

Журнал общей биологии, 2019, T. 80, № 5, стр. 348-363
К вопросу о протяженности гидроплазматических течений у колониального гидроида Dynamena pumila (L., 1758)
Н. Н. Марфенин 1, *, В. С. Дементьев 1
1 Московский государственный университет им. М.В. Ломоносова,
биологический факультет, каф. зоологии беспозвоночных
119234 Москва, Ленинские горы, 1, Россия
* E-mail: marf47@mail.ru
Поступила в редакцию 28.01.2019
После доработки 08.04.2019
Принята к публикации 26.06.2019
Аннотация
Статья посвящена исследованию протяженности гидроплазматических течений (ГПТ) у колониального гидроида Dynamena pumila (L., 1758) и детализации ранее изложенных результатов. Использован метод визуального сканирования колоний под микроскопом и регистрации в каждом модуле столона направления течений и насыщенности их пищевыми частицами. В результате удалось установить, что неразрывное однонаправленное ГПТ может соответствовать по протяженности расстоянию от основания материнского побега до верхушки столона. Получено подтверждение теории последовательного участия промежуточных побегов в продвижении гидроплазмы по столону и формировании течения. Перемещение пищевых частиц по столону возможно не только вследствие поступления гидроплазмы из одного источника (материнского побега), но и при последовательном ответвлении ГПТ в побеги по пути продвижения, что активизирует участие промежуточных побегов в дальнейшем продвижении частиц по столону.
Функциональная интеграция организма колониальных гидроидов осуществляется посредством перемещения внутриполостной жидкости (гидроплазмы) по трубковидному телу, представленному побегами и соединяющими их столонами (рис. 1). Гидроплазма перемещается по колонии за счет поперечных (латеральных) пульсаций ценосарка и гидрантов, что приводит к уменьшению величины просвета канала в одних местах и расширению в других (Hale, 1960; Fulton, 1963; Марфенин, 2002, 2016). Такая распределительная система была названа пульсаторно-перистальтической (Марфенин, 1985). Переносимые гидроплазмой пищевые частицы позволяют отслеживать ее перемещение. Ранее было показано, что гидроплазматические течения (ГПТ) могут быть достаточно протяженными и продолжительными, чтобы транспортировать пищу по всему колониальному организму (Карлсен, Марфенин, 1976, 1984). Эти данные были получены с помощью непосредственных наблюдений за перемещением частиц в колонии, причем первичные результаты регистрации, за исключением нескольких примеров, не были опубликованы (Марфенин, 1985, 1988). Впоследствии вывод о существовании протяженных ГПТ был подвергнут сомнению на основании таких же непосредственных наблюдений (Бурыкин, 2010, 2013, 2015). Это побудило нас провести дополнительные исследования с использованием простого метода сканирования колоний под микроскопом и регистрации направления ГПТ в каждом модуле столона. Выяснение истинной протяженности магистральных и компенсаторных ГПТ необходимо для определения способа и эффективности функционирования распределительной системы, выполняющей ключевую роль в жизнедеятельности колонии.
Цель исследования – определение протяженности неразрывных ГПТ в столоне колониального гидроида Dynamena pumila (L., 1758) на основе регулярной регистрации направления перемещения частиц в гастроваскулярной полости. Полученные данные необходимы для понимания процесса генерации ГПТ и масштаба функциональной интеграции у колониальных гидроидов.
Функционирование распределительной системы пульсаторно-перистальтического типа наименее изучено вследствие ряда методических сложностей, связанных, прежде всего, с непостоянством направления и скорости перемещения гидроплазмы. Немногочисленные исследования выполнены на книдариях, у которых ток гидроплазмы происходит по параллельным каналам в противоположных направлениях (Winkle, Blackstone, 1997; Parrin et al., 2010; Harmata et al., 2013), или же ограничиваются регистрацией пульсаций ценосарка (Blackstone, 1996) и усвоения пищи в отдельных местах (Wagner et al., 1998; Dudgeon et al., 1999). Изучению закономерностей перемещения пищи в колониях с пульсаторно-перистальтической распределительной системой посвящен ряд работ (Карлсен, Марфенин, 1976, 1984; Марфенин, 1985, 1993; Бурыкин, 2006, 2008, 2010, 2013, 2015).
Настоящее исследование продолжает серию работ авторов, посвященную изучению перемещений гидроплазмы в колониальных гидроидах (Дементьев, Марфенин, 2017, 2018, 2019; Марфенин, Дементьев, 2017, 2018а, б; Marfenin, Dementyev, 2017).
МАТЕРИАЛ И МЕТОДЫ
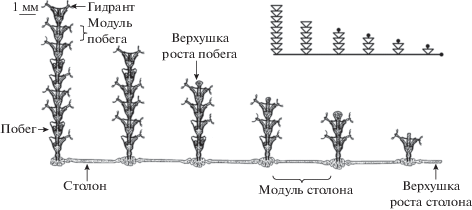
Для исследования был выбран колониальный гидроид Dynamena pumila (L.) (Sertulariidae, Leptothecata, Hydrozoa) – пример модульного организма (рис. 1), в котором гидранты расположены на побегах, а побеги на примерно одинаковых расстояниях отходят от прямолинейных столонов, редко образующих анастомозы. Гидранты на побегах расположены в два ряда строго супротивно. На стволах побегов и первичных столонах образуются боковые ветви, устроенные так же, как и те, от которых они отходят. Гидранты вместе с соединяющим их трубковидным ценосарком образуют единое тело разветвленного модульного организма, в котором все составляющие связаны между собой не только телесно, но и общей гастроваскулярной полостью, заполненной жидкостью – гидроплазмой.
В модульных организмах, к которым относятся и колониальные гидроиды, тело построено только из повторяющихся частей нескольких разновидностей, которые принято называть модулями. Модулем или сегментом столона мы будем называть участок столона между двумя побегами, а модулем побега – пару супротивно расположенных гидрантов и участок ценосарка ствола между ними (рис. 1). Расширенные дистальные участки ценосарка в столонах и побегах называются верхушками роста, так как с ними связано удлинение ценосарка. У нерастущих столонов и побегов торцевые участки ценосарка обычно не расширены.
Сбор и культивирование гидроидов осуществляли по проведенной ранее методике (Дементьев, Марфенин, 2017, 2018, 2019; Марфенин, Дементьев, 2017, 2018а, б; Marfenin, Dementyev, 2017). Исследование осуществлено на Беломорской биологической станции (ББС) им. Н.А. Перцова биологического факультета МГУ (пос. Приморский), находящейся на берегу пролива Великая Салма Кандалакшского залива Белого моря. Материал для исследования был собран в июле 2016 г. на Еремеевском пороге Ругозерской губы Кандалакшского залива. Колонии D. pumila выращивали из фрагментов отдельных побегов, которые были вырезаны из зарослей гидроида на талломе Fucus serratus с тем, чтобы получить максимально генетически однородный материал, и закреплены по методу Кровелла (Crowell, 1957) на предметных стеклах. В процессе репаративной регенерации материнский побег на месте среза прилипал к стеклу выделяемым на верхушке роста перисарком. В дальнейшем формировался столон. Формирующиеся от материнского побега колонии кормили ежедневно двухдневной культурой науплиев Artemia salina (L., 1758). Предметные стекла с колониями содержали в замкнуто-проточных аэрируемых аквариумах в контролируемых условиях: постоянная температура 15 ± 1.5°C, соленость 26 ± 1‰, автоматизированная циркуляция морской воды (при помощи помпы) через проточный холодильник, ежедневное кормление науплиями в концентрации 1.6–3.4 экз./мл. Замену воды в аквариумах на чистую морскую производили раз в три дня. Во время регистрации распределения ГПТ по колонии температура оставалась в пределах 13.7–14.3°С (табл. 1).
