при низких температурах, а также экспрессия ключевых генов-кандидатов (Pain-1, SUS4, Inh2), отвечающих за процесс холодового осахаривания.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Растительный материал. В исследовании использовали сорта картофеля Никулинский, Симфония и Невский СибНИИРС ИЦиГ СО РАН. Растения выращивали на поле СибНИИРС ИЦиГ СО РАН летом 2020 года в одинаковых условиях. Урожай убирали в конце августа и отбирали свежие здоровые клубни массой 100–130 г. Клубни просушивали на открытом воздухе в течение 7 дней при температуре 23°С, затем упаковывали в небольшие бумажные пакеты и хранили в холодильной камере (Pozis, линия Paracels) при фиксированной температуре 4, 8 и 12°С до 120 дней.
Анализ содержания крахмала. Содержание крахмала определяли в контрольной точке (0 дней, сразу после высушивания) в трех клубнях каждого сорта. Использовали метод, основанный на определении количества глюкозы, образовавшейся после ферментативного расщепления крахмала.
Пробу гомогенизировали, брали навеску 200 мг, добавляли 1 мл смеси ДМСО и соляной кислоты (16 мл ДМСО, 4 мл концентрированной HCl), инкубировали при температуре 60°С в течение 60 мин, тщательно перемешивая. Пробу переносили в стеклянные пробирки, добавляли 2.5 мл дистиллированной воды, смесь разбавляли в 5 раз и затем доводили pH до 4–5 с помощью гидроксида натрия (5 моль/л). Далее добавляли реактивы и инкубировали смесь согласно протоколу набора “Boehringer Ingelheim” (Германия).
Анализ содержания сахаров. В клубнях каждого сорта определяли содержание сахаров в контрольной точке (0 дней, сразу после высушивания), спустя 15, 30, 60, 90 и 120 дней хранения при указанных температурах. Брали по три клубня каждого сорта, промывали их и разрезали продольно пополам. Общее количество сахаров определяли с использованием антронового метода [32]. Содержание фруктозы и сахарозы определяли резорциновым методом [32], глюкозы – глюкозооксидазным методом, используя набор Глюкоза-Ново (“Вектор Бест”, Россия). Оптическую плотность раствора измеряли на сканирующем спектрофотометре СПЕК ССП-705. Содержание сахаров рассчитывали по калибровочным графикам. Средние значения, определенные по трем биологическим повторностям, обрабатывали статистически для оценки стандартного отклонения и представления экспериментальных данных в виде гистограмм.
Анализ экспрессии генов. Суммарную РНК выделяли из срединной части клубней (по три клубня каждого сорта) в контрольной точке (после высушивания), через 15 и 90 дней хранения при 4°C с использованием RNeasy Plant Mini Kit (“Qiagen”, Германия). Выделенную РНК обрабатывали ДНКазой (RNase-Free DNase Set, “Qiagen”). Концентрацию определяли с использованием NanoDrop 2000 (“Thermo Scientific”, США), качество РНК оценивали электрофоретически и с помощью Qubit 3.0 Fluorometer (“Thermo Scientific”). кДНК синтезировали с помощью iScriptTM Reverse Transcription Supermix for RT-qPCR (“Bio-Rad”, США) и набора реактивов ОТ M-MuLV–RH (“Биолабмикс”, Россия). Количество РНК на образец – 100 нг. Для анализа экспрессии полученную кДНК разводили в 10 раз. Количественную ПЦР проводили в автоматическом анализаторе LightCycler® 96 (“Roche”, Швейцария) в объеме 15 мкл, включая 7.5 мкл БиоМастер HS-qPCR SYBR Blue (×2) (“Биолабмикс”), 0.4 мкл каждого праймера (10 мкМ), 3 мкл кДНК. Условия реакции c каждой парой праймеров: 95°C, 10 мин; затем 45 циклов – 95°C, 10 с; 61°C, 10 с; 72°C, 15 с. Нуклеотидные последовательности праймеров приведены в табл. 1.
Таблица 1.
