

Океанология, 2020, T. 60, № 2, стр. 233-243
Моделирование и реконструкция биотического потока энергии в экосистеме Белого моря на основе балансового принципа
В. В. Бульон 1, *, В. Я. Бергер 1
1 Зоологический институт РАН
Санкт-Петербург, Россия
* E-mail: vboulion@mail.ru
Поступила в редакцию 13.04.2019
После доработки 29.10.2019
Принята к публикации 16.12.2019
Аннотация
Представлены результаты анализа масс–балансовой модели, имитирующей биотический поток энергии в экосистеме Белого моря. Модель предназначена для прогнозирования годовой продукции автотрофных организмов, редуцентов и консументов разного порядка, включая рыб и морских млекопитающих. Результаты анализа модели даются в табличной форме и включают в себя биотический баланс моря, т.е. биомассу и продукцию ключевых групп гидробионтов и значения энергетических потоков, текущих по трофическим цепям по типу “жертва–хищник”. При прогнозировании продукции бактериопланктона учитывалось, что бактерии ассимилируют не только продукцию фитопланктона, но и аллохтонное РОВ. Продукция морских млекопитающих (белух), прогнозируемая моделью, близка по значению к продукции, рассчитанной по данным авиаучета численности животных и количеству потребленной ими пищи. Согласно модели продукция рыбного сообщества ≈1.2 ккал/(м2 год). По статистическим данным продукция рыб примерно в 5 раз меньше прогнозируемой величины. Это связано, по-видимому, с существенной недооценкой запаса рыб в Белом море, на основании которого рассчитывалась продукция всего рыбного сообщества. Согласно прогнозу, годовой вылов рыбы в Белом море мог бы составлять 4 кг/га (0.4 ккал/м2 год), что очень близко к тому, что имело место в далеком прошлом, около 150 лет назад (по архивным данным). В настоящее время промысел рыб в Белом море резко сократился, либо плохо учитывается.
Знания о механизме функционирования водных экосистем нуждаются в обобщении, которое может быть исполнено путем моделирования трофических связей и биотического потока энергии при строгом соблюдении первого закона термодинамики. Биотические и абиотические факторы через систему соподчиненных связей формируют поток энергии от фотосинтезирующих организмов и редуцентов к консументам разного порядка. Такой системный подход в сочетании с балансовым принципом был использован при создании масс–балансовой модели Белого моря. Первоначально модель предназначалась для прогнозирования скорости биотических процессов в экосистемах озерного типа. Она успешно прошла тестирование по эмпирическим данным для озер Белоруссии, Карелии, Карельского перешейка и водохранилищ Дальнего Востока [10, 12–14, 23]. Данная работа – первый опыт применения модели для оценки биологической продуктивности Белого моря, которое, будучи внутренним морем, по морфологическим и гидрологическим параметрам имеет некоторое сходство с крупными озерными экосистемами: сравнительно слабый водообмен с Северным Ледовитым океаном, сток рек играет большую роль в формировании гидрохимического режима [33]. Результаты модели сопоставлены с опубликованными материалами по Белому морю [3, 4, 26–28].
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Модель представляет собой трофическую пирамиду (рис. 1), в основании которой – первичные продуценты (фитопланктон Bphp, микрофитобентос Bphb, макрофиты Bmac и эпифиты Bepi). Следующий трофический уровень представлен редуцентами (бактериопланктоном Bbcp и бактериобентосом Bbcb), за которыми следуют консументы 1-го и 2-го порядка (нехищный Bhzo и хищный Bpzo зоопланктон и зообентос Bzbe) и консументы 3-го порядка (планктоноядные Bplf и бентосоядными рыбами Bbef). В вершине трофической пирамиды – хищные рыбы Bpsf и морские млекопитающие Bmam. Связи между компонентами модели изображены одинарными стрелками, каналы потока энергии – двойными стрелками. В качестве входных параметров модели используются следующие независимые переменные.
Рис. 1.
Схема модели для прогнозирования биомассы и продукции ключевых групп гидробионтов (Lat, Tem, Dmean, Dmax, Sec, PO, TP – входные параметры). Объяснения символов в тексте.
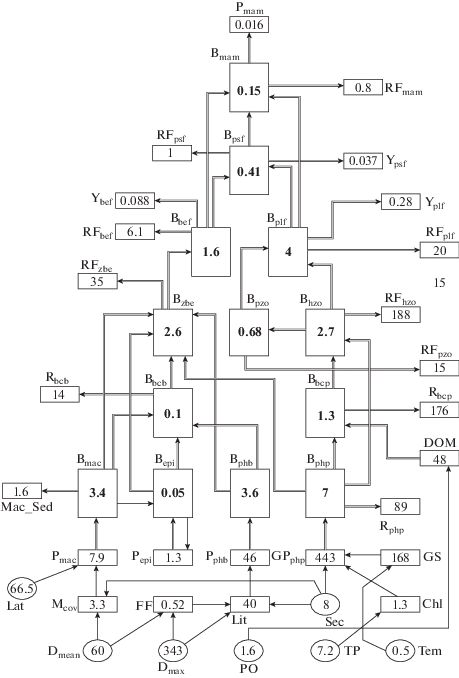
Абиотические факторы: Lat – географическая широта, °с.ш.; Tem – среднегодовая температура воздуха, °C; Dmean и Dmax – средняя и максимальная глубина моря, м; Sec – прозрачность воды по диску Секки, м; PO – перманганатная окисляемость воды, мг O2/л; TP – содержание в воде общего фосфора, мкг/л (табл. 1).