Таблица 1.
Характеристики исследованных колоний и показатели динамики гидроплазматических течений в них, полученные методом визуального посегментного сканирования модулей столона
| № сканирования ГПТ (бланка) | № колонии | Дата сканирования | t, °С | Длина столона, мм | Число побегов | Время от начала кормления до начала регистрации, ч | Начальная концентрация науплиев, экз./мл | Доля ГПТ, % | Доля покоя, % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ненасыщенных (+)ГПТ | насыщенных (+)ГПТ | всего (+)ГПТ | ненасыщенных (–)ГПТ | насыщенных (–)ГПТ |
всего (–)ГПТ | всего ГПТ | |||||||||
| 1 | 8-6 (2016) | 15 июля 2016 | 14 | 20.6 | 6 | 4.5 | 2.1 | 8.5 | 1.4 | 9.9 | 7.5 | 0.2 | 7.7 | 17.6 | 82.4 |
| 4 | 2-5 (2016) | 17 июля 2016 | 14.3 | 27.9 | 7 | 3 | 11 экз. дозированно | 6.2 | 32.3 | 38.5 | 14.0 | 20.6 | 34.6 | 73.1 | 26.9 |
| 7 | 1-8 (2016) | 19 июля 2016 | 13.8 | 23.1 | 7 | 4 | 1.6 | 3.9 | 9.8 | 13.7 | 9.1 | 8.1 | 17.2 | 30.9 | 69.1 |
| 11 | 8-6 (2016) | 21 июля 2016 | 14.2 | 24 | 7 | 1 | 12 экз. дозированно | 4.2 | 8.6 | 12.8 | 7.8 | 11.7 | 19.5 | 32.3 | 67.7 |
| 16 | 5-7 (2016) | 24 июля 2016 | 13.7 | 22.4 | 7 | 72 | 2.3 | 3.9 | 8.9 | 12.8 | 13.9 | 4.9 | 18.8 | 31.6 | 68.4 |
Для исследования использовали прямолинейные колонии с неразветвленными столонами, в которых по числу гидрантов доминировали материнские побеги, из которых они были выращены. Остальные побеги включали всего по несколько пар гидрантов каждый. Перед регистрацией пространственного распределения ГПТ в столоне составляли схемы колонии по методу картирования (Марфенин, 1980). Общая картина ГПТ была получена нами для восьми колоний (16 бланков учета), состоявших из шести–восьми побегов. Результаты пяти подобных бланков регистрации, являющихся наиболее типичными, были взяты в качестве образца для данной публикации (табл. 1). В колонии № 8-6 (2016) регистрацию системы ГПТ провели дважды с промежутком в шесть суток.
Исследованные колонии:
Бланк 1. Колония № 8-6 (2016) – через 4.5 ч после кормления;
Бланк 4. Колония № 2-5 (2016) – через 3 ч после кормления (получила дозированно 11 науплиев в некоторые гидранты);
Бланк 7. Колония № 1-8 (2016) – через 4 ч после кормления;
Бланк 11. Колония № 8-6 (2016) повторно – через 1 ч после кормления (получила дозированно 12 науплиев), спустя шесть дней после первой съемки;
Бланк 16. Колония № 5-7 (2016) – спустя 3 дня голодания.
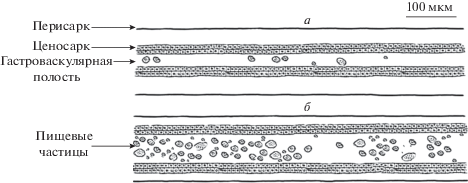
Протяженность неразрывного гидроплазматического течения определяли визуально, просматривая каждую минуту колонию на просвет в течение 10–20 с и нанося на схему колонии посегментно (по модулям столона) направление ГПТ и насыщенность ГПТ частицами в двух градациях: ненасыщенные ГПТ – мало частиц (тонкие стрелки → или ←) и насыщенные ГПТ – много частиц (толстые стрелки < или >). Под ненасыщенными ГПТ мы подразумеваем перемещение гидроплазмой отдельных разрозненных частиц. Насыщенные течения характеризуются множеством частиц, целиком заполняющих просвет гастроваскулярной полости в пределах поля зрения (рис. 2). При этом просвет ценосарка, как правило, расширен, поэтому насыщенные ГПТ мы также называем сильными, а ненасыщенные – слабыми. Размер частиц специально не учитывали: мелкие и крупные частицы могли присутствовать как в насыщенных, так и в ненасыщенных течениях. Процедуру регулярной регистрации ГПТ в каждом модуле столона колонии мы называем посегментной регистрацией или сканированием ГПТ. На каждом бланке все строки пронумерованы в формате времени (0:00), что позволяет ссылаться в тексте статьи на определенное место бланка регистрации и учитывать продолжительность ГПТ.
Рис. 2.
Фрагмент столона с малым (а) и большим (б) числом взвешенных в гидроплазме частиц. При визуальной посегментной регистрации ГПТ колоний состоянию а соответствуют тонкие стрелки (→ ←), а состоянию б – толстые (> <). Обычно при движении большого скопления частиц (б) просвет гастроваскулярной полости соответствующего модуля расширен.
При регистрации продолжительности стадий перемещения гидроплазмы или его отсутствия не разделяли стадии полного покоя и хаотического перемещения на одном месте, обозначая и ту, и другую (σ). По числу последовательных модулей столона с толстыми однонаправленными стрелками определяли протяженность неразрывного ГПТ – величину LHF (мкм).
Для определения долей фаз покоя, магистрального и компенсаторного ГПТ удобно подсчитывать процентное соотношение ячеек на всем бланке с соответствующими значками: (σ), (>) и (<). В тексте мы называем результаты такого учета как “% модулей” в соответствующем состоянии.
По направлению различали магистральные и компенсаторные ГПТ. Магистральными принято называть протяженные ГПТ, направленные к верхушке роста столона, а компенсаторными – протяженные ГПТ от верхушки столона (Карлсен, Марфенин, 1984). Нумерацию побегов осуществляли от материнского, который считается № I, а модулей столона – от его верхушки, так как морфофункциональное состояние модуля столона зависит от его удаленности от верхушки роста (Marfenin, Dementyev, 2017).
Также высчитали долю покоя в работе распределительной системы по числу модулей столона, в которых не было ГПТ на момент регистрации, от общего их количества в столоне (табл. 1).
РЕЗУЛЬТАТЫ
Большую часть времени частицы в ценосаркальной трубке или остаются в покое, или вертятся на одном месте, или движутся рывками попеременно в противоположных направлениях с меняющейся скоростью и разной продолжительностью движения. Лишь время от времени частицы устремляются потоком в одном направлении, после чего иногда наступает затишье, а затем – движение назад. Нередко обратное перемещение частиц наступает сразу же после окончания поступательного движения. Однонаправленные течения характеризуются большой максимальной скоростью, большей продолжительностью и, как правило, сопровождаются расширением просвета ценосаркальной трубки, т.е. такой поток гидроплазмы быстрее и дольше обычных “подергиваний”. Чаще максимальная протяженность неразрывного ГПТ проявлялась в течение двух–трех минут, т.е. за это время длина ГПТ увеличивалась. Иногда максимальная протяженность ГПТ устанавливалась за одну минуту.