Праймеры, использованные для анализа экспрессии генов
| Праймер | Нуклеотидная последовательность (5'→3') | Источник |
|---|---|---|
| PAIN1_rt_F | GAGGCTCCGGGAGTTGGTAA | [24] |
| PAIN1_rt_R | CCAAATCTTGACGGAGGCAG | |
| SUS4_rt_F | GCATCCTTTCCTAAACCAACC | Разработаны в данной работе |
| SUS4_rt_R | GAGGTATTCCTCTGCCTTCC | |
| Inh2_rt_F | ACCACCCTAGGCCTCATCAT | |
| Inh2_rt_R | ACGGCTACCGTTACATCAGC | |
| APRT_rt_F | GAACCGGAGCAGGTGAAGAA | [33] |
| APRT_rt_R | GAAGCAATCCCAGCGATACG | |
| ef1α_rt_F | ATTGGAAACGGATATGCTCCA | [24] |
| ef1α_rt_R | TCCTTACCTGAACGCCTGTCA | |
| sec3_rt_F | GCTTGCACACGCCATATCAAT | |
| sec3_rt_R | TGGATTTTACCACCTTCCGCA |
Предварительно экспрессию референсных генов-кандидатов APRT (CK270447), EF-1-α (AB061263) и SEC3A (XM_006342541.2) [24] анализировали в различных точках, для итогового анализа в качестве референсного выбрали ген APRT (аденин-фосфорибозилтрансфераза), имеющий наиболее стабильный уровень экспрессии в клубнях. Стабильный уровень экспрессии этого гена в клубнях картофеля при холодовом стрессе показан и в работе [33]. Уровень транскриптов вакуолярной инвертазы (Pain-1, HQ110080.1), сахарозосинтазы (Sus4, NM_001288357.1) и ингибитора инвертазы (Inh2, XM_006349784.2) относительно референсного гена оценивали при помощи LightCycler® 96 Software.
Статистическую обработку результатов проводили с помощью однофакторного дисперсионного анализа (ANOVA) с последующим использованием критерия попарных сравнений Тьюки (https:// a-statsa.com/OneWay_Anova_with_TukeyHSD/). Значимыми считали p ≤ 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Содержание крахмала
Содержание крахмала определяли ферментативным методом в трех клубнях каждого сорта (Невский, Никулинский, Симфония) (табл. 2). Можно отметить, что содержание крахмала в клубнях этих сортов соответствует данным оригинаторов и не выходит за пределы групп.
Таблица 2.
Содержание крахмала в клубнях картофеля
| Сорт | Среднее содержание, % | Данные Гос. реестра сортов России (http://reestr.gossort.com) |
|---|---|---|
| Невский | 13.3 ± 0.40 | 12.0–14.0 |
| Симфония | 15.5 ± 0.15 | 13.0–19.0 |
| Никулинский | 17.5 ± 0.40 | 12.0–21.0 |
Содержание сахаров
Результаты нашего исследования показали, что в клубнях картофеля при хранении при низких температурах (4 и 8°C) происходит интенсивное накопление редуцирующих сахаров и сахарозы. Содержание сахаров, низкое в свежесобранных клубнях всех сортов, увеличивается по мере хранения (рис. 2а). При хранении при температуре 12ºC, наоборот, наблюдалось снижение содержания сахаров в клубнях (рис. 2а).
Рис. 1.
Гены, вовлеченные в процесс холодового осахаривания клубней картофеля. SS – крахмалсинтазы, INV – инвертазы (вакуолярная и апопластические), INH – ингибиторы инвертаз, Sbe – крахмал-ветвящий фермент, AMY – амилазы, SP – фосфорилаза крахмала, UDP – UDP-глюкоза-пирофосфорилаза, AGP – ADP-глюкоза-пирофосфорилаза, GapC – глицеральдегид-3-фосфат-дегидрогеназа, S6P – сахарозофосфатаза, SUS – сахарозосинтаза, SPS – сахарозофосфатсинтаза.
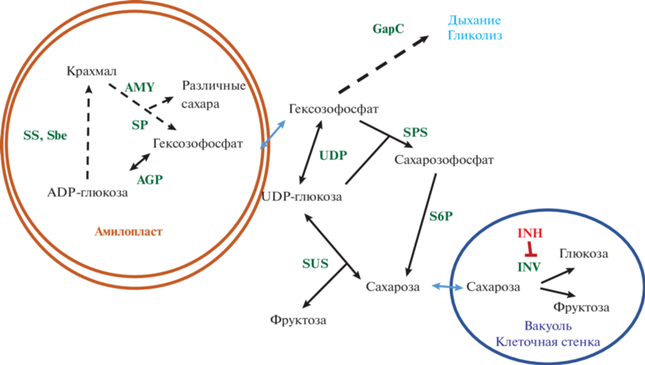
Рис. 2.