Таблица 1.
Средние значения входных абиотических параметров модели экосистемы Белого моря: [2, 4, 16, 24, 25]
| Географическая широта, Lat | 66.5 °с.ш. |
| Среднегодовая температура воздуха, Tem | 0.5°C |
| Средняя глубина, Dmean | 60 м |
| Максимальная глубина, Dmax | 343 м |
| Прозрачность воды, Sec | 8 м |
| Перманганатная окисляемость воды, PO | 1.6 мг O2/л |
| Содержание в воде общего фосфора, TP | 7.2 мкг/л |
Биотические факторы: Скорость оборота биомассы гидробионтов, суточное ассимиляционное число хлорофилла “a” фитопланктона (табл. 2).
Таблица 2.
Средние значения входных биотических параметров модели: [1, 4, 5, 17, 19, 20, 27, 30, 35, 41, 42, 50, 52]
| Трофическое звено | Скорость оборота биомассы, сут–1 | Эффективность роста | ||
|---|---|---|---|---|
| Фитопланктон | Vphp | 0.30 | Ephp | 0.8 |
| Макрофиты | Vmac | 0.014 | – | – |
| Эпифиты | Vepi | 0.15 | – | – |
| Микрофитобентос | Vphb | 0.075 | – | – |
| Бактериопланктон | Vbcp | 0.4 | Ebcp | 0.33 |
| Бактериобентос | Vbcb | 0.4 | Ebcb | 0.33 |
| Нехищный зоопланктон | Vhzo | 0.08 | Ehzo | 0.16 |
| Хищный зоопланктон | Vpzo | 0.06 | Epzo | 0.32 |
| Зообентос | Vzbe | 0.015 | Ezbe | 0.16 |
| Планктоноядные рыбы | Vplf | 0.0025 | Eplf | 0.08 |
| Бентосоядные рыбы | Vbef | 0.0020 | Ebef | 0.08 |
| Хищные рыбы | Vpsf | 0.0016 | Epsf | 0.1 |
| Млекопитающие | Vmam | 0.0006 | Emam | 0.019 |
| DAN* 25 | ||||
Эффективность использования водными организмами потребленной энергии на рост (эффективность роста) (табл. 2).
Коэффициенты распределения продукции гидробионтов между потребителями (табл. 3).
Таблица 3.
Коэффициенты распределения (DC) продукции гидробионтов между потребителями: [4, 6, 35, 41]
| Трофические цепи | Коэффициенты распределения | |
|---|---|---|
| Макрофиты: | ||
| бактериобентос | DCmac_bcb | 0.3 |
| зообентос | DCmac_zbe | 0.5 |
| донные отложения | 1 – DCmac_bcb – DCmac_zbe | 0.2 |
| Эпифиты: | ||
| бактериобентос | DCepi_bcb | 0.4 |
| зообентос | 1 – DCepi_bcb | 0.6 |
| Микрофитобентос: | ||
| бактериобентос | DCphb_bcb | 0.4 |
| зообентос | 1 – DCphb_bcb | 0.6 |
| Нехищный зоопланктон: | ||
| хищный зоопланктон | DChzo_pzo | 0.6 |
| планктоноядные рыбы | 1 – DChzo_pzo | 0.4 |
| Планктоноядные рыбы: | ||
| хищные рыбы | DCplf_psf | 0.5 |
| млекопитающие | DCplf_mam | 0.33 |
| вылов* | 1 – DCplf_psf – DCplf_mam | 0.17 |
| Бентосоядные рыбы: | ||
| хищные рыбы | DCbef_psf | 0.5 |
| млекопитающие | DCbef_mam | 0.33 |
| вылов* | 1 – DCbef_psf – DCbef_mam | 0.17 |
| Хищные рыбы | ||
| млекопитающие | DCpsf_mam | 0.67 |
| вылов* | 1 – DCpsf_mam | 0.33 |
Уравнения, используемые в модельном алгоритме:
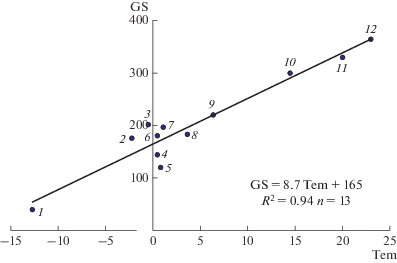
GS = 8.7 Tem + 164 – длительность вегетационного сезона, дни (рис. 2).
Рис. 2.
Зависимость длительности вегетационного сезона (GS, дни) от среднегодовой температуры воздуха (Tem, °C). Озеро на о-ве Хейса в архипелаге Земля Франца-Иосифа (1), Зейское (2) и Бурейское (3) водохранилища, озера Кривое в Северной Карелии (4) и Зеленецкое на Кольском п-ве (5), Белое море (6), Богучанское водохранилище (7), озера Красное на Карельском перешейке (8), Нарочь в Беларуси (9), Инкити в Грузии (10), Киннерет в Израиле (11), Джордж в Уганде и Кодаи в Индии (12) [12, 13, 23, 48, 51].
Chl = 0.103 TP1.29 – концентрация хлорофилла a, мкг/л [9].
Mcov = 25 (Sec/Dmean ) – площадь зеркала водоема, занимаемая макрофитами, % [8].
Lit = (1 – ((Dmax – 2 Sec)/(Dmax + 2 Sec EXP(3 – – FF1.5)))(0.5/FF)) 100 – отношение площади литоральной зоны к площади зеркала водоема, % [40].