Визуальное сканирование состояния гидроплазмы по модулям столона показало, что на фоне всевозможных вариантов перемещений гидроплазмы проявляются регулярные протяженные ГПТ, насыщенные пищевыми частицами. Рассмотрим разнообразие динамики перемещения гидроплазмы на конкретных примерах.
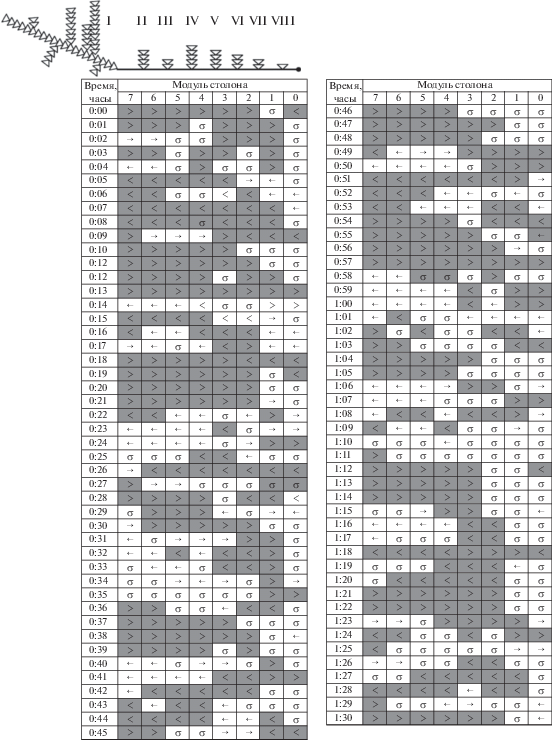
Бланк 16. Колония № 5-7 (2016) является примером распределительной системы, в которой доминирует материнский побег (рис. 3). Насыщенные частицами протяженные ГПТ отмечены на бланке скоплениями серых ячеек со знаками (>) или (<). Несмотря на то, что колония не получала пищу в течение трех дней, в ней были зарегистрированы протяженные и ритмичные магистральные ГПТ (8.9% модулей), а также связанные с ними компенсаторные ГПТ (4.9% модулей). Также преобладали состояния покоя (68.4%) или ненасыщенных ГПТ (18.8%) (табл. 1). Всего за 96 мин было зарегистрировано пять магистральных ГПТ, всякий раз начинавшихся от материнского побега. Насыщенные течения гидроплазмы были достаточно протяженные (показатель LHF). Они охватывали единовременно от двух до восьми модулей столонов, т.е. иногда всю колонию. Максимальной протяженности ГПТ достигали за 2 мин.
Рис. 3.
Бланк 16. Колония № 5-7 (2016) – пример распределительной системы, в которой доминирует материнский побег. Здесь и далее на рис. 4–7 вверху представлена схема колонии; серым цветом выделены протяженные ГПТ.
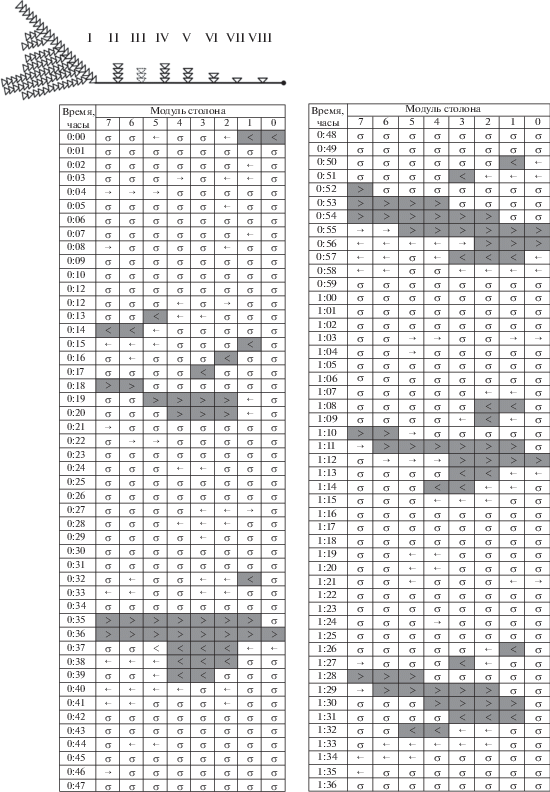
Магистральные ГПТ повторялись каждые 17–18 мин в такой последовательности: 17, 17, 18, 18 мин. Магистральные ГПТ начинались от материнского побега, охватывали несколько модулей столона, затем продвигались далее к верхушке столона, при этом становясь более протяженными, хотя в ближайших к материнскому побегу модулях столона ГПТ могли затихать. Наибольшая протяженность однонаправленного ГПТ (LHF) к верхушке столона на данном примере составляла восемь модулей столона (т.е. около 24 мм), что повторилось три раза из пяти ГПТ. Достигнув верхушки, течение практически без периода покоя обращалось в противоположную сторону (см. строки 0:37, 0:57, 1:13, 1:31 ч) и компенсаторным током двигалось к материнскому побегу. До материнского побега компенсаторное течение дошло в трех случаях из пяти. Изредка выходящее из промежуточного побега течение становилось разнонаправленным (← →): часть его двигалась к верхушке столона, другая – к материнскому побегу (модули 1–2 (0:27 ч), модули 0–1 (1:21 ч)). Продолжительность однонаправленного ГПТ в отдельных модулях, принимая в расчет как насыщенные, так и ненасыщенные ГПТ, не превышала 3 мин. Суммарное число модулей, охваченных магистральными ГПТ, больше, чем компенсаторными ГПТ (табл. 1).
Бланк 7. Колония № 1-8 (2016) получила пищу за 4 ч до визуального сканирования ГПТ. Места распределения проглоченной пищи не известны, но о них можно судить по направлениям ГПТ. Фаза покоя, т.е. доля модулей, в которых не были зарегистрированы ГПТ, составляла 69.1% (табл. 1). На примере этой колонии можно показать инициирующую роль в формировании картины ГПТ не материнского, а промежуточных побегов (рис. 4).
Рис. 4.
Бланк 7. Колония № 1-8 (2016) – пример распределительной системы, в которой доминирует один из промежуточных побегов (№ IV).