Влияние температуры и длительности хранения на содержание общих сахаров (а), глюкозы (б), фруктозы (в) и сахарозы (г) в клубнях картофеля. Каждый столбец отражает среднее значение, вычисленное по трем биологическим повторам. Приведено стандартное отклонение.
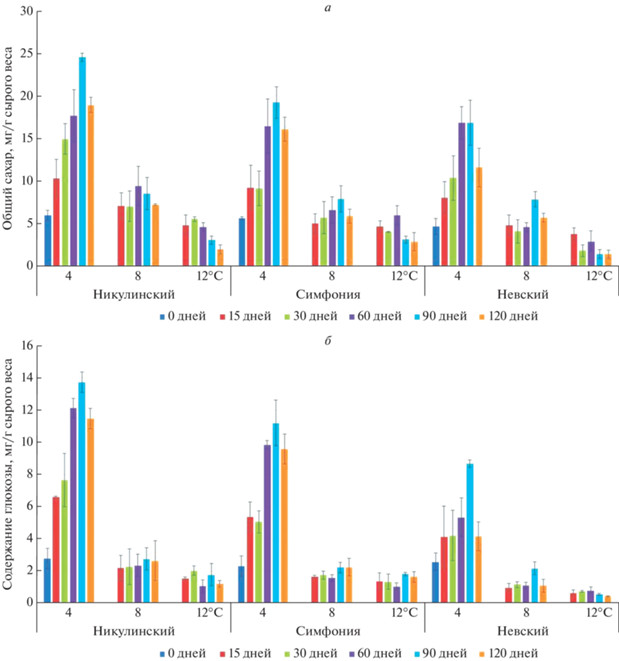
Рис. 2.
Окончание.

Рис. 3.
Относительный уровень экспрессии генов Pain-1 (а), Inh2 (б), SUS4 (в) в контроле и при хранении в течение 15 и 90 дней при 4°C в клубнях сортов Невский, Симфония и Никулинский. Приведены средние значения, вычисленные по трем биологическим повторам, и ошибка среднего. Буквы над столбцами отражают статистически значимые различия (p-value <0.05).

Наиболее интенсивно сахара накапливались при 4°C, причем пик содержания всех трех видов сахаров у каждого из трех сортов приходился на срок 90 дней, далее содержание сахаров немного снижалось (рис. 2а).
Накопление сахаров было наиболее выраженным у сорта Никулинский, наиболее слабым – у сорта Невский. Самым значительным оказалось накопление глюкозы при хранении. Содержание глюкозы в клубнях сорта Никулинский возросло в 5 раз, а у сорта Невский в 3 раза (рис. 2б). Конечное содержание фруктозы было примерно равно исходному содержанию глюкозы, поэтому увеличение содержания фруктозы у сорта Никулинский в 3 раза незначительно повышало уровень общих сахаров (рис. 2в).
Конечное содержание сахарозы было примерно равно исходному содержанию общих сахаров, при этом содержание сахарозы в клубнях сорта Никулинский возросло не более чем в 2.5 раза, что вдвое ниже конечного повышения содержания общих сахаров (рис. 2г). Таким образом, глюкоза вносит наибольший вклад в увеличение содержания общих сахаров в клубнях каждого из трех сортов картофеля, хранящихся в течение 90 дней при 4 и 8°C.