FF = 3Dmean/Dmax – фактор формы котловины водоема.
Модельный алгоритм:
Фитопланктон
Bphp = EphpGPphp/(VphpGS) – биомасса фитопланктона, ккал/м2.
GPphp = DAN Chl Sec × GS/100 – валовая продукция фитопланктона, ккал/(м2 год).
Rphp = GPphp (1 – Ephp) – потери энергии фитопланктоном при дыхании, ккал/(м2 год).
Макрофиты
Bmac = Pmac/(VmacGS) – биомасса макрофитов, ккал/м2.
Pmac = 10 ^ (2.21 + 1.08 LOG10(Mcov) – ‒ 0.49 (90/(90 – Lat))) – чистая продукция макрофитов, ккал/(м2 год) [8].
Mac_Sed = Pmac (1 – DCmac_bcb – DCmac_zbe) – продукция макрофитов, захороненная в донных отложениях, ккал/(м2 год)
Микрофитобентос
Bphb = Pphb/(VphbGS) – продукция микрофитобентоса, ккал/м2.
Pphb = EphpGPphp × 0.32 Lit – чистая продукция микрофитобентоса, ккал/(м2 год), как функция продукции фитопланктона и относительной площади литоральной зоны [8].
Эпифиты
Bepi = 0.015 Bmac – биомасса эпифитов, ккал/м2. Принято, что биомасса эпифитов составляет ≈1.5% биомассы макрофитов [8].
Pepi = 0.015 PmacVepi/Vmac – чистая продукция эпифитов, ккал/(м2 год).
Бактериопланктон
Bbcp = Ebcp (Php_Bcp + DOM)/(VbcpGS) – биомасса бактериопланктона, ккал/м2.
Php_Bcp = (0.2/Ebcp)Pphp – продукция фитопланктона, ассимилированная бактериопланктоном, ккал/(м2 год), где Pphp = BphpVphpGS. Принято, что ≈20% чистой продукции фитопланктона включается в бактериальную продукцию [7].
DOM = ((0.3 PO) × 0.001DmeanGS) 10 – аллохтонное РОВ, ассимилированное бактериопланктоном, ккал/(м2 год). Принято, что константа скорости ассимиляции бактериопланктоном гуминовых соединений составляет ≈0.001 сут–1 [15, 36–38, 49]. Концентрация в воде гуминовых веществ, мг C/л, оценивалась по перманганатной окисляемости воды [31, 32].
Rbcp = (DOM + Php_Bcp)(1 – Ebcp) – потери энергии бактериопланктоном при дыхании, ккал/(м2 год).
Бактериобентос
Bbcb = Ebcb(Mac_Bcb + Epi_Bcb + Phb_Bcb)/ (VbcbGS) – биомасса бактериобентоса, ккал/м2.
Mac_Bcb = DCmac_bcbPmac – продукция макрофитов, ассимилированная бактериобентосом, ккал/(м2 год).
Epi_Bcb = DCepi_bcbPepi – продукция эпифитов, ассимилированная бактериобентосом, ккал/(м2 год).
Phb_Bcb = DCphb_bcbPphb – продукция микрофитобентоса, ассимилированная бактериобентосом, ккал/(м2 год).
Rbcb = (Epi_Bcb + Mac_Bcb + Phb_Bcb)(1 – Ebcb) – потери энергии бактериобентосом при дыхании, ккал/(м2 год).
Нехищный зоопланктон
Bhzo = Ehzo(Php_Hzo + Bcp_Hzo)/(VhzoGS) – биомасса нехищного зоопланктона, ккал/м2.
Php_Hzo = Pphp – Php_Bcp – Php_Zbe – продукция фитопланктона, потребленная нехищным зоопланктоном, ккал/(м2 год).
Bcp_Hzo = Pbcp – продукция бактериопланктона, потребленная нехищным зоопланктоном, ккал/(м2 год), где Pbcp = BbcpVbcpGS.
RFhzo = (Bcp_Hzo + Php_Hzo)(1 – Ehzo) – потери энергии нехищным зоопланктоном при дыхании и с неусвоенной пищей, ккал/(м2 год).
Хищный зоопланктон
Bpzo = EpzoHzo_Pzo/(VpzoGS) – биомасса хищного зоопланктона, ккал/м2.
Hzo_Pzo = DChzo_pzoPhzo – продукция нехищного зоопланктона, потребленная хищным зоопланктоном, ккал/(м2 год), где Phzo = BhzoVhzoGS.
RFpzo = Hzo_Pzo(1 – Epzo) – потери энергии хищным зоопланктоном при дыхании и с неусвоенной пищей, ккал/(м2 год).
Зообентос
Bzbe = Ezbe(Mac_Zbe + Epi_Zbe + Phb_Zbe + + Bcb_Zbe + Php_Zbe)/(VzbeGS) – биомасса зообентоса, ккал/м2.
Mac_Zbe = DCmac_zbePmac – продукция макрофитов, потребленная зообентосом, ккал/(м2 год).
Epi_Zbe = (1 – DCepi_bcb)Pepi – продукция эпифитов, потребленная зообентосом, ккал/(м2 год).
Phb_Zbe = (1 – DCphb_bcb)Pphb – продукция микрофитобентоса, потребленная зообентосом, ккал/(м2 год).
Php_Zbe = 0.1 BphpGS/Dmean – продукция фитопланктона, осевшая на дно и потребленная зообентосом, ккал/(м2 год). 0.1 – скорость седиментации фитопланктона, м/сут.