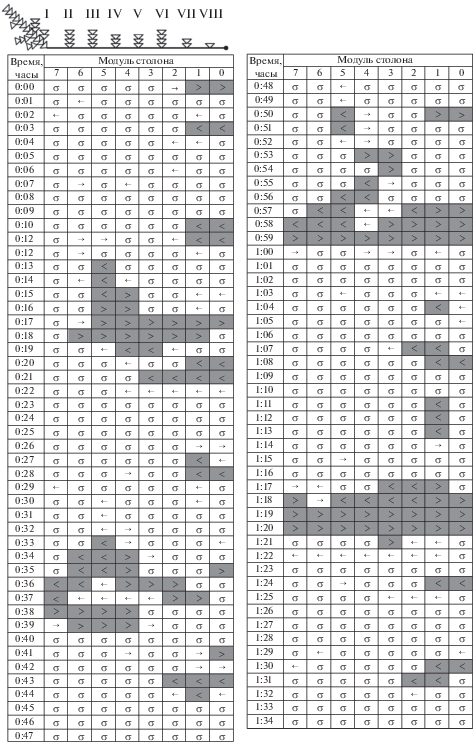
Фаза протяженных ГПТ наступала четыре раза за 1.5 ч сканирования. В трех случаях активность начиналась от побега № IV, который отстоял от материнского на три модуля (около 8 мм). Отчетливо видно, как ведущий побег с периодичностью в 17–19 мин был источником двух разнонаправленных потоков гидроплазмы: модули 4–5 (0:15, 0:34–0:35 ч) и 1–2 (0:57 и 1:17–1:18 ч). Один из них направлялся в сторону материнского побега и либо доходил до него, либо быстро затухал, пройдя около 3 мм столона. В случае достижения потоком материнского побега из последнего спустя 1–2 мин выходило насыщенное ГПТ (0:38, 0:59 ч) в прилежащий седьмой модуль столона. Второй поток устремлялся к верхушке столона, иногда достигая последней (0:17 ч). В двух–трех дистальных модулях столона часто возникали насыщенные, но непротяженные течения, не обладавшие явной периодичностью (строки 0:03, 0:10–0:11, 0:27–0:28, 0:43–0:44, 0:50, 1:04, 1:07–1:08 ч и т.д.). Эти короткие течения не сливались с ГПТ из лидирующего побега и циркулировали между верхушкой столона и расположенными в непосредственной близости от нее побегами, охватывая два–три модуля столона. Под конец регистрации (1:18–1:20 ч) материнский побег стал инициатором магистрального ГПТ. Тем не менее в двух случаях из четырех магистральное ГПТ было зарегистрировано сразу по всем восьми модулям столона (LHF = 23.1 мм), еще в одном случае ГПТ было в семи модулях столона (LHF = 21 мм) и только в одном случае – не более пяти модулей. Продолжительность однонаправленного ГПТ в отдельных модулях, принимая в расчет как насыщенные, так и ненасыщенные ГПТ, обычно составляла не более 2 мин, но в одном случае магистральное ГПТ длилось до 4 мин (строки 1:17–1:20), а компенсаторное ГПТ – до 8 мин (строки 0:30–0:37). Суммарное число модулей, охваченных магистральными ГПТ, не намного больше, чем компенсаторными ГПТ (табл. 1). Итак, в колонии № 1-8 (2016) насыщенные ГПТ начинались не в материнском побеге (в трех случаях из четырех), но наиболее протяженные ГПТ были сопряжены с исходом ГПТ из материнского побега.
Бланк 1. Колония № 8-6 (2016) представляла пример очень слабых ГПТ (рис. 5). Доля покоя составила в этой колонии 82.4% (табл. 1). За 1.5 ч наблюдений было зарегистрировано шесть коротких насыщенных ГПТ (< и >), что составляет всего 1.6% от всех учтенных модулей. Их протяженность не превышала двух модулей (LHF < 6 мм). Ненасыщенных (слабых) ГПТ (← и →) за все время было обнаружено 16% от всех модулей. В девяти случаях из восьмидесяти девяти регистраций они были противонаправленными, т.е. в пределах одного акта сканирования были зарегистрированы в противоположных направлениях. В трех случаях стрелки ненасыщенных ГПТ отходили от одного побега (← →) или были направлены к нему (→ ←). Это средние побеги в колонии. Наибольшая протяженность сплошного неразрывного ГПТ с учетом как насыщенных, так и ненасыщенных частицами течений составляла 11.4 мм (четыре модуля), а без учета ненасыщенных ГПТ – только два модуля. Продолжительность однонаправленного ГПТ, принимая в расчет как насыщенные, так и ненасыщенные ГПТ, составляла не более 1–2 мин, за одним исключением (до 3 мин). Периодичность ГПТ была не выражена. Суммарное число модулей, охваченных магистральными ГПТ, больше, чем компенсаторными (табл. 1).
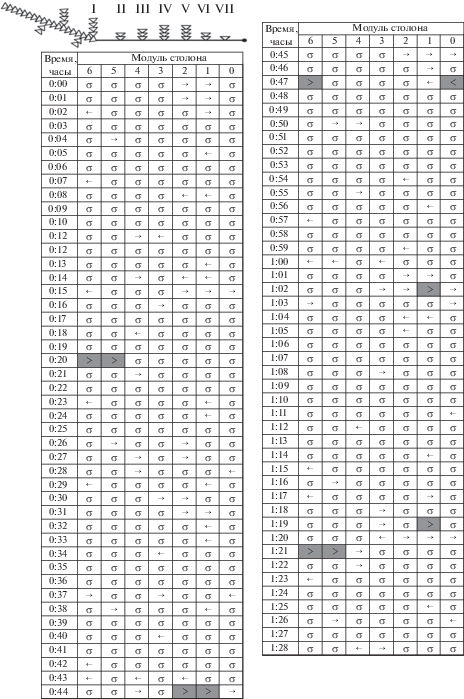
Бланк 11. Колония № 8-6 (2016) является примером неупорядоченных токов гидроплазмы (рис. 6). Это повторное визуальное сканирование ГПТ исследованной ранее колонии (бланк 1) через шесть дней после первого сканирования. Колония непосредственно перед сканированием была покормлена дозированно – прицельно гидрантам разных побегов, что повлияло на интенсивность ГПТ (табл. 1). Всего было проглочено 12 науплиев: из них шесть – гидрантами материнского побега; один науплиус был в ближайшем к материнскому побеге; три науплия – в среднем побеге (№ V), удаленном от материнского на четыре модуля столона; еще два науплия были в гидрантах предпоследнего побега – № VII (см. обозначения на верхней схеме рис. 6). В ближайшем к верхушке столона побеге еще не было гидрантов. В такой системе отсутствовал четко выраженный лидирующий побег, задающий ритмику магистральных течений. Каждый побег в колонии, включая материнский, генерировал непродолжительные (длительностью 2–3 мин) разнонаправленные течения, сталкивающиеся и гасящие друг друга, не образующие общий поток: например, модули 2–3 (0:05, 0:48 ч), 3–5 (0:30 ч). Некоторые побеги единовременно выдавливали в столон насыщенные (< >) разнонаправленные ГПТ: модули 1–2 (0:00 ч) и 3–4 (1:01 ч). Несмотря на то, что практически все побеги демонстрировали ритмику и периодичность в генерации ГПТ, общая по столону картина динамики токов выглядела случайной и хаотичной. Доля покоя составляла 67.7% (табл. 1). Однако даже в течение этого непродолжительного срока наблюдений активность ГПТ заметно снижалась, а фаза покоя увеличивалась. Так, для первых 45 мин регистрации доля покоя составляла 58.9%, а в последующие 45 мин была 75.2%. В первые 45 мин максимальная протяженность магистрального ГПТ даже в такой неупорядоченной системе достигала 15 мм (пять модулей столона). Во второй полупериод регистрации протяженность однонаправленных насыщенных ГПТ уменьшилась и не превышала трех модулей. Ни один из побегов не стал ведущим в формировании общеколониальных ГПТ. Суммарное число модулей, охваченных магистральными ГПТ, несколько меньше, чем компенсаторными (табл. 1).
Рис. 6.
Бланк 11. Колония № 8-6 (2016) через шесть дней после первого сканирования – пример распределительной системы с неупорядоченными токами гидроплазмы.