Экспрессия генов
Экспрессию генов мы анализировали при 4°C, так как именно при этой температуре наблюдалось максимальное накопление сахаров, и именно эта температура используется при промышленном хранении картофеля. Были выбраны временные точки – 15 и 90 дней, поскольку в этом временном диапазоне накопление сахаров достигает наибольшего значения, а значит, отражает процессы, происходящие в клубнях и на ранних этапах холодового стресса, и на поздних – в точке перехода в стадию снижения концентрации сахаров. Результаты оценки экспрессии генов приведены на рис. 3 . Показано, что экспрессия всех трех исследуемых генов под воздействием низких температур увеличилась по сравнению с контролем (0 дней), однако степень увеличения зависела от сорта. Экспрессия гена вакуолярной инвертазы в клубнях сорта Симфония увеличилась наиболее значимо по сравнению с остальными сортами, достигая максимального значения на 15-й день и снижаясь через 90 дней до уровня контрольной точки. В клубнях сорта Никулинский экспрессия гена вакуолярной инвертазы также значимо повысилась на 15-й день и также вернулась к первоначальному состоянию через 90 дней. У сорта Невский экспрессия инвертазы увеличилась через 15 дней и оставалась примерно на том же уровне через 90 дней (рис. 3 а).
Экспрессия гена ингибитора вакуолярной инвертазы увеличилась через 15 дней в клубнях сортов Невский и Никулинский и снизилась через 90 дней. В клубнях сорта Симфония экспрессия данного гена, если и увеличилась, то весьма незначительно на 15-й день и немного снизилась в конечной точке (рис. 3 б).
Экспрессия гена сахарозосинтазы значительно увеличилась через 15 дней в клубнях сортов Никулинский и Невский и незначительно – у Симфонии. Через 90 дней экспрессия этого гена у сорта Симфония повысилась так же заметно, как у двух других сортов на 15-й день, и оставалась стабильно высокой у сортов Никулинский и Невский (рис. 3 в).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Хранение картофеля в условиях холода предотвращает их прорастание и гниение, но вызывает холодовое осахаривание, что негативно сказывается на товарных качествах клубней. В нашей работе проанализирована динамика накопления сахаров в клубнях трех сортов картофеля при низких температурах, а также экспрессия ключевых генов-кандидатов, отвечающих за процесс холодового осахаривания.
Установлено, что свежесобранные клубни всех изученных нами сортов характеризовались низким содержанием сахаров и накапливали сахара при хранении при низких температурах. Количество сахаров, накапливаемых при 4°C, было больше, чем при 8 и 12°C независимо от сорта, т.е. степень накопления сахаров отрицательно коррелировала с температурой хранения. Это согласуется с данными предыдущих исследований [15, 34]. Основной вклад в накопление общих сахаров вносила глюкоза, концентрация которой в начальной и во всех последующих точках измерения значительно превышала концентрацию фруктозы и сахарозы (рис. 2б). Большие концентрации глюкозы свидетельствуют об активном разложении крахмала и сахарозы на простые сахара, что наблюдалось и у других сортов, также подверженных холодовому осахариванию [15, 24]. Интересно, что в нашем исследовании концентрации глюкозы превышали концентрации фруктозы, хотя обычно после хранения на холоде концентрации этих двух сахаров имели близкие значения [24, 35]. Возможно, это связано с особенностями генотипов использованных сортов или с активностью других ферментов, ответственных за метаболизм углеводов. Об этом свидетельствует тот факт, что сорта изначально различались по уровню сахаров в клубнях и динамике их накопления (рис. 2).
Содержание сахаров у сортов Никулинский и Симфония было во всех точках выше, чем у сорта Невский. Концентрация сахаров может быть пропорциональной концентрации крахмала – основного источника редуцирующих сахаров, образующихся в результате его гидролиза. Содержание крахмала в клубнях сорта Невский по нашим данным составляет 13.3%, в то время как сорта Никулинский и Симфония содержат 17.5 и 15.5% крахмала соответственно.
Накопление сахаров достигало пика на 90-й день хранения, далее их концентрация снижалась, хотя и не в одинаковой степени у разных сортов. Подобное снижение отмечено, например, Abbasi K.S. и соавт. [36], которые наблюдали снижение содержания общих сахаров после 70 дней хранения. Это снижение может быть связано с активацией окисления редуцирующих сахаров в процессе дыхания [37] или с ресинтезом крахмала в клубнях [22].
Нами выявлена активация экспрессии генов вакуолярной инвертазы (Pain-1), сахарозосинтазы (SUS4) и ингибитора инвертазы (Inh2) в ответ на воздействие низких температур (рис. 3 ), что согласуется с данными [20, 23, 24]. Однако заметные различия паттерна экспрессии этих генов в разных сортах могут свидетельствовать о генотипзависимой регуляции холодового осахаривания.