Bcb_Zbe = Pbcb – продукция бактериобентоса, потребленная зообентосом, ккал/(м2 год), где Pbcb = BbcbVbcbGS.
RFzbe = (Bcb_Zbe + Epi_Zbe + Mac_Zbe + + Phb_Zbe + Php_Zbe)(1 – Ezbe) – потери энергии зообентосом при дыхании и с неусвоенной пищей, ккал/(м2 год).
Планктоноядные рыбы
Bplf = Eplf(Hzo_Plf + Pzo_Plf)/(VplfGS) – биомасса планктоноядных рыб, ккал/м2.
Hzo_Plf = (1 – DChzo_pzo)BhzoVhzoGS – продукция нехищного зоопланктона, потребленная планктоноядными рыбами, ккал/(м2 год).
Pzo_Plf = Ppzo – продукция хищного зоопланктона, потребленная планктоноядными рыбами, ккал/(м2 год), где Ppzo = BpzoVpzoGS.
Yplf = (1 – DCplf_psf – DCplf_mam)Pplf – вылов человеком и (или) естественная смертность планктоноядных рыб, ккал/(м2 год), где Pplf = BplfVplf GS.
RFplf = (Hzo_Plf + Pzo_Plf)(1 – Eplf) – потери энергии планктоноядными рыбами при дыхании и с неусвоенной пищей, ккал/(м2 год).
Бентосоядные рыбы
Bbef = EbefZbeBef/(VbefGS) – биомасса бентосоядных рыб, ккал/м2.
Zbe_Bef = Pzbe – продукция зообентоса, потребленная бентосоядными рыбами, ккал/(м2 год), где Pzbe = BzbeVzbeGS.
Ybef = (1 – DCbef_psf – DCbef_mam)Pbef – вылов человеком и (или) естественная смертность бентосоядных рыб, ккал/(м2 год), где Pbef = BbefVbefGS.
RFbef = Zbe_Bef(1 – Ebef) – потери энергии бентосоядными рыбами при дыхании и с неусвоенной пищей, ккал/(м2 год).
Хищные рыбы
Bpsf = Epsf(Plf_Psf + Bef_Psf)/(VpsfGS) – биомасса хищных рыб, ккал/м2.
Plf_Psf = DCplf_psfPplf – продукция планктоноядных рыб, потребленная хищными рыбами, ккал/(м2 год).
Bef_Psf = DCbef_psfPbef – продукция бентосоядных рыб, потребленная хищными рыбами, ккал/(м2 год).
Ypsf = (1 – DCpsf_mam)Ppsf – вылов человеком и (или) естественная смертность хищных рыб, ккал/(м2 год), где Ppsf = BpsfVbefGS.
RFpsf = (Bef_Psf + Plf_Psf)(1 – Epsf) – потери энергии хищными рыбами при дыхании и с неусвоенной пищей, ккал/(м2 год).
Морские млекопитающие
Bmam = Emam(Plf_Mam + Bef_Mam + Psf_Mam)/ (VmamGS) – биомасса морских млекопитающих, ккал/м2.
Plf_Mam = DCplf_mamPplf – продукция планктоноядных рыб, потребленная морскими млекопитающими, ккал/(м2 год).
Bef_Mam = DCbef_mamPbef – продукция бентосоядных рыб, потребленная морскими млекопитающими, ккал/(м2 год).
Psf_Mam = DCpsf_mamPpsf – продукция хищных рыб, потребленная морскими млекопитающими, ккал/(м2 год).
Pmam = BmamVmamGS – продукция морских млекопитающих, ккал/(м2 год).
RFmam = (Plf_Mam + Bef_Mam + Psf_Mam)(1 – – Emam) – потери энергии морскими млекопитающими при дыхании и с неусвоенной пищей, ккал/(м2 год).
Биотические параметры V, E и DC, заданные как x-переменные, выведены на связанные с моделью панели, аналогичные табл. 2 и 3. Результаты анализа модели даются в табличной форме и включают в себя биотический баланс моря, т.е. биомассу и продукцию ключевых групп гидробионтов (табл. 4) и значения энергетических потоков, текущих по трофическим цепям по типу “жертва–хищник” (табл. 5).
Таблица 4.
Средние значения биомассы (В), продукции (Р), скорости оборота биомассы (Р/В) и отношения продукции к суммарной первичной продукции (P/Pprim) для организмов разных трофических групп Белого моря. Результаты моделирования
| Трофическое звено | B, ккал/м2 | Р, ккал/м2 за вегетационный сезон | P/B, за вегетационный сезон | P/Pprim, % |
|---|---|---|---|---|
| Фитопланктон | 7 | 354 | 51 | 87 |
| Макрофиты | 3.4 | 7.9 | 2.4 | 1.9 |
| Эпифиты | 0.05 | 1.3 | 25 | 0.31 |
| Микрофитобентос | 3.6 | 46 | 13 | 11 |
| Первичные продуценты | – | 409 | – | 100 |
| Бактериопланктон | 1.3 | 87 | 67 | 21 |
| Бактериобентос | 0.1 | 7 | 67 | 1.7 |
| Нехищный зоопланктон | 2.7 | 36 | 13 | 8.8 |
| Хищный зоопланктон | 0.68 | 6.9 | 10 | 1.7 |
| Сообщество зоопланктона | – | 21 | – | 5.1 |
| Зообентос | 2.6 | 6.6 | 2.5 | 1.6 |
| Планктоноядные рыбы | 4 | 1.7 | 0.42 | 0.42 |
| Бентосоядные рыбы | 1.6 | 0.53 | 0.34 | 0.13 |
| Хищные рыбы | 0.41 | 0.11 | 0.27 | 0.027 |
| Сообщество рыб | – | 1.2 | – | 0.27 |
| Морские млекопитающие | 0.15 | 0.016 | 0.1 | 0.0038 |
Таблица 5.