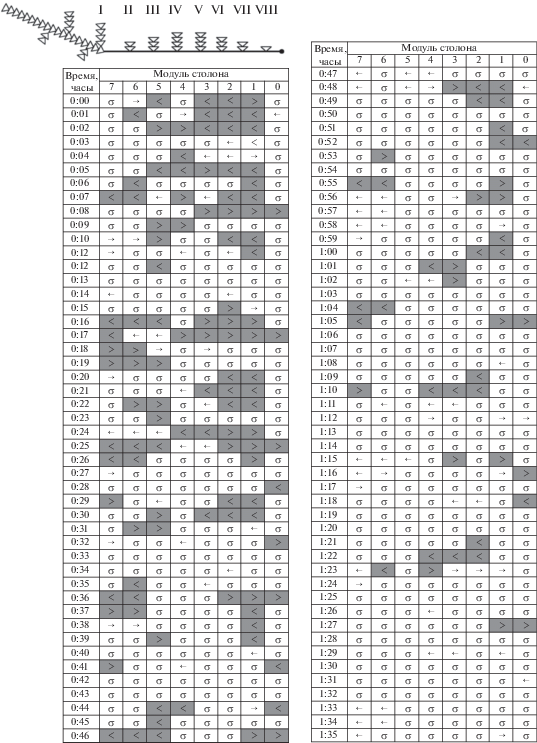
Бланк 4. Колония № 2-5 (2016), накормленная дозированно за 3 ч перед экспериментом, обладала чрезвычайно мощными ГПТ, проходящими через весь столон, т.е. около 28 мм (восемь модулей). Интенсивность таких течений не снижалась в течение всего периода регистрации (рис. 7). Колония получила 10 науплиев, из них пять достались гидрантам материнского побега, а остальные пять – побегам № IV, V и VII. Периоды покоя (σ) были непродолжительными, суммарно для всего периода регистрации фаза покоя составляла всего 26.9% (табл. 1). Однако движение потоков не было хаотичным. Магистральные течения покидали материнский побег и, дойдя за 2–4 мин до верхушки или расположенного вблизи верхушки побега (строчки 0:10–0:14, 0:27–0:31, 0:36–0:41, 0:45–0:49, 0:54–0:57, 1:03–1:07, 1:21–1:24 ч), как правило, сразу меняли направление, устремляясь мощным потоком к материнскому побегу, где безостановочно (строчки 0:08–0:09, 0:16–0:17, 0:26–0:27, 0:43–0:44, 0:53–0:54, 1:09–1:11, 1:28–1:29 ч) вновь обращались в противоположную сторону. Доля периода покоя для модуля столона, ближайшего к материнскому побегу, составляла 10.9%. Суммарное число модулей, охваченных магистральными ГПТ, больше, чем компенсаторными (табл. 1).
ГПТ очень частые, периодичность их возникновения составляла строго 9 мин. Всего за 1.5 ч наблюдений было зарегистрировано 11 магистральных ГПТ, берущих начало от материнского побега. Восемь из них достигли верхушки столона. Два ГПТ из 11 были протяженными: от материнского побега до верхушки столона без разрыва. В остальных случаях магистральное ГПТ было меньшей протяженности, достигая шестого побега (от материнского) и далее завершаясь течением от этого побега к верхушке столона. Такими же промежуточными “помпами” иногда были четвертый (строчки 0:49 и 1:18) и пятый побеги (строчки 0:31 и 0:50), хотя крошечный четвертый побег получил только одного науплиуса, а пятый побег вообще не получил пищи перед сканированием.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Главная задача нашего исследования заключалась в определении протяженности неразрывных течений гидроплазмы (ГПТ) в столоне колониального гидроида с пульсаторно-перистальтической распределительной системой. В ней перемещение гидроплазмы непостоянно. Взвешенные в гидроплазме частицы переносятся течением то в одном направлении, то в противоположном. Создается впечатление, что частицы колеблются на одном месте. Так и должно было бы быть, если перемещение гидроплазмы вызывается пульсациями многочисленных гидрантов и верхушек роста, что было отмечено и в ранних исследованиях (Hale, 1960), и в более поздних (Карлсен, Марфенин, 1976, 1984; Марфенин, 1985; Бурыкин, 2010). Тем не менее частицы пищи успешно переносятся на большие расстояния, соизмеримые с длиной столона колонии.
Фултон (Fulton, 1963) описал перистальтические сокращения в гидрантах, побегах и дистальных участках столонов колоний Cordylophora sp. Казалось бы, дистально направленные перистальтические сокращения ценосарка в столонах способны обеспечить дальний перенос частиц в гидроплазме, однако оставалось невыясненным, как взаимодействуют между собой перистальтические волны от разных пульсаторов. По нашим наблюдениям у D. pumila перистальтические волны сжатия и расширения ценосарка ограничены лишь непротяженным дистальным участком ценосарка вблизи верхушки роста; они могут быть направлены как дистально, так и проксимально. Следовательно, нет оснований считать, что перистальтика является основным способом перемещения частиц по ценосарку у колониальных гидроидов. Тогда как же достигается перемещение частиц через всю колонию от мест захвата добычи на периферию колонии к местам ее роста? Насколько закономерны протяженные ГПТ?
Ранее было показано (Косевич, 1991; Марфенин, 1993), что в линейных колониях Gonothyraea loveni, в которых науплиев A. salina давали гидрантам самого старого (первичного) побега, расположение растущих побегов зависело от количества получаемой пищи. При минимальном рационе (один науплиус в сутки) рос только самый дальний побег, ближайший к верхушке столона. При суточном рационе в пять науплиев росли четыре–пять побегов, ближайших к верхушке столона. При 10 экз./сут росли 10 побегов, и только при избыточном кормлении росли все побеги колонии (Косевич, 1991; Марфенин, 1993). Каким-то образом пища была доставлена на периферию колонии, причем быстро – в считанные часы, о чем можно было судить по изменению окраски тканей дистальной области столона. Такой же результат был получен и при радиоуглеродном исследовании сложноорганизованных колоний Pennaria tiarella с перистыми побегами (Rees et al., 1970).
Выяснение истинной протяженности магистральных и компенсаторных ГПТ необходимо для определения способа и эффективности функционирования распределительной системы. Эту задачу ранее пытались решить тремя способами: 1) непосредственным отслеживанием под микроскопом перемещения частиц по ценосаркальной трубке; 2) использованием радиоактивных или иных меток; 3) расчетным методом, используя для этого результаты непрерывной локализованной в одном месте видеосъемки или визуальной регистрации с хронометрированием.