Полученные нами данные на позволяют однозначно утверждать, что вакуолярная инвертаза является ключевым ферментом и основным лимитирующим фактором холодового осахаривания, как предполагалось ранее [22], так как повышение экспрессии гена Pain-1, наблюдаемое у всех сортов в начале срока хранения, было статистически значимым только у сорта Симфония. У двух же других сортов, включая Никулинский, который характеризуется самым высоким уровнем накопления сахаров, степень повышения экспрессии гена Pain-1 была не столь значительной (рис. 3 а). Кроме того, у сорта Никулинский эта степень отрицательно коррелировала с уровнем экспрессии гена ингибитора инвертазы Inh2, который статистически значимо увеличился, особенно при сопоставлении с низким уровнем экспрессии этого гена в клубнях сорта Симфония (рис. 3 б). Следовательно, можно предположить, что активность гена Pain-1 в клубнях сорта Никулинский снижена за счет ее ингибирования продуктом гена Inh2, и она не вносит существенного вклада в осахаривание. Пик экспрессии гена вакуолярной инвертазы спустя 7–14 дней хранения с последующим ее снижением наблюдали и ранее [20, 23]. Уровень экспрессии гена вакуолярной инвертазы в клубнях всех сортов снизился практически до исходных значений через 90 дней хранения, а экспрессия гена сахарозосинтазы значимо повысилась, начиная с 15-го дня хранения (рис. 3 в). При этом уровень экспрессии гена сахарозосинтазы оставался повышенным и на 90-й день хранения, в отличие от инвертазы.
Повышенная экспрессия гена сахарозосинтазы сохраняется даже после хранения клубней в условиях холода в течение нескольких месяцев, что согласуется с данными [20, 24], указывающими на важную роль сахарозосинтазы в холодовом осахаривании. Согласно Baroja-Fernández E. и соавт. [19], существует обратная корреляция между уровнями экспрессии и активности инвертазы и сахарозосинтазы, кроме того, в трансгенных клубнях с повышенной экспрессией гена сахарозосинтазы наблюдалось повышенное содержание крахмала. Известно, что сахароза, поступающая из апопласта в цитозоль клеток клубней, гидролизуется сахарозосинтазой до UDP-глюкозы и фруктозы. Следовательно, от уровня экспрессии этого фермента может зависеть степень накопления моносахаридов. Однако на степень накопления может влиять и обратный процесс – скорость синтеза олиго- и полисахаридов, особенно крахмала. В работе Liи Х. и соавт. [22] показана активация генов синтеза крахмала в растениях картофеля с генотипом, устойчивым к холодовому осахариванию. Кроме того, Shumbe L. и соавт. [15] наблюдали повышение экспрессии сахарозосинтазы в клубнях сорта, устойчивого к осахариванию, по сравнению с чувствительным сортом. Таким образом, точная роль сахарозосинтазы, как и двух других изученных в нашей работе ферментов, в процессах метаболизма углеводов в клубнях картофеля до сих пор не определена.
В нашей работе мы анализировали гены, которые, как считается, вносят основной, но не единственный вклад в холодовое осахаривание. Известно, что с накоплением сахаров при воздействии холода связаны и другие гены углеводного метаболизма – как структурные, так и регуляторные. Щелочные/нейтральные инвертазы гидролизуют сахарозу в цитоплазме в ответ на воздействие низких температур [38], экспрессия амилаз и их ингибиторов коррелирует с накоплением редуцирующих сахаров в клубнях [39]. Некоторые аллели гена UDP-глюкоза-пирофосфорилазы ассоциированы с устойчивостью к холодовому осахариванию [40, 41]. Как упоминалось выше, вакуолярная инвертаза может регулироваться не только INH2, но и ингибитором INH1, а также протеинкиназой SnRK1 [42]. Недавно показали, что транскрипционный фактор StTINY3 влияет на устойчивость к холодовому осахариванию, повышая экспрессию ингибитора инвертазы INH2 и взаимодействуя с другими генами метаболизма углеводов [43]. Скорее всего, эти гены обуславливают также генотип-специфичную регуляцию холодового осахаривания, поэтому требуется дальнейшее изучение роли генов в данном процессе.