Распределение биотического потока энергии между компонентами трофической сети в экосистеме Белого моря. Результаты моделирования
| Трофические цепи | Поток, ккал/(м2 год) | Трофические цепи | Поток, ккал/(м2 год) |
|---|---|---|---|
| Фитопланктон: | Нехищный зоопланктон: | ||
| бактериопланктон | 215 | хищный зоопланктон | 23 |
| нехищный зоопланктон | 138 | планктоноядные рыбы | 14 |
| зообентос* | 2 | Хищный зоопланктон: | |
| Макрофиты: | планктоноядные рыбы | 6.9 | |
| бактериобентос | 2.4 | Зообентос: | |
| зообентос | 4 | бентосоядные рыбы | 6.6 |
| донные отложения | 1.6 | Планктоноядные рыбы: | |
| Эпифиты: | хищные рыбы | 0.85 | |
| бактериобентос | 0.51 | млекопитающие | 0.57 |
| зообентос | 0.76 | вылов** | 0.28 |
| Микрофитобентос: | Бентосоядные рыбы: | ||
| бактериобентос | 18 | хищные рыбы | 0.26 |
| зообентос | 27 | млекопитающие | 0.18 |
| Аллохтонное РОВ: | вылов** | 0.088 | |
| бактериопланктон | 87 | Хищные рыбы: | |
| Бактериопланктон: | млекопитающие | 0.074 | |
| нехищный зоопланктон | 48 | вылов** | 0.037 |
| Бактериобентос: | |||
| зообентос | 7 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На основании PO оценивалось содержание в воде растворенного органического вещества РОВ, поступающего в море извне, преимущественно, с речным стоком. По М.П. Максимовой [22], PO в Белом море изменяется по акватории и глубинам в узких границах – 1.4–2.0 при средней 1.6 мг О2/л. Для перехода величины PO к содержанию аллохтонного РОВ, представленного гуминовыми соединениями (в мг C/л), применялся коэффициент 0.3 (в соответствии с дыхательным коэффициентом 0.8).
По И.Ю. Потаповой и Ю.Л. Сластиной [26], содержание TP в море достаточно стабильно, в границах 12–18, в среднем 15 мкг/л. Соотношение масс TN и TP значительно выше 7, молярное соотношение >15 [11, 26], что свидетельствует о роли фосфора как фактора, лимитирующего продукцию фитопланктона. По В.В. Сапожникову и М.П. Метревели [29], если принять во внимание все формы азота, а также прямую фиксацию азота из атмосферы, то положение с азотом в морских водах не так катастрофично, как принято считать. Показано, что лимитирование роста водорослей фосфором может чередоваться ограничительной ролью азота, то есть вопрос о том, в каком из двух элементов (азоте или фосфоре) более нуждаются растительные организмы, не является корректным [45].
Содержание хлорофилла a в планктоне Белого моря обычно варьирует от 0.9 до 2 мкг/л при средней ≈1.3 мкг/л [21, 24, 34, 46], соотношение Chl/TP = 0, 11 ± 0.04. Такое соотношение может применяться для оценки Chl по TP только в условиях Белого моря. Зависимость концентрации Chl от содержания фосфора в северных морских водах, в том числе в Белом море демонстрируется на рис. 3. Видно, что на фоне линий регрессий, описывающих зависимость Chl от TP в природных водах [9, 39, 43, 47], значения TP и Chl для Белого моря неплохо согласуются между собой, хотя некоторое отклонение от линий регрессий имеет место.
Рис. 3.
Зависимость концентрации хлорофилла a от содержания взвешенного (1, 2) и общего фосфора (3) в морских водах. 1 – пролив Ермак, о-в Хейса, 2 – Баренцево море 69°16′–80°40′ с.ш. [9], 3 – Белое море [26]. Линии регрессии: сплошная – Chl по PP [9], пунктирная – Chl по TP [39].
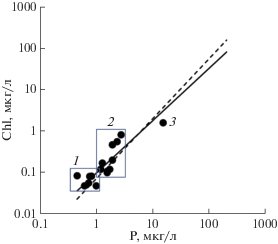
Обсуждаемая в данной работе модель позволяет реконструировать содержание TP по концентрации Chl в планктоне: если, согласно [21, 24, 34, 46], Chl = 1.3 мкг/л, то TP = 7.2 мкг/л. На этом основании нами было сделано предположение, что в Белом море только около половины TP находится в биологически доступной форме. Нелогично искать биологически активный фосфор в свободной растворенной форме, так как при дефиците фосфора и вследствие быстрого его оборота биологически доступный P практически весь ассимилируется фитопланктоном и таким образом трансформируется во взвешенную фракцию (PP). Такая точка зрения согласуется с моделью Смита [47], согласно которой существует вполне определенная достаточно тесная связь между PP и Chl, не зависящая от отношения TN/TP. Кроме того, TP ≈ PP при крайне высоких отношениях TN/TP. Значения всех входных абиотических параметров модели указаны в табл. 1.