Первый способ. Перемещение взвешенных в гидроплазме частиц по столону действительно можно отслеживать под микроскопом (Карлсен, Марфенин, 1976, 1984) при условии, что колонии выращены на прозрачном субстрате, сам ценосарк достаточно прозрачен, а перисарк свободен от обрастателей. Вырастить колониальных гидроидов на стеклах вполне возможно. Впервые этот метод был описан Кровеллом (Crowell, 1957), а в дальнейшем слегка модифицирован (Бурыкин и др., 1984). Прозрачности перисарка также можно достичь, если выращивать колонии в темноте. Намного сложнее добиться прозрачности ценосарка по всей его длине. Обычно более-менее прозрачными остаются лишь участки ценосарка вблизи верхушек роста. Чем дальше от зоны роста, тем темнее становится ценосарк. Это мешает отслеживанию перемещений отдельных частиц внутри ценосаркальной трубки. Нельзя быть до конца уверенными, что после очередного побега в поле зрения осталась та же самая частица, перемещение которой отслеживает исследователь. Для того, чтобы надежно различать отдельные частицы, они должны быть крупными, а ценосарк прозрачным, что получается, если выращивать колонии при ограниченном кормлении, а перед регистрацией давать дозированно науплиев определенным побегам. Кроме того, для подобных исследований нужны короткие молодые колонии, а не колонии с длинными столонами, у которых прозрачность столона снижается по мере удаления от его верхушки. Тем не менее при некотором навыке и повторности можно с уверенностью установить маршрут отдельных частиц по ценосарку столона. Так, было установлено, что максимальная протяженность перемещения частицы в столоне D. pumila составляет 50 мм (Карлсен, Марфенин, 1976, 1984), а у более коротких колоний – 22 мм (Марфенин, Дементьев, 2017). И в том и в другом случае максимальная протяженность пробега отдельных частиц была ограничена размерами столона колонии.
Второй способ. Направление и скорость миграции пищи внутри колонии отслеживаются по распространению метки, полученной гидрантами с пищей. Исследование, выполненное с использованием радиоактивной метки, было проведено на колониальном гидроиде Pennaria tiarella11. Таким способом было установлено, что радиоуглеродная метка уже через полчаса после кормления оказывается в ценосарке и поступает в зоны роста (Rees et al., 1970). Аналогичное исследование, выполненное на стелющемся колониальном гидроиде Podocoryna carnea M. Sars, 1846 с использованием искусственной пищи с флуоресцентной меткой, показало, что пища через несколько часов после кормления концентрировалась в некоторых верхушках столонов (Buss et al., 2015).
Третий способ. Расчетный метод определения протяженности ГПТ был предложен недавно (Marfenin, 2015). Этот метод основан на данных стационарной микровидеосъемки перемещения частиц в выбранном для этого месте ценосарка в пределах участка длиной около 0.7 мм обычно в середине модуля столона. Суммируя пошаговые значения скоростей движения частиц, полученные ежесекундно, можно для однонаправленного ГПТ определить расстояние, которое проходит взвешенная в гидроплазме частица от начала течения до его окончания. На практике удобнее определять скорость движения частиц в ценосаркальном канале не ежесекундно, а с интервалом в 30 с. В таком случае суммируются произведения значений скоростей на 30 с. Согласно результатам, полученным этим методом, протяженность даже мощных ГПТ не превышает двух модулей столона, т.е. не более 7 мм (Марфенин, Дементьев, 2017).
Результаты, полученные описанными выше способами, оказались несовпадающими. Согласно визуальному отслеживанию наибольшая протяженность перемещения частиц в столонах достигает 50 мм (Карлсен, Марфенин, 1976, 1984; Бурыкин, 2006), а согласно расчетному способу наибольшая протяженность строго фиксируемого при помощи цейтраферной микровидеосъемки ГПТ составляет менее 7 мм. Это противоречие актуализирует задачу определения протяженности неразрывного ГПТ. Тем более что Ю.Б. Бурыкин (2006, 2008, 2010) по результатам собственных исследований формирования ГПТ в маленьких колониях гидроида Gonothyraea loveni предложил альтернативную гипотезу, согласно которой регулярных протяженных ГПТ в столонах у колониальных гидроидов нет, движение гидроплазмы между пульсаторами происходит хаотически, а перемещение пищи к дистальным концам столонов осуществляется как бы “по эстафете” и в значительной мере случайно, от одного модуля к другому.
В данной статье представлены результаты четвертого способа изучения протяженности ГПТ. Использованная нами ежеминутная посегментная регистрация направления движения частиц в каждом модуле столона в течение 1.5 ч для каждой колонии позволила описать пространственную картину динамики гидроплазмы в столонах небольших линейных колоний D. pumila, чего раньше не удавалось достичь другими способами. Мы регистрировали не перемещение отдельных частиц вдоль всего столона, а направление движения любых частиц последовательно в каждом модуле (посегментно) за 10–20 с сканирования, т.е. почти одномоментно. При таком учете можно точно определить протяженность всего ГПТ в целом по сумме длин модулей, в которых были зарегистрированы однонаправленные движения частиц, не придавая значения маршруту отдельных частиц.
Как и следовало ожидать, система течений в столонах разнообразна. В представленных нами результатах имеются примеры как отсутствия протяженных ГПТ (рис. 5) или неупорядоченных коротких ГПТ (рис. 6), так и четкой периодики и направления мощных ГПТ (рис. 3).
Неупорядоченность коротких ГПТ и невыраженность протяженных течений вполне естественны для модульных организмов, у которых отсутствует централизованное управление, а гидроплазма приводится в движение не одним пропульсаторным органом, а множеством пульсаторов (Марфенин, 1993, 2016). Интерес представляют не разнообразные варианты неупорядоченности ГПТ, а примеры регулярных протяженных однонаправленных ГПТ, которые представлены на бланках 16 и 7.
На бланке 16 видно, что время от времени в колонии D. pumila возникают сплошные сквозные ГПТ, сопоставимые по протяженности с длиной всего столона (рис. 3). Оказалось, что сквозные ГПТ от одного до другого конца столона могут возникать менее чем за 1 мин, хотя чаще формируются на протяжении 2–4 мин, постепенно распространяясь вперед.
Означает ли это, что при возникновении сквозных магистральных ГПТ, частицы проходят за 1 мин путь, равный длине всего столона? Особенность метода сканирования заключается в том, что по каждому модулю определяется лишь направление ГПТ, а не скорость движения частиц. По результатам сканирования обычно нельзя определенно ответить на вопрос: прошел ли фронт ГПТ всю дистанцию от основания материнского побега до верхушки столона. Ясно лишь, что частицы во всех модулях движутся в одном направлении. Но с какой скоростью? И какой путь они проходят за однонаправленное ГПТ? Теоретически возможны, по крайней мере, две версии ответа на эти вопросы.
По первой версии частицы в ценосаркальной трубке могут одновременно сдвинуться в одну сторону при условии увеличения давления с одной стороны ценосарка, например, в результате сжатия гидрантов в материнском побеге или ослабления давления в противоположной части ценосарка, т.е. прекращения сжатия ценосарка вблизи верхушки роста или при ослаблении гидрантов. Протяженность такого ГПТ может быть значительной, но длина пути, пройденного частицами, намного меньшей. В движение приходит гидроплазма по всему столону, но сдвигается она ненамного, лишь на величину рабочего объема растягиваемого пульсатора – дистального участка столона. После того, как иссякнет течение из материнского побега, ГПТ в столоне остановится, а затем гидроплазма начнет двигаться в противоположном направлении, так как растянувшийся дистальный участок ценосарка столона начнет сжиматься.
По второй версии при тех же условиях частицы, теоретически, могут пройти весь путь от материнского побега до верхушки роста столона при условии, что емкость принимающего пульсатора столь велика, что превышает объем полости всего ценосарка столона. Однако проведенные ранее расчеты опровергают такое предположение. Рабочие объемы пульсаторов составляют не больше двух–трех модулей столона (Марфенин, Дементьев, 2017).