ЗАКЛЮЧЕНИЕ
Впервые показана генотип-специфичная динамика накопления редуцирующих сахаров в клубнях отечественных сортов картофеля Никулинский, Невский и голландского сорта Симфония при хранении при низких температурах, а также выявлены различия в экспрессии генов, связанных с процессом холодового осахаривания. Выдвинуто предположение о различном вкладе ферментов, кодируемых данными генами, в регуляцию холодового осахаривания. Полученные результаты способствуют расширению представлений о молекулярно-генетических механизмах реакции растений на воздействие низких температур.
Авторы выражают благодарность С.В. Герасимовой и Д.А. Афонникову за помощь в проведении исследования и при написании статьи.
Анализ содержания сахаров в процессе хранения проведен в рамках подпрограммы КНТП “Развитие селекции и семеноводства картофеля в Российской Федерации” проект № FWNR-2019-0012. Разработка праймеров и анализ экспрессии генов выполнен в рамках бюджетного проекта № FWNR-2022-0017.
Настоящая статья не содержит каких-либо исследований с использованием животных в качестве объектов.
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Bianchi G., Scalzo R.L., Testoni A., Maestrelli A. (2014) Nondestructive analysis to monitor potato quality during cold storage. J. Food Quality. 37, 9–17.
Zhang H., Hou J., Liu J., Zhang J., Song B., Xie C. (2017) The roles of starch metabolic pathways in the cold-induced sweetening process in potatoes. Starch-Stärke. 69, 1600194.
McCullough M.L., Hodge R.A., Um C.Y., Gapstur S.M. (2019) Dietary acrylamide is not associated with renal cell cancer risk in the CPS-II nutrition cohort. Cancer Epidemiol. Prevention Biomarkers. 28, 616–619.
Sowokinos J.R. (2001) Biochemical and molecular control of cold-induced sweetening in potatoes. Am. J. Potato Res. 78, 221–236.
Amrein T.M., Schönbächler B., Rohner F., Lukac H., Schneider H., Keiser A., Escher F., Amadò R. (2004) Potential for acrylamide formation in potatoes: data from the 2003 harvest. Eur. Food Res. Technol. 219, 572−578.
Chen S., Hajirezaei M.R., Zanor M.I., Hornyik C., Debastn S., Lacomme C., Fernie A.R., Sonnewald U., Boernke F. (2008) RNA interference-mediated repression of sucrose-phosphatase in transgenic potato tubers (Solanum tuberosum) strongly affects the hexose-to-sucrose ratio upon cold storage with only minor effects on total soluble carbohydrate accumulation. Plant Cell Environ. 31, 165–176.
Xiong X., Tai G.C.C., Seabrook J.E.A., Wehling P. (2002) Effectiveness of selection for quality traits during the early stage in the potato breeding population. Plant Breed. 121, 441–444.
Hamernik A.J., Hanneman R.E., Jansky S.H. (2009) Introgression of wild species germplasm with extreme resistance to cold sweetening into the cultivated potato. Crop Sci. 49, 529–542.
Liu X., Zhang C., Ou Y., Lin Y., Song B., Xie C., Liu J., Li X.Q. (2011) Systematic analysis of potato acid invertase genes reveals that a cold-responsive member, StvacINV-1, regulates cold-induced sweetening of tubers. Mol. Genet. Genom. 286, 109–118.
Bhaskar P.B., Wu L., Busse J.S., Whitty B.R., Hamernik A.J., Jansky S.H., Jiang J. (2010) Suppression of the vacuolar invertase gene prevents cold-induced sweetening in potato. Plant Physiol. 154, 939–948.
Clasen B.M., Stoddard T.J., Luo S., Demorest Z.L., Li J., Cedrone F., Tibebu R., Davison S., Ray E.E., Daulhac A., Coffman A. (2015) Improving cold storage and processing traits in potato through targeted gene knockout. Plant Biotechnol. J. 14, 169–176.
Draffehn A.M., Meller S., Li L., Gebhardt C. (2010) Natural diversity of potato (Solanum tuberosum) invertases. BMC Plant Biol. 10, 1−15.
Слугина М.А., Снигирь Е.А., Рыжова Н.Н., Кочиева Е.З. (2013) Структура и полиморфизм фрагмента локуса Pain-1, кодирующего вакуолярную инвертазу видов Solanum. Молекуляр. биология. 47, 243–243.