В среднем за сезон суточная продукция фитопланктона варьирует от 115 до 670 мг С/(м2 сут) и составляет в среднем ≈ 250 мг С/(м2 сут) [4, 18]. Если глубина эвфотной зоне в Белом море 15–20 м [18] при Sec ≈ 8 м [16, 25], то скорость фотосинтеза на глубине с оптимальными световыми условиями, регистрируемая, как правило, у самой поверхности, может быть оценена как частное от деления суточной продукции фитопланктона на прозрачность воды по диску Секки, т.е. ≈30 мкг С/(л сут). Средняя концентрация Chl в поверхностных водах акватории Белого моря составляет, как показано выше, 1.3 мкг/л, следовательно, DAN ≈ 25 мг C/мг (табл. 2).
Исходя из значений входных биотических и абиотических параметров (табл. 1–3), суммарная первичная продукция Белого моря, включающая продукцию фитопланктона, макрофитов, эпифитов и микрофитобентоса, Pprim ≈ 410 ккал/(м2 год). Основная часть суммарной первичной продукции (87%) создается фитопланктоном. На долю микрофитобентоса приходятся 11%, на долю макрофитов и эпифитов лишь 2% суммарной первичной продукции. Согласно модели, величины чистой продукции фитопланктона и макрофитов очень близки к данным натурных наблюдений (рис. 4).
Рис. 4.
Сравнение эмпирических данных по продукции гидробионтов Белого моря [4] с результатами модели. 1 – морские млекопитающие, 2 – вылов рыбы (по архивным данным), 3 – продукция рыб, 4 – зообентоса, 5 – макрофитов, 6 – зоопланктона, 7 – фитопланктона, 8 – бактериопланктона (ккал/м2 год).
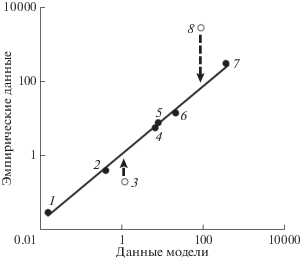
При прогнозировании продукции бактериопланктона учитывалось, что бактерии ассимилируют не только продукцию фитопланктона, но и аллохтонное РОВ. Бактерии, согласно модели, ассимилируют около 50 ккал/(м2 год) из фонда аллохтонного РОВ и 215 ккал/(м2 год) из фракции РОВ, продуцируемого фитопланктоном (табл. 5). Таким образом, общая ассимиляция РОВ бактериопланктоном ≈ 265 ккал/(м2 год).
По модели продукция бактериопланктона за счет ассимиляции аллохтонного РОВ и продукции фитопланктона ≈ 85 ккал/(м2 год). По натурным наблюдениям продукция бактерий почти в 30 раз превышает показания модели (рис. 4). Этот факт легко объясняется тем, что продукция бактерий измерялась в районе мидиевых хозяйств, где наблюдалось повышенное содержание РОВ и более высокая бактериальная активность по сравнению с пелагической зоной Белого моря. Если продукцию бактериопланктона, измеренную в узкой прибрежной зоне, занимающей около 3% площади моря, равномерно распределить по всей акватории моря, то ее величина приблизится к величине, прогнозируемой моделью, и практически не будет отличаться от результатов, полученных в серии экспедиций 2002–2006 гг. [28].
Модель позволяет раздельно оценивать продукцию нехищного и хищного зоопланктона (табл. 5). Продукция сообщества зоопланктона в целом (Pzpl) равна сумме продукций нехищного и хищных животных за вычетом рациона хищных животных. По модели Pzpl составляет 20 ккал/(м2 год). Эта величина близка к эмпирически установленной продукции зоопланктона – 14 ккал/(м2 год). Продукция зообентоса по данным модели ≈6.6 ккал/(м2 год). Она практически неотличима от данных, опубликованных В.Я. Бергером [3] (рис. 4).
Модель позволяет прогнозировать раздельно продукцию планктоноядных, бентосоядных, хищных рыб, а также продукцию сообщества рыб в целом (табл. 4). Согласно модели продукция рыбного сообщества ≈1.2 ккал/(м2 год). По статистическим данным [4], продукция рыб ≈0.2 ккал/(м2 год), т.е. примерно в 5 раз меньше прогнозируемой величины. Это связано, по-видимому, с существенной недооценкой запаса рыб в Белом море, на основании которого рассчитывалась продукция всего рыбного сообщества. Согласно прогнозу, годовой вылов рыбы в Белом море мог бы составлять 4 кг/га (0.4 ккал/м2 год), что очень близко к тому, что имело место в далеком прошлом, около 150 лет назад (по архивным данным [4]). В настоящее время промысел рыб в Белом море резко сократился, либо плохо учитывается.
Среди морских млекопитающих основной ихтиофаг – белуха. На втором месте по потреблению рыбы – кольчатая нерпа, морской заяц и, в малой мере, гренландский тюлень. Продукция морских млекопитающих, прогнозируемая моделью, близка по значению к продукции, рассчитанной косвенным способом – по данным авиаучета численности животных и количеству потребленной ими пищи (рис. 4).
По результатам модели, в основание которой заложен балансовый принцип, расходование продукции рыб протекает следующим образом: потребление морскими млекопитающими ≈0.82, хищными рыбами ≈1.1, оптимальный допустимый улов людьми в сумме с естественной смертностью рыб ≈0.4 ккал/(м2 год). По расчетам В.Я. Бергера [4], морские птицы используют лишь малую долю продукции зообентоса (≈1.4%) и рыб (≈1.2%), поэтому ролью птиц в биотическом балансе экосистемы Белого моря можно пренебречь.