Тем не менее, в некоторых случаях на бланках представлены явные доказательства того, что за минуту частицы прошли расстояние, соответствующее длине нескольких модулей столона. Например, когда зарегистрировано, что ненасыщенное ГПТ сменяется в модулях насыщенным, можно предполагать, что частицы действительно преодолели расстояние, равное совокупной длине этих модулей, за время, не превышающее 1 мин. Такие примеры мы находим на описанных выше бланках. Например, на бланке 7 (рис. 4, строчки 0:37 и 0:38). Здесь компенсаторное ненасыщенное ГПТ в пределах трех модулей за 1 мин сменяется магистральным насыщенным ГПТ. Такой же переход на бланке 11 (рис. 6, строчки 0:24 и 0:25), на бланке 4 (рис. 7, строчки 0:14 и 0:15, 0:41 и 0:42).
Подобные примеры дают нам основание считать, что менее чем за 1 мин фронт ГПТ может распространиться на расстояние, равное длине, по меньшей мере, трех модулей, т.е. в среднем на 8–9 мм. В таком случае скорость фронта ГПТ превышает 150 мкм/с. Этот результат согласуется с данными прямого учета скорости ГПТ по движению взвешенных в гидроплазме частиц, полученными ранее (Карлсен, Марфенин, 1984). Было показано, что средняя скорость ГПТ в некоторых столонах D. pumila равна 200 мкм/с, а максимальная – 800 мкм/с. При таких скоростях за 1 мин частицы могут пройти внутри ценосаркальной трубки от 12 до 48 мм, что и было зарегистрировано этими авторами при непосредственном визуальном отслеживании перемещения отдельных частиц по столону.
Однако надо учитывать, что перемещение частиц по столону возможно не только вследствие поступления гидроплазмы из одного источника (одного побега), но и при последовательном поступлении непротяженных ГПТ в побеги по пути продвижения. Опубликованные ранее результаты наблюдений свидетельствуют, что в ответ на поступление ГПТ в побег сразу же образуется исходящее ГПТ, которое оказывается очередным двигателем магистрального ГПТ. Так, подключаясь поочередно, промежуточные побеги участвуют в продвижении гидроплазмы, в результате чего магистральное ГПТ оказывается неограниченно протяженным (Марфенин, 1985, 1993). Поясним это на следующем примере. Из побега, получившего пищу, выходит в столон насыщенное частицами ГПТ. Продвижение этого фрагмента ГПТ по столону возможно лишь при условии, что находящийся перед ним в столоне “пустой” объем гидроплазмы должен также сместиться. Если смещение ограничится только зоной столона, то эффект получится таким же, как описан в первой версии. Иной результат получится в том случае, если “пустой” объем гидроплазмы продвинется не только по столону, но зайдет в следующий побег, что неизбежно приведет к ответному сжатию ценосарка в нем и выдавливанию того же “пустого” объема гидроплазмы обратно в столон. Однако между входом ГПТ в побег и выходом из него проходит некоторое незначительное время. Было замечено, что исходящая из очередного побега порция пустой гидроплазмы прошла по столону дальше, а “пустой” объем гидроплазмы, выдавленной обратно в столон, оказался в хвосте продвинутой вперед порции гидроплазмы с частицами пищи, т.е. эти объемы гидроплазмы меняются местами. Инициатива в проталкивании ГПТ переходит ко второму побегу, после чего частицы пищи продолжают движение по столону, а “пустой” объем гидроплазмы перед ними загоняется в следующий побег. И снова расширение ценосарка провоцирует его сжатие и, следовательно, генерацию исходящего ГПТ, а “пустой” объем гидроплазмы, который сначала был перед объемом, насыщенным частицами, оказывается позади него. Как в игре в “пятнашки”. Такая модель однонаправленного непрерывного ГПТ полностью снимала противоречие между тем, что частицы могут пройти весь путь от материнского побега до верхушки столона за один акт однонаправленного перемещения, и тем, что протяженность локального ГПТ, регистрируемая видеосъемкой в любом месте столона, оказывается значительно меньше протяженности всего столона.
Данные визуального сканирования ГПТ, приведенные на рис. 3–7, дают примеры, подтверждающие эту теорию. На бланке 16 (рис. 3) видно, что в арьергарде ГПТ возникают зоны покоя (строчки 0:18–0:20, 1:28–1:30). Следовательно, двигателем ГПТ в это время являлся уже не материнский побег, а один из промежуточных побегов: дистально от него ГПТ есть, а проксимально – нет.
Еще ярче пример на том же бланке, когда от промежуточного побега идут противоположно направленные ГПТ. Так, на строчке 0:56 от побега № V отходят два ненасыщенных ГПТ (рис. 3), которые пришли на смену насыщенному однонаправленному магистральному ГПТ на строчке 0:55. Побег № V, а за ним № VI стали главными двигателями магистрального ГПТ.
В то время как фронт ГПТ приближается к верхушке столона, задняя граница ГПТ отдаляется от побега, из которого оно изначально вышло. Если первоначально магистральное ГПТ начиналось от первого побега, то спустя минуту – уже от третьего побега, а еще через минуту – от четвертого (рис. 3, строчки 0:18–0:20). Такая последовательность смены “толкателей” – задних побегов, из которых выходит течение, формирующее магистральное ГПТ в столоне, – повторяется в приведенном примере четыре раза из пяти.
Во многих случаях методом посегментного сканирования можно не только определить заднюю границу магистрального или компенсаторного ГПТ, но и переднюю, т.е. выделить продвижение фронта ГПТ вперед по столону. На бланке 16 наиболее ярко это проявилось на строчках 0:52–0:55 (рис. 3). Если ГПТ есть в одном модуле и отсутствует в следующем, значит, гидроплазма поступает в побег, расположенный на границе этих модулей. В указанном примере приемником гидроплазмы были последовательно побеги № II, V, VII и затем верхушка столона.
Через 1–2 мин после поступления ГПТ в побег происходит инверсия, и течение выходит из побега, что может быть зарегистрировано методом визуального сканирования в тех случаях, когда на смену покоя (σ) или ненасыщенного ГПТ (→) приходит насыщенное ГПТ (>), как, например, представлено на строчках 0:15–0:19 (рис. 3).
Следовательно, перемещение частиц из одного конца столона до другого не соответствует модели двух пульсаторов, соединенных трубкой. Пульсаторы могут поочередно подключаться к “прокачиванию” ГПТ. Это скорее соответствует теории участия промежуточных побегов в генерации протяженного насыщенного частицами ГПТ (Марфенин, 1985, 1993).
Получается, что транспортировка частиц пищи по столону на значительные расстояния, сопоставимые с длиной всего столона колонии, возможна за одно или несколько последовательных магистральных ГПТ, т.е. менее чем за 1 ч. Однонаправленность транзита частиц усиливается благодаря тому, что в компенсаторных ГПТ их оказывается меньше, чем в магистральных. Этот факт был установлен почти во всех сеансах сканирования ГПТ. Несмотря на то, что частицы движутся по столону попеременно в две противоположные стороны, они все равно преимущественно смещаются к верхушке столона. Но это тема другой статьи.
ЗАКЛЮЧЕНИЕ
Согласно результатам визуального ежеминутного сканирования, неразрывное однонаправленное ГПТ может соответствовать по протяженности расстоянию от основания материнского побега до верхушки столона – в приведенных примерах до восьми модулей столона (27 мм). Это еще не означает, что такое расстояние проходят частицы за однонаправленное ГПТ, но в ряде случаев этим методом было установлено, что частицы в гидроплазме могут пройти расстояние до 9 мм за интервал менее 1 мин. Кроме того, получено подтверждение теории последовательного участия промежуточных побегов в продвижении ГПТ по столону. Согласно этой теории, изложенной ранее (Марфенин, 1985, 1993), возможно неограниченное однонаправленное перемещение частиц пищи при согласованной реакции побегов на проходящее под ними в столоне ГПТ.