Ou Y., Song B., Liu X., Xie C., Li M., Lin Y., Zhang H., Liu J. (2013) Promoter regions of potato vacuolar invertase gene in response to sugars and hormones. Plant Physiol. Biochem. 69, 9–16.
Shumbe L., Visse M., Soares E., Smit I., Dupuis B., Vanderschuren H. (2020) Differential DNA methylation in the Vinv promoter region controls cold induced sweetening in potato. bioRxiv. 062562.
Brummell D.A., Chen R.K., Harris J.C., Zhang H., Hamiaux C., Kralicek A.V., McKenzie M.J. (2011) Induction of vacuolar invertase inhibitor mRNA in potato tubers contributes to cold-induced sweetening resistance and includes spliced hybrid mRNA variants. J. Exp. Botany. 62, 3519–3534.
Liu X., Lin Y., Liu J., Song B., Ou Y., Zhang H., Li M., Xie C. (2013) StInvInh2 as an inhibitor of Stvac INV 1 regulates the cold-induced sweetening of potato tubers by specifically capping vacuolar invertase activity. Plant Biotechnol. J. 11, 640–647.
Liu X., Cheng S., Liu J., Ou Y., Song B., Zhang C., Lin Y., Li X., Xie C. (2013) The potato protease inhibitor gene, St-Inh, plays roles in the cold-induced sweetening of potato tubers by modulating invertase activity. Postharvest Biol. Tec. 86, 265–271.
Baroja-Fernández E., Muñoz F.J., Montero M., Etxeberria E., Sesma M.T., Ovecka M., Bahaji A., Ezquer I., Li J., Prat S., Pozueta-Romero J. (2009) Enhancing sucrose synthase activity in transgenic potato (Solanum tuberosum L.) tubers results in increased levels of starch, ADPglucose and UDPglucose and total yield. Plant Cell Physiol. 50, 1651–1662.
Bagnaresi P., Moschella,A., Beretta O., Vitulli F., Ranalli P., Perata P. (2008) Heterologous microarray experiments allow the identification of the early events associated with potato tuber cold sweetening. BMC Genomics. 9, 1–23.
Baldwin S.J., Dodds K.G., Auvray B., Genet R.A., Macknight R.C., Jacobs J.M.E. (2011) Association mapping of cold-induced sweetening in potato using historical phenotypic data. Ann. Appl. Biol. 158, 248–256.
Liu X., Chen L., Shi W., Xu X., Li Z., Liu T., He Q., Xie C., Nie B., Song B. (2021) Comparative transcriptome reveals distinct starch-sugar interconversion patterns in potato genotypes contrasting for cold-induced sweetening capacity. Food Chem. 334, 127550.
Wiberley-Bradford A.E., Bethke P.C. (2017) Rate of cooling alters chip color, sugar contents, and gene expression profiles in stored potato tubers. Am. J. Potato Res. 94, 534–543.
Slugina M.A., Shchennikova A.V., Meleshin A.A., Kochieva E.Z. (2020) Homologs of vacuolar invertase inhibitor INH2 in tuber-bearing wild potato species and Solanum tuberosum: gene polymorphism and co-expression with saccharolytic enzyme genes in response to cold stress. Sci. Horticult. 269, 109425.
Дорошков А.В., Симонов А.В., Сафонова А.Д., Афонников Д.А., Лихенко И.Е., Колчанов Н.А. (2016) Оценка количественных характеристик опушения листьев картофеля с использованием анализа цифровых микроизображений. Достижения науки и техники АПК. 30, 12–14.
Альт В.В., Гурова Т.А., Елкин О.В., Клименко Д.Н., Максимов Л.В., Пестунов И.А., Дубровская О.А., Генаев М.А., Эрст Т.В., Генаев К.А., Комышев Е.Г., Хлесткин В.К., Афонников Д.А. (2020) Использование гиперспектральной камеры Specim IQ для анализа растений. Вавил. Журн. Генет. Селекции. 24, 259–266.
Антонова О.Ю., Швачко Н.А., Новикова Л.Ю., Шувалов О.Ю., Костина Л.И., Клименко Н.С., Шувалова А.Р., Гавриленко Т.А. (2016). Генетическое разнообразие сортов картофеля российской селекции и стран ближнего зарубежья по данным полиморфизма SSR-локусов и маркеров R-генов устойчивости. Вавил. Журн. Генет. Селекции. 20, 596–606.
Totsky I.V., Rozanova I.V., Safonova A.D., Batov A.S., Gureeva Yu.A., Khlestkina E.K., Kochetov A.V. (2021) Genotyping of potato samples from the GenAgro ICG SB RAS collection using DNA markers of genes conferring resistance to phytopathogens. J. Genet. Breed. 25, 677–686.
Khlestkin V.K., Erst T.V., Rozanova I.V., Efimov V.M., Khlestkina E.K. (2020) Genetic loci determining potato starch yield and granule morphology revealed by genome-wide association study (GWAS). Peer. J. 8, e10286.
Khlestkin V.K., Rozanova I.V., Efimov V.M. Khlestkina E.K. (2019) Starch phosphorylation associated SNPs found by genome-wide association studies in the potato (Solanum tuberosum L.). BMC Genet. 20, 45–53.
Ibragimova S., Romanova A., Saboiev I., Salina E., Kochetov A. PlantGen2021: The 6th Internat. Sci. Conf. (2021) Novosibirsk, Russia. Abstract book. P. 95. Abstract 79.
Туркина М.В., Соколова С.В. (1971) Методы определения моносахаридов и олигосахаридов. Биохимические методы в физиологии растений. Москва: Наука, 7–34.
Lopez-Pardo R., Ruiz de Galarreta J.I., Ritter E. (2013) Selection of housekeeping genes for qRT-PCR analysis in potato tubers under cold stress. Mol. Breed. 31, 39–45.
Matsuura-Endo C., Ohara-Takada A., Chuda Y., Ono H., Yada H., Yoshida M., Kobayashi A., Tsuda S., Takigawa S., Noda T. (2006) Effects of storage temperature on the contents of sugars and free amino acids in tubers from different potato cultivars and acrylamide in chips. Biosci. Biotechnol. Biochem. 70, 1173–1180.
Lin Q., Xie Y., Guan W., Duan Y., Wang Z., Sun C. (2019) Combined transcriptomic and proteomic analysis of cold stress induced sugar accumulation and heat shock proteins expression during postharvest potato tuber storage. Food Chem. 297, 124991.
Abbasi K.S., Masud T., Qayyum A., Khan S.U., Abbas S., Jenks M.A. (2016) Storage stability of potato variety Lady Rosetta under comparative temperature regimes. Sains Malaysiana. 45, 677–688.
Sonnewald U. (2001) Control of potato tuber sprouting. Trends Plant Sci. 6, 333–335.
Datir S.S., Regan S. (2022) Role of alkaline/neutral invertases in postharvest storage of potato. Postharvest. Biol. Technol. 184, 111779.
Слугина М.А., Кочиева Е.З. (2018) Использование генов углеводного обмена для улучшения качества клубней картофеля (Solanum tuberosum L.). Сельскохоз. биология. 53, 450–463.
Gupta S.K., Crants J. (2019) Identification and impact of stable prognostic biochemical markers for cold-induced sweetening resistance on selection efficiency in potato (Solanum tuberosum L.) breeding programs. PLoS One. 14, e0225411.
McKenzie M.J., Sowokinos J.R., Shea I.M., Gupta S.K., Lindlauf R.R., Anderson J.A. (2005) Investigations on the role of acid invertase and UDP-glucose pyrophosphorylase in potato clones with varying resistance to cold-induced sweetening. Am. J. Potato Res. 82, 231–239.
Lin Y., Liu T., Liu J., Liu X., Ou Y., Zhang H., Li M., Sonnewald U., Song B., Xie C. (2015) Subtle regulation of potato acid invertase activity by a protein complex of invertase, invertase inhibitor, and sucrose nonfermenting1-related protein kinase. Plant Physiol. 168, 1807–1819.
Shi W., Ma Q., Yin W., Liu T., Song Y., Chen Y., Song L., Sun H., Hu S., Liu T., Jiang R. (2022) StTINY3 enhances cold-induced sweetening resistance by coordinating starch resynthesis and sucrose hydrolysis in potato. J. Exp. Botany. 73. 4968–4980.
Дополнительные материалы отсутствуют.
Инструменты
Молекулярная биология