ЗАКЛЮЧЕНИЕ
Модель экосистемы Белого моря, созданная с учетом накопленных за многие десятилетия знаний о механизме функционирования водных экосистем, сама становится “экосистемой”, которая подлежит исследованию. Анализ представленной здесь модели показал, что скорость потока энергии от первичных продуцентов и редуцентов к верхним трофическим уровням регулируется внешними факторами (продолжительностью вегетационного сезона через температуру, солнечной радиацией через ассимиляционное число, биогенными элементами и аллохтонным РОВ), т.е. “снизу”. Это согласуется с трофодинамической концепцией функционирования водных экосистем и с первым положением (эффектом “bottom-up”) распространенной концепции “трофического каскада” [44].
Преимущество модели – в ее доступности и прозрачности. Результаты ее анализа демонстрируются в виде биотического баланса водного объекта в наглядной и привычной для гидробиологов табличной форме. Модель может стать инструментом для реконструкции элементов пищевой сети и экспертной оценки трофического состояния экосистемы. В целом можно сделать вывод о том, что предложенная модель обладает достаточно высокой прогностической точностью и пригодна для составления биотических балансов не только озер, но и Белого моря, и, что важно, для взаимной проверки надежности эмпирических и предсказанных моделью данных.
Источник финансирования. Работа выполнена при поддержке гос. задания номер № АААА-А19-119020690091-0 “Исследования биологического разнообразия и механизмов воздействия антропогенных и естественных факторов на структурно-функциональную организацию экосистем континентальных водоемов. Систематизация биоразнообразия соленых озер и неполносоленых внутренних морей в зоне критической солености, изучение роли солоноватоводных видов в экосистемах”.
Список литературы
Алимов А.Ф. Элементы теории функционирования водных экосистем. СПб.: Наука, 2000. 147 с.
Белое море и его водосбор под влиянием климатических и антропогенных факторов / Под ред. Филатова Н.Н., Тержевика А.Ю. Петрозаводск: КарНЦ РАН, 2007. 349 с.
Бергер В.Я. Продукционный потенциал и промысловая бедность Белого моря // 30 лет морской биологической станции Санкт-Петербургского университета: итоги и перспективы. СПб: СПб ГУ, 2005. С. 7–24.
Бергер В.Я. Продукционный потенциал Белого моря. СПб: ЗИН РАН, 2007. 292 с.
Бергер В.Я., Бульон В.В. Оценка запасов рыб Белого моря с помощью экосистемного анализа // Вопросы промысловой океанологии. 2012. Вып. 9. № 1. С. 171–191.
Бульон В.В. Зависимость рыбопродуктивности водоемов от первичной продукции // Сборник научных трудов ГосНИОРХ. 1983. Вып. 196. С. 3–11.
Бульон В.В. Внеклеточная продукция фитопланктона и методы ее исследования // Гидробиол. журнал. 1988. Т. 24. № 3. С. 64–73.
Бульон В.В. Вклад основных групп автотрофных организмов в первичную продукцию водоемов // Водные ресурсы. 2004. Т. 31. № 1. С. 98–108.
Бульон В.В. Новый взгляд на парадигму фосфорного контроля в лимнологии // Успехи современной биологии. 2016. Т. 136. № 3. С. 311-318.
Бульон В.В., Сиротский С.Е. Биологическая продуктивность Богучанского водохранилища: моделирование и прогноз // Изв. РАН. 2015. Сер. биологическая. № 4. С. 431–440.
Водные ресурсы Европейского Севера России: итоги и перспективы исследований / Под ред. Филатова Н.Н. и др. Петрозаводск: КарНЦ РАН, 2006. 536 с.
Гидроэкологический мониторинг зоны влияния Бурейского гидроузла / Под ред. Сиротского С.Е. Хабаровск: ИВЭП ДВО РАН, 2007. 273 с.
Гидроэкологический мониторинг зоны влияния Зейского гидроузла / Под ред. Сиротского С.Е. Хабаровск: ИВЭП ДВО РАН, 2010. 354 с.
Динамика биологического разнообразия и биоресурсов континентальных водоемов / Под ред. Алимова А.Ф., Голубкова С.М. СПб.: Наука, 2012. 369 с.
Драчев С.М. Борьба с загрязнением рек, озер и водохранилищ промышленными и бытовыми стоками. М.: Наука, 1964. 274 с.
Жилинский А.А. Крайний Север европейской России. Петроград: СЗОПС, 1919. 296 с.
Иванова М.Б. Продукция планктонных ракообразных. Л.: ЗИН АН СССР, 1985. 222 с.
Ильяш Л.В., Радченко И.Г., Кузнецов Л.Л. и др. Пространственная вариабельность состава, обилия и продукции фитопланктона Белого моря в конце лета // Океанология. 2011. Т. 51. № 1. С. 24–32.
Инкина Г.А. Скорость потребления кислорода бактериопланктоном // Экспериментальные и полевые исследования биологических основ продуктивности озер. Л.: ЗИН АН СССР, 1979. С. 103–120.
Коржев В.А., Забавников В.Б. Оценка численности гренландского тюленя беломорской популяции в условиях недостатка биологической информации // Тр. ВНИРО. 2017. Т. 168. С. 105–116.
Кравчишина М.Д., Буренков В.И., Копелевич О.В. и др. Новые данные о пространственно-временной изменчивости концентрации хлорофилла а в Белом море // Докл. РАН. 2013. Т. 448. № 3. С. 342–348.
Максимова М.П. Органический углерод и окисляемость в водах Белого моря // Изв. Карельского и Кольского филиалов АН СССР. 1959. № 1. С. 71–74.
Многолетние изменения биологических сообществ мезотрофного озера в условиях климатических флуктуаций и эвтрофирования / Под ред. Трифоновой И.С. СПб.: Из-во ЛЕМА, 2009. 246 с.
Мордасова Н.В. Косвенная оценка продуктивности вод по содержанию хлорофилла // Тр. ВНИРО. 2014. Т. 152. С. 41–56.
Морской энциклопедический словарь. Т. 1 / Под ред. акад. Исанина Н.Н. Л.: Судостроение, 1991. 504 с.
Потапова И.Ю., Сластина Ю.Л. Распределение хлорофилла и биогенных элементов в воде Онежского залива Белого моря // Материалы IV Школы-конференции молодых ученых с международным участием. Петрозаводск: КарНЦ РАН, 2011. С. 71–74.
Примаков И.М., Бергер В.Я. Продукция планктонных ракообразных в Белом море // Биология моря. 2007. Т. 33. № 5. С. 356–360.
Саввичев А.С., Русанов И.М., Захарова Е.Е. и др. Микробные процессы циклов углерода и серы в Белом море // Микробиология. 2008. Т. 77. № 6. С. 823–838.
Сапожников В.В., Метревели М.П. Стехиометрическая модель органического вещества – основа количественного изучения продукционно-деструкционных процессов в океане // Тр. ВНИРО. 2015. Т. 155. С. 135–145.
Семенченко В.П. Консументы и их роль в экосистемах: Курс лекций. Минск: БГУ, 2004. 95 с.
Скопинцев Б.А. Органическое вещество в природных водах (водный гумус). Л.: Гидрометеоиздат, 1950. 290 с.
Скопинцев Б.А., Гончарова И.А. Использование значений отношений различных показателей органического вещества природных вод для его качественной оценки // Современные проблемы региональной и прикладной гидрохимии. Л.: Гидрометеоиздат, 1987. С. 95–117.
Стунжас П.А., Бородкин С.О. Гидрохимические критерии обмена поверхностных и глубинных вод Белого моря // Океанология. 2004. Т. 44. № 2. С. 189–198.
Шевченко В.П., Здоровеннов Р.Э., Кравчишина М.Д. и др. Системные исследования Белого моря в период летней межени 2009 г. в рейсе научно-исследовательского судна “Эколог” // Океанология. 2010. Т. 50. № 4. С. 666–670.
Экологическая система Нарочанских озер / Под ред. Винберга Г.Г. Минск: Из-во Университетское, 1985. 303 с.
Baпo N., Moraп M.A., Hodson R.E. Bacterial utilization of dissolved humic substances from a freshwater swamp // Aquat. Microb. Ecol. 1997. V. 12. № 3. P. 233–238.
Berggren M., Laudon H., Jansson M. Bacterial utilization of imported organic material in three small nested humic lakes // Verh. Internat. Verein. Limnol. 2010. V. 30. № 9. P. 1393–1396.
Bussmann I. Bacterial utilization of humic substances from the Arctic Ocean // Aquat. Microb. Ecol. 1999. V. 19. № 6. P. 37–45.
Dillon P.J., Rigler F.H The phosphorus-chlorophyll relationship in lakes // Limnol. Oceanogr. 1974. V. 19. № 5. P. 767–773.
Håkanson L. Water pollution – methods and criteria to rank, model and remediate chemical threats to aquatic ecosystems. Leiden: Backhuys Publishers, 1999. 299 p.
Håkanson L., Boulion V.V. The lake foodweb – modelling predation and abiotic/biotic interactions. Leiden: Backhuys Publ., 2002. 344 p.
Håkanson L., Boulion V.V. Modelling production and biomasses of zoobenthos in lakes // Aquatic Ecology. 2003. V. 37. № 3. P. 277–306.
Jones J.R., Bachmann R.W. Prediction of phosphorus and chlorophyll levels in lakes // J. Water Poll. Control Fed. 1976. V. 48. № 9. P. 2176–2182.
Kitchell J.F., Carpenter S.R. Cascading trophic interactions // The trophic cascade in lakes. Cambridge: University press, 1993. P. 1–14.
Lewis W.M., Wurtsbaugh W.A. Control of lacustrine phytoplankton by nutrients: erosion of the phosphorus paradigm // Internat. Rev. Hydrobiol. 2008. V. 93. № 4/5. P. 446–465.
Mordasova N.V. Chlorophyll in the White Sea // ICES J. Mar. Sci. 1999. Sup. 56. P. 215–218.
Smith V.H. The nitrogen and phosphorus dependency of algal biomass in lakes: An empirical and theoretical analysis // Limnol. Oceanogr. 1982. V. 27. № 6. P. 1101–1112.
The functioning of freshwater ecosystems / Eds. Kajak Z., Hillbricht-Ilkowska A. Cambridge: Univ. Press, 1980. 918 p.
Tulonen T. Role of allochthonous and autochthonous dissolved organic matter (DOM) as a carbon source for bacterioplankton in boreal humic lakes. Helsinki: University, 2004. 32 p.
www.ecoportal.info/kolchataya-nerpa
www.pogodaiklimat.ru
www.rgo.ru/bely-kit-beluha
Дополнительные материалы отсутствуют.