Список литературы
Бурыкин Ю.Б., 2006. Механизмы формирования течений гидроплазмы в колониях гидроидных полипов // Мат-лы X науч. конф. ББС МГУ. М.: Изд-во “Гриф и К”. С. 96–99.
Бурыкин Ю.Б., 2008. Формирование распределительной системы в процессе развития колоний гидроидных полипов // Онтогенез. Т. 39. № 3. С. 212–221.
Бурыкин Ю.Б., 2010. Основы функционирования распределительной системы колониальных гидроидов // Онтогенез. Т. 41. № 4. С. 300–311.
Бурыкин Ю.Б., 2013. Эстафетный способ перемещения гидроплазмы в колониях гидроидных полипов // Онтогенез. Т. 44. № 2. С. 115–125.
Бурыкин Ю.Б., 2015. Функционирование распределительной системы у колониального гидроида Dynamena pumila (L., 1758) // Вестн. МГУ. Сер. 16. Биол. Вып. 3. С. 44–48.
Бурыкин Ю.Б., Марфенин Н.Н., Карлсен А.Г., 1984. Опыт содержания морского колониального гидроида Dynamena pumila (L.) в лабораторных условиях // Биол. науки. № 1. С. 102–106.
Дементьев В.С., Марфенин Н.Н., 2017. Функциональная морфология столонов у колониального гидроида Gonothyraea loveni (Allman, 1859): рост, пульсации и перемещение гидроплазмы // Изучение, рациональное использование и охрана природных ресурсов Белого моря. СПб.: ЗИН РАН. С. 66–69.
Дементьев В.С., Марфенин Н.Н., 2018. Влияние опреснения на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 79. № 5. С. 376–392.
Дементьев В.С., Марфенин Н.Н., 2019. Воздействие температуры на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 80. № 1. С. 22–42.
Карлсен А.Г., Марфенин Н.Н., 1976. Упорядоченность перемещения гидроплазмы в колониальном гидроиде Dynamena pumila (L.) (Thecaphora, Sertulariidae) // Журн. общ. биологии. Т. 37. № 6. С. 917–923.
Карлсен А.Г., Марфенин Н.Н., 1984. Перемещение гидроплазмы в колонии у гидроидов на примере Dynamena pumila (L.) и некоторых других видов гидроидов // Журн. общ. биологии. Т. 45. № 5. С. 670–680.
Косевич И.А., 1991. Сравнение функционирования верхушек роста побегов и столонов в колонии Obelia loveni (Allm.) (Hydrozoa, Campanulariidae) // Вестн. МГУ. Сер.16. Биол. № 2. С. 44–52.
Марфенин Н.Н., 1980. Метод картирования пространственной организации колониальных Hydrozoa и его значение при изучении частей колонии // Теоретическое и практическое значение кишечнополостных. Л.: ЗИН АН СССР. С. 66–69.
Марфенин Н.Н., 1985. Функционирование распределительной системы пульсаторно-перистальтического типа у колониальных гидроидов // Журн. общ. биологии. Т. 46. № 2. С. 153–164.
Марфенин Н.Н., 1988. Функционирование распределительной системы в колонии у колониальных гидроидов: новый метод и факты // Губки и книдарии. Современное состояние и перспективы исследований. Л.: Зоол. ин-т АН СССР. С.103–111.
Марфенин Н.Н., 1993. Феномен колониальности. М.: Изд-во МГУ. 237 с.
Марфенин Н.Н., 2002. Нецентрализованная саморегуляция целостности колониальных организмов // Журн. общ. биологии. Т. 63. № 1. С. 26–39.
Марфенин Н.Н., 2016. Децентрализованный организм на примере колониальных гидроидов // Биосфера. Т. 8. № 3. С. 315–337.
Марфенин Н.Н., Дементьев В.С., 2017. Парадокс протяженных течений гидроплазмы в колониальном гидроиде Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 78. № 4. С. 3–20.
Марфенин Н.Н., Дементьев В.С., 2018а. Продольные пульсации столона у колониального гидроида Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 79. № 2. С. 85–96.
Марфенин Н.Н., Дементьев В.С., 2018б. Рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) в проточной и непроточной кюветах // Журн. общ. биологии. Т. 79. № 2. С. 97–107.
Blackstone N.W., 1996. Gastrovascular flow and colony development in two colonial hydroids // Biol. Bull. V. 190. № 1. P. 56–68.
Buss L.W., Anderson C.P., Perry E.K., Buss E.D., Bolton E.W., 2015. Nutrient distribution and absorption in the colonial hydroid Podocoryna carnea is sequentially diffusive and directional // PLoS One. V. 10. № 9. P. 1–36. https://doi.org/10.1371/journal.pone.0136814
Crowell S., 1957. Differential responses of growth zones to nutritive level, age, and temperature in the colonial hydroid Campanularia // J. Exp. Zool. V. 134. № 1. P. 63–90.
Dudgeon S.R., Wagner A., Vaisnys J.R., Buss J.W., 1999. Dynamics of gastrovascular circulation in the hydrozoan Podocoryne carnea: The one-polyp case // Biol. Bull. V. 196. P. 1–17.
Fulton C., 1963. Rhythmic movements in Cordilophora // J. Cell. Comp. Physiol. V. 61. № 1. P. 39–51.
Hale L.J., 1960. Contractility and hydroplasmic movements in the hydroid Clytia johnstoni // Quart. J. Microscop. Sci. V. 101. № 3. P. 339–350.
Harmata K.L., Parrin A.P., Morrison P., Bross L.S., Blackstone N.W., 2013. Quantitative measures of gastrovascular flow in octocorals and hydroids: Towards a comparative biology of transport systems in cnidarians // Invertebr. Biol. V.132. № 4. P. 291–304.
Marfenin N.N., 2015. A new method for studying the transport system in colonial hydroids // Hydrobiologia. V. 759. № 1. P. 133–146.
Marfenin N.N., Dementyev V.S., 2017. Functional morphology of hydrozoan stolons: Stolonal growth, contractility, and hydroplasmic movement in Gonothyraea loveni (Allman, 1859) // Mar. Biol. Res. V. 13. № 5. P. 521–537.
Parrin A.P., Netherton S.E., Bross L.S., McFadden C.S., Blackstone N.W., 2010. Circulation of fluids in the gastrovascular system of a stoloniferan octocoral // Biol. Bull. V. 219. № 2. P. 112–121.
Rees J., Davis L.V., Lenhoff H.M., 1970. Paths and rates of food distribution in the colonial hydroid Pennaria // Comp. Biochem. Physiol. V. 34. № 2. P. 309–316.
Wagner A., Dudgeon S., Vaisnys J.R., Buss L.W., 1998. Nonlinear oscillations in polyps of the colonial hydroid Podocoryne carnea // Naturwissenschaften. V. 85. P. 117–120.
Winkle D.H., van, Blackstone N.W., 1997. Video microscopical measures of gastrovascular flow in colonial hydroids // Invertebr. Biol. V. 116. № 1. P. 6–16.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии