

Океанология, 2020, T. 60, № 3, стр. 381-392
Принципы биономического районирования береговой зоны и шельфа Мирового океана
К. М. Петров *
Санкт-Петербурский государственный университет
Санкт-Петербург, Россия
* E-mail: k.petrov@spbu.ru
Поступила в редакцию 30.07.2018
После доработки 18.07.2019
Принята к публикации 16.12.2019
Аннотация
Обсуждаются принципы создания иерархической системы единиц районирования береговой зоны и шельфа Мирового океана по биономическим (экосистемным) признакам. Решение ряда научных и прикладных задач требует районирования экорегионов (морских бассейнов) на региональном и топологическом уровнях. Предлагается система единиц, учитывающая зональные, глубинные и азональные особенности экорегионов. Основной исходной единицей биономического районирования является подводный ландшафт. В качестве примера районирования на топологическом и региональном уровнях рассматривается морфологическое строение ландшафтов трех округов подобласти Япономорское прибережье Сахалинской орогенной области экорегиона Японское море. Иерархическая система единиц биономического районирования позволяет выявлять ландшафты-аналоги. Прикладное значение выделения которых состоит в возможности прогнозировать интродукцию полезных видов и предусматривать опасность инвазии вредных видов.
ВВЕДЕНИЕ
Биогеографическое районирование океана предполагает наряду с биотическими (флористическими и фаунистическими) признаками использование биономических (экосистемных) признаков. В статье обсуждаются принципы создания иерархической системы единиц районирования по биономическим признакам.
1. Большие морские экосистемы. В конце XX– начале XXI веков было предложено понятие “Large Marine Ecosystems”, т.е. “Большие морские экосистемы” (БМЭ) [14, 15, 18]. Согласно определению БМЭ – это большие акватории, характеризующиеся своими особенностями батиметрии, гидрографии, биогеографии, продуктивности, экологии и трофически взаимосвязанными популяциями. На основе представлений о БМЭ сотрудниками Американского института биологических наук разработаны принципы районирования береговой зоны и шельфа Мирового океана. Выделяются 12 царств, 62 провинции и 232 экорегиона [19]. Царства характеризуются уникальной эволюционной историей, сходством гидробионтов высокого таксономического ранга и эндемичными таксонами на уровне семейств и родов. Ведущие факторы формирования царств – изоляция, связь с температурой воды, взаимодействие пелагических и донных форм жизни. Внутри царств обособляются провинции. Они характеризуются своеобразием биоты со своей эволюционной историей. Эндемизм поддерживается преимущественно на видовом уровне. Многие отличительные черты биоты – следствие исторической изоляции и влияния абиотических факторов, которые часто контролируют границы провинций. Эти факторы включают особенности геоморфологии (характер берегов и шельфа, изолированные острова, внутренние моря), гидрологии (температура, соленость, течения, апвеллинги, господствующие ветры, динамика ледового покрова) и гидрохимии (соленость, снабжение питательными веществами). Внутри провинций выделяются экорегионы (морские бассейны). Каждый экорегион отличается своеобразными чертами геоморфологического строения, гидрологических условий и доминированием определенных сообществ гидробионтов.
В Северном полушарии выделяются царства: Арктическое, Умеренное Северо-Атлантическое, Умеренное Северо-Пацифическое, Тропическое Атлантическое, Западное Индо-Пацифическое, Центральное Индо-Пацифическое, Восточное Индо-Пацифическое, Тропическое Восточно-Пацифическое. В Умеренном Северо-Пацифическом царстве у берегов России располагается Холодно-умеренная Западно-Пацифическая провинция, в которой выделяются экорегионы: Охотское море, Камчатский шельф и берег, Японское море и др. (рис. 1).
Рис. 1.
Экорегионы Холодно-умеренной Западно-Пацифической провинции Умеренного Северо-Пацифического царства [19]: 45 – Охотское море, 46 – Камчатский шельф и берег, 49 – Японское море.

Система единиц районирования БМЭ отражает наиболее крупные глобальные особенности жизни в океане. Оценка биоразнообразия, продуктивности, а также использование и охрана биологических ресурсов требуют разработки более детальной иерархической системы единиц биономического районирования экорегионов на региональном и топологическом уровнях.
2. Иерархическая система единиц биономического районирования. Биономические особенности береговой зоны и шельфа Мирового океана отражают влияние географической зональности, вертикальной поясности и характера геолого-геоморфологического строения на распределение сообществ гидробионтов.
Широтная зональность. Общепринятой системы природных зон в океане не существует. Большинство исследователей [1, 2, 6 ] принимают во внимание, прежде всего, океанические фронты, между которыми располагаются зоны, отличающиеся своими водными массами, интенсивностью природных процессов, химической активностью и биологической продуктивностью. Однако следует иметь в виду, что границы гидрологических фронтов в океане разделены, как правило, не линейными рубежами, а переходными полосами, положение которых меняется во времени. Поэтому поиски критериев, которые более всего соответствовали бы положению зональных границ в океане – задача весьма актуальная. В качестве общей модели географической зональности в океане принято выделять по два пояса – холодный и умеренный в каждом полушарии и разделяющий их теплый пояс [5]. Они опоясывают Землю, включая поверхностную толщу океана и прибрежные мелководья до глубин, подверженных сезонным колебаниям температур.
В начале XX века Сетчелл [17], исследуя географические закономерности распределения подводной растительности, выделил стенотермные группировки морских донных водорослей, характеризующиеся приуроченностью к водам, температурный режим которых различается на 5–10°С. Биологическое объяснение существованию стенотермных групп водорослей объяснила Перестенко [7], она установила, что индивидуальному развитию и жизненным циклам макрофитов свойственны температурные градиенты, которым соответствуют значения 0, 5, 10, 15, 20, 25°С. Эти термические границы предлагается использовать для выделения инварианта зонального подразделения Мирового океана. В Северном и Южном полушариях выделяются соответственно зоны арктическая и антарктическая, субарктическая и субантарктическая, холоднобореальная (бореальная) и холоднонотальная, теплобореальная (неморальная) и теплонатальная, а также расположенная между ними тропическая зона [11]. В пределах морских мелководий границы поверхностных и донных зон сливаются, на больших глубинах природные зоны однотипные с зонами, выделяемыми на поверхности океана, не прослеживаются.
Вертикальная поясность (зональность). Современная океанология придерживается хорошо выраженной четырехслойной стратификации океанических вод, выделяются: поверхностные (до 200 м), промежуточные (до 600–1000 м), глубинные (до 3500–4000 м) и придонные водные массы [13]. В местах соприкосновения этих вод с шельфом, материковым склоном и ложем океана предлагается выделять соответствующие ярусы: неритический, батиальный и абиссальный.
Неритический ярус, в свою очередь, целесообразно подразделять на пояса. Верхний пояс (береговая зона) находится в пределах волнового воздействия, ему присуща сезонная ритмика (прежде всего, прогрев в теплый период года) и освещенность достаточная для развития сообществ макрофитов. Средний пояс располагается ниже термоклина, волновая деятельность ослаблена, сезонная ритмика слабо выражена, освещенность низкая. Здесь встречаются единичные многоклеточные и некоторые одноклеточные водоросли. Нижний пояс соответствует перегибу профиля дна в сторону материкового склона, возрастает гидродинамическая активность, что приводит к размыву дна, образованию скульптурных форм рельефа и к увеличению разнообразия донных группировок.
Азональные особенности береговой зоны и шельфа. Важным фактором смены биономических условий в береговой зоне и на шельфе являются изменения характера рельефа и донных отложений. Ведущая роль в этом процессе принадлежит геолого-структурному строению окраины материков, новейшим и современным тектоническим движениям. В неоген–четвертичное время происходило развитие морфоструктур от планетарных до локальных, что определило основные черты рельефа морских окраин платформ, орогенов и островных дуг. Тектонические структуры контролируют рельеф побережья, контур береговой линии и приглубость подводного склона. Благодаря интенсивным дифференцированным унаследованным неотектоническим движениям происходит четкая локализация областей размыва дна и осадконакопления. Коллекторами денудационного материала являются региональные и локальные тектонические депрессии. В их пределах происходит формирование выровненного аккумулятивного рельефа. На месте унаследованных тектонических поднятий наблюдается отрицательная седиментация, обнажения коренных пород, происходит формирование скульптурного рельефа.
На материковых окраинах в соответствии со знаком неотектонических движений образуются два особых типа подводных ландшафтов со свойственными им донными группировками. В результате седиментационного выравнивания тектонических депрессий возникают аккумулятивные ландшафты с рыхлыми песчано-илистыми грунтами и сообществами закапывающихся в грунт гидробионтов. На месте структур, испытывающих поднятие, формируются абразионные ландшафты, характеризующиеся обнажением коренных пород, пересеченным рельефом и сообществами обрастателей. Широкое распространение имеет абразионно-аккумулятивный тип ландшафтов морских мелководий. Здесь донные природные комплексы (ДПК), связанные с обнажением коренных пород и формированием скульптурного рельефа, сочетаются с ДПК, образованными скоплением рыхлых осадков различного состава и генезиса, слагающих разнообразные аккумулятивные формы рельефа или выровненные пространства дна.
Большое значение имеет соотношение простирания морфоструктур с береговой линией: продольное или поперечное. При продольном простирании структур, испытывающих поднятие – берега высокие с клифом и поясом скал в подводной части склона, профиль дна – приглубый. При поперечном простирании морфоструктур, испытывающих поднятие, формируются обширные мелководья, детали строения которых связаны с дифференцированным развитием положительных и отрицательных локальных структур. Это находит выражение в формировании сложной мозаичной структуры подводного ландшафта; профиль дна пересеченный: аккумулятивные формы чередуются с абразионными. В целом, геолого-геоморфологическое строение морского дна служит каркасом для установления системы азональных единиц биономического районирования. Предлагается выделять: морские бассейны, области и округа береговой зоны и шельфа.
3. Подводные ландшафты. Исследования береговой зоны и шельфа, опирающиеся на общую теорию ландшафтоведения, определились в середине XX века [4, 9]. Однако развитию подводного ландшафтоведения в известной мере препятствовало отсутствие понятийного аппарата, который бы естественно вошел в теорию океанологии. Прежде всего, необходимо признать, что ландшафт является общим понятием, применимым при исследовании объектов как на суше, так и на морском дне.
Разнообразие подводных ландшафтов определяется рядом факторов. Гидрологические условия связаны с климатом и метеорологическим режимом атмосферы, что определяет сезонный ритм природных процессов в береговой зоне. Проникновение солнечной радиации поддерживает фотосинтез фитопланктона и фитобентоса. Подвижность вод контролирует процессы абразии, литодинамики и аккумуляции осадков, а также способствует хорошей аэрации, притоку питательных веществ и разносу зачатков организмов. Разгрузка жидкого и твердого стока суши вызывает сильную изменчивость солености морских вод и приводит к обогащению биогенными и органическими веществами. Большое видовое разнообразие и богатство жизненных форм определяет высокую плотность заселения разнообразных экологических ниш.
Подводные ландшафты береговой зоны и шельфа несут отпечаток влияния плейстоценовой регрессии, с которой связаны реликтовые формы рельефа, наличие субаэральных отложений и дизюнкция ареалов гидробионтов, и голоценовой трансгрессии, определившей молодость подводных ландшафтов.
Характерные черты подводного ландшафта в верхнем поясе шельфа определяются следующими свойствами:
1. Ландшафт морского дна обособляется, как правило, на одной региональной морфоструктуре.
2. Каждому ландшафту свойствен определенный набор литологических разностей современных донных отложений или выходов коренных пород разного вещественного состава, контролирующих характер микро- и мезоформ подводного рельефа.
3. Повышенная гидродинамическая активность, однородный режим температуры и солености морских вод, освещенность достаточная для существования фитобентоса.
4. Разнообразие форм рельефа, грунтов, гидроклиматических обстановок определяют пестроту условий местообитания и соответственно разнообразие донных биоценозов.
Перечисленные свойства служат основанием для выделения системы морфологических единиц внутриландшафтной вертикальной и горизонтальной дифференциации. Морфологические донные природные комплексы (ДПК) занимают особое место в учении о подводных ландшафтах. Они являются непосредственным объектом исследований и картографирования. В результате анализа их закономерных пространственных сочетаний выделяются и сами ландшафты.
Морфологические единицы вертикальной дифференциации. Верхний пояс шельфа принято делить по вертикали на супралитораль, литораль и сублитораль. Глубже сублиторали еще в конце XIX века Чельман [16] выделял элитораль. Опыт подводных исследований подтверждает объективность выделения этой зоны: у открытых берегов на глубине 30–40 м на каменистых грунтах заросли макрофитов сменяются группировками сессильной фауны, на подводном склоне аккумулятивного типа активная литодинамика уступает место спокойному осадконакоплению, на илистой поверхности дна выражена светлая бактериальная пленка [8].
Внутри зон выделяются этажи и ступени. В литорали они отражают высоту и ритмы приливо-отливных явлений, в сублиторали – ослабление воздействия волн на дно и угасание освещенности.
Отметим таксономическое непостоянство рассматриваемой категории единиц. Строго говоря, вертикальные зоны являются единицами не морфологической, а региональной размерности. Не зоны выделяются внутри ландшафта, а целые ландшафты умещаются внутри зон. Это правило справедливо для сублиторали и элиторали. Самые верхние “земноводные” зоны сравнительно редко представляют арену формирования особых супралиторальных и литоральных ландшафтов. Например, таких как ландшафт ваттов Северного моря.
В верхних поясах неритического яруса в качестве вертикальных зон региональной размерности предлагается различать береговую зону в целом и элиторальную зону. В этом случае супралитораль, литораль и сублитораль будут морфологическими единицами вертикального расчленения подводных ландшафтов, относящихся к береговой зоне моря.
Морфологические единицы горизонтальной дифференциации. Основными единицами горизонтального расчленения береговой зоны и шельфа являются подводные угодья и фации. Их обособление происходит под влиянием местных изменений рельефа, грунта и гидрологических условий, что находит отражение в своеобразии донных группировок гидробионтов.
Подводные угодья – это мезоформы рельефа, формирование которых происходит под решающим влиянием волновых и седиментационных процессов. На месте абразии, где вскрываются коренные породы, характер угодий определяется вещественным составом материнских пород и особенностями их залегания. Границы подводных угодий абразионного типа совпадают с геологическими телами и тектоническими формами, выраженными в рельефе дна. Своеобразие подводных угодий в местах аккумуляции определяется литологией и мощностью отложений. Особые условия наблюдаются на участках активной литодинамики, где формируются разнообразные аккумулятивные формы. В спокойной гидродинамической обстановке на участках устойчивого осадконакопления происходит аккумулятивное выравнивание дна, формирование монотонного плоского рельефа. Свойства литогенной основы являются экологическим фактором, определяющим своеобразие жизненных форм бентоса. Хорошо известны группы организмов, в облике которых находят отражение приспособительные признаки, обусловленные образом жизни на каменистом, песчаном, илистом грунтах и т.п.
Подводные фации – это одна форма микрорельефа или один элемент мезорельефа (вершина, склон, подножие банки). Каждая фация расположена в определенном интервале глубин, сложена одной литологической разностью современных осадков или приурочена к однородному по петрографическому составу комплексу горных пород и занята одним биоценозом. Подводные фации являются наименьшими ДПК, они весьма динамичны в пространстве и времени. Внутренняя структура фаций может осложняться скульптурными элементами размером от нескольких сантиметров до метра. Они могут быть представлены мозаичностью грунтов, мелкими грядами, отпрепарированными в результате абразии, а также мозаичностью горизонтальных синузий бентоса и т.п.
Пестрая картина морфологической дифференциации береговой зоны моря может быть представлена мозаикой или рядами фаций. Мозаика фаций одного угодья представляет собой сочетание генетически разнородных ДПК, распространенных в пределах равных глубин. Ряд фаций одного угодья образуют сопряженную систему, в которой смена фаций с увеличением глубины, обусловлена ослаблением волновых процессов и угасанием освещенности. Вертикальные ряды фаций кладутся в основу выделения морфологических единиц вертикального расчленения подводных ландшафтов.
4. Биономическая структура экорегиона Японское море. Тихоокеанские моря у берегов России Спалдинг с соавторами [19] относят к Умеренному Северо-Пацифическому царству, к Холодно-умеренной Западно-Пацифической провинции, в которой выделяют экорегионы: Охотское море, Камчатский шельф и берег, Японское море и др. (см. рис. 1).
При описании биономической структуры береговой зоны и шельфа Японского моря на региональном и топологическом уровнях предлагается выделять единицы широтной зональности, вертикальной поясности и азональные единицы биогеографического районирования. Поверхностную толщу вод Японского моря принято относить к бореальной (холоднобореальной) и неморальной (теплобореальной) зонам. Шельф и береговая зона входят в неритический ярус. По азональным признакам в бассейне Японского моря на границе с Охотским морем выделяется Сахалинская орогенная область (рис. 2). Япономорское прибережье острова Сахалин рассматривается как подобласть, которая делится на три округа: Северный и Южный – продольных берегов вдоль западных крыльев антиклинальных зон и между ними Центральный округ (перешеек Поясок) – замедленных поднятий, осложненных брахиантиклинальными складками. Гидрологические особенности Северного и Центрального округов по сравнению с Южным определяются проникновением охлажденных вод с севера, вызывающих понижение летних температур на 3–5°С. В составе биоты присутствуют холодноводные сообщества, например, ассоциация Fucus evanescens + Pelvetia wrightii в литорали, что позволяет отнести береговую зону этих округов к бореальной зоне. Гидрологические особенности Южного округа определяются теплыми водами Цусимского течения. Бентос образован тепловодными формами, например, для литорали характерны асс. Sargassum miyabei и Corallina pilulifera (“коралиновые тротуары”), характерно участие иглокожих Patiria pectinifera, Strongylocentrotus nudus. Это позволяет береговую зону Южного округа отнести к неморальной зоне.
Рис. 2.
Орогенная область остров Сахалин. 1 – участки поднятия земной коры (а – интенсивного, б – замедленного); 2 – абразионные берега; 3 – подводные ландшафты абразионного типа; 4 – участки опускания земной коры (а – аккумулятивные берега с широкими песчаными пляжами, б – подводные ландшафты аккумулятивного типа); 5 – оси крупных тектонических структур, испытывающих поднятие.

В округах обособляются подводные ландшафты. Каждый подводный ландшафт – это наименьшая региональная единица, он не пересекается границами таксонов высших порядков и обладает уникальными особенностями морфологического строения. Анализ фрагмента ландшафтной карты западного побережья Южного Сахалина (рис. 3) показывает, что если следовать с севера на юг, то морфологическое строение береговой зоны и шельфа заметно меняется. Это служит основанием для выделения нескольких ландшафтов. Биономические особенности Центрального и Южного округов связаны с морфологическим строением следующих ландшафтов: в первом округе – Ильинским, во-втором – Холмско-Невельским.
Рис. 3.
Подводные ландшафты западного побережья Южного Сахалина [12]. Ландшафты: I – Ильинский, II – Мыса Слепиковского, III – Холмско-Невельский, IV – Лопатинский, V – Шебунинский. Морфология берегов: 1 – абразионные берега с активным клифом; 2 – абразионные берега с отмершим клифом, окаймленные низкой (5–8 м) аккумулятивной морской террасой; 3 – аккумулятивные берега с широким песчаным пляжем. Угодья подводного берегового склона: 4 – пояс скал: а) с хорошо выраженной осушкой, б) без осушки; 5 – песчаные равнины; 6 – реликтовое гравийно-галечное поле; 7 – бровка материковой отмели.
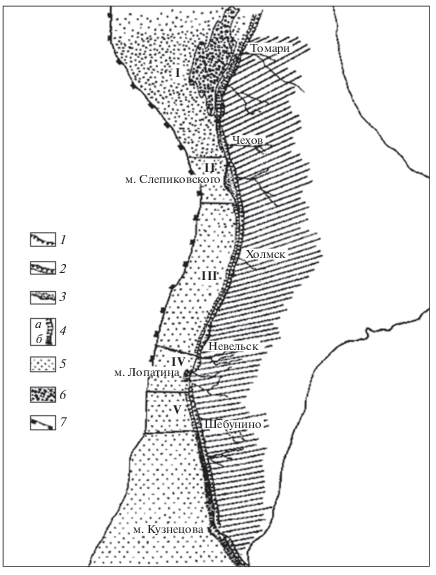
Ландшафт Ильинский простирается между м. Ламанон, расположенным за рамкой карты на севере, и м. Слепиковского на юге. Здесь бровка шельфа отступает далеко от берега, ширина мелководья у пос. Ильинского достигает 65 км. Ландшафт характеризуется развитием подводной абразионно-аккумулятивной платформы на месте простирающихся в море брахиантиклинальных складок. Основными элементами морфологического строения ландшафта, определяющими своеобразие биотопов и донных группировок, являются прибрежный пояс скал, каменистые банки вдали от берега, песчаные равнины, реликтовые поля гальки.
На границе Центрального и Южного округов расположен ландшафт м. Слепиковского – единственный для западных берегов Южного Сахалина пример ландшафта аккумулятивного типа. Он сформирован на месте синклинального прогиба, компенсированного накоплением толщи песчаных отложений. Береговая зона здесь резко сужается, изобата 40 м проходит на удалении 2–3 км от берега. Морфологическое строение ландшафта характеризуется доминированием одного биотопа песчаных равнин с характерной для него группировкой беспозвоночных.
Холмско-Невельский ландшафт простирается вдоль западного крыла Западно-Сахалинских гор от м. Слепиковского на севере до города Невельска на юге. Большие глубины котловины Японского моря прижимаются к берегу, сужая материковую отмель до 6–8 км. Подводный береговой склон абразионного типа. Характерными элементами морфологической структуры Холмско-Невельского ландшафта являются вытянутые вдоль берега и сменяющие друг друга по мере увеличения глубин угодья каменистой осушки, абразионного скульптурно-грядового склона, песчаной отсыпи. Южнее расположены ландшафты Лопатинский и Шебунинский, их описание не входит в задачу настоящей статьи.
Рассмотрим более подробно морфологическое строение Холмско-Невельского ландшафта. Основные элементы его морфологического строения отображены на аэрофотоснимке (рис. 4) и эколого-топологическом профиле (рис. 5).
Рис. 4.
Аэрофотоснимок ключевого участка Холмско-Невельского ландшафта, масштаб 1 : 5000. [3]. Элементы изображения. Побережье: 1 – береговой обрыв (отмерший клиф), 2 – низкая аккумулятивная морская терраса с расположенными на ней дорогой и домами. Угодье каменистой осушки: 3 – сообщество зостеры, 4 – выровненная поверхность с тонким слоем щебнистых отложений и разреженным покровом водорослей, 5 – внешний открытый прибою край каменистой осушки (темные пятна – сообщества морской травы филлоспадикс). Угодье подводного скульптурно-грядового берегового склона: 6 – крупные пятна и полосы интенсивного черного тона – сообщества крупных бурых водорослей ламинарии на пограничных грядах; 7 – полосчатость обусловленная грядовым микрорельефом и мелкая пятнистость, связанная с куртинами филлоспадикса, глубина до 5 м; 8 – неясные темные пятна, обусловленные зарослями красной водоросли одонталия, глубина до 10–15 м; 9 – желобообразное понижение, приуроченное к зоне разлома. Рисунок в виде сетки пересекающихся под разными углами линий, обусловлен трещинами, дробящими пласты пород на многочисленные блоки.

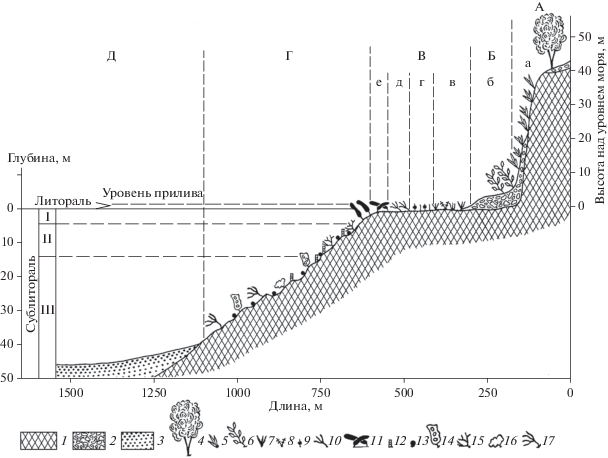
Рис. 5.
Вертикальная структура подводного Холмско-Невельского ландшафта. А, а – абразионный берег с отмершим клифом; Б, б – низкая аккумулятивная морская терраса; В – каменистая осушка: в – асс. Zostera marina, г – выровненная скульптурная поверхность с разреженным покровом водорослей (асс. Chondria dasyphylla + Sphaerotrichia divaricata), д – внешний край осушки с асс. морской травы Phyllospadix iwatensis, е – сообщество Laminaria japonica на грядах, окаймляющих каменистую осушку; Г – подводный скульптурно-грядовый склон; Д – песчаная отсыпь. Элементы вертикального подразделения сублиторальной зоны: I – верхний этаж, II – средний этаж, III – нижний этаж. Условные обозначения. Грунты: 1 – скальный (коренные породы), 2 – щебнистый, 3 – песчаный. Доминанты растительного покрова: 4 – каменная береза, 5 – курильский бамбук, 6 – сахалинская гречиха, 7 – Zostera marina, 8 – Sargassum miyabei, 9 – Sphaerotrichia divaricata, 10 – Phyllospadix iwatensis, 11 – Laminaria japonica, 12 – Bossiella cretacea, 13 – корковые известковые красные водоросли, 14 – Agarum cribrosum, 15 – Tichocarpus crinitus, 16 – Turnerella mertensiana, 17 – Dichloria viridis.
Угодье каменистой осушки представляет собой выровненную абразионно-аккумулятивную террасу шириной 150–300 м, выработанную в коренных породах. Поверхность террасы почти полностью лежит в интервале приливо-отливных вод. При отливе вода местами задерживается в понижениях рельефа (литоральных ваннах). Во время полной воды глубина моря не превышает 0.5 м. Значительная ширина осушки, небольшая глубина во время полной воды обусловливают спокойный гидродинамический режим в ее прибрежной части и усиление воздействия волн по мере удаления в сторону открытого моря. Соответственно меняется характер грунтов и растительности, что позволяет выделить три типа фаций.
В прибрежной части на песчано-илистом грунте почти сплошной пояс образует асс. Zostera marina. Здесь обитают креветки Pandalus latirostris. В местах, где слой песчаных отложений выклинивается, на каменистом грунте формируется асс. Sargassum miyabia. В средней части осушки волнение усиливается, рельеф мелко грядовый. На каменистом грунте формируется мозаичный растительный покров асс. Chondria dasyphylla + + Sphaerotrichia divaricata с участием зеленых водорослей Enteromorpha linza, Ulva fenestrata и бурых водорослей Punctaria plantaginea, Scytosiphon lomentaria. Внешняя, мористая часть осушки подвержена воздействию прибоя и характеризуется грядовым рельефом. Здесь характерна асс. морской травы Phyllospadix iwatensis, куртины которого образуют большие, диаметром в несколько метров, сомкнутые заросли. Отшлифованные прибоем каменистые поверхности обрастают корковыми известковыми красными водорослями. К вершинам пограничных гряд, отделяющих каменистую осушку от крутого абразионного склона, приурочены сообщества красной известковой кустистой водоросли Corallina pilulifera, плотные заросли которой именуются “кораллиновыми тротуарами”. С ними сочетаются типичные для прибойных местообитаний группировки красных водорослей Neorhodomela larix, Polysiphonia yendoi, Ceramium kondoi, Laurencia nipponica и др. Характерным компонентом биоценозов являются морские ежи Strongylocentrotus intermedius. Порой до нескольких десятков особей морских ежей на квадратный метр устилают межгрядовые понижения.
Угодье подводного скульптурно-грядового берегового склона прослеживается до глубины 20–30 (40) м. На этой глубине в прибрежных водах Южного Сахалина солнечная энергия, доступная для фотосинтеза, почти полностью поглощается, приблизительно на этой же глубине прекращается воздействие волн на дно. По этим признакам угодье подводного скульптурно-грядового склона относится к сублиторальной зоне. Описываемое угодье образовано флишеобразной толщей коренных пород. Различная устойчивость пластов к абразии обуславливает формирование резко пересеченного скульптурного рельефа (грядового, ступенчатого, глыбового и т.п.). Фации подводного берегового склона образуют вертикальный ряд, индицируемый последовательной сменой по глубинам сообществ макрофитобентоса [10].
Верхний этаж сублиторали в интервале глубин 0–5 м подразделяется на две ступени: верхнюю – от 0 до 2 м и нижнюю – от 2 до 5 м.
К верхней ступени приурочена серия пограничных гряд, отделяющих угодье каменистой осушки от угодья абразионного подводного берегового склона. Ширина комплекса пограничных гряд достигает нескольких десятков метров. Вершины гряд покрыты сообществом Laminaria japonica – Mesoalgosa – Corallina polulifera – Crustida (корковые известковые красные водоросли). Заросли ламинарии вытянуты вдоль берега узкими полосами длиной в сотни метров. Сомкнутый покров ламинарии практически не дает возможности формированию нижних ярусов фитобентоса. Синузия Mesoalgosa, входящая в состав второго яруса, приурочена к просветам в зарослях. Здесь растут кустовидные багрянки Ceramium kondoi, Ptilota filicina, Tichocarpus crinitus и др. Третий ярус представлен синузией Corailina pilulifera, которую можно рассматривать как верный индикатор условий обитания в первой ступени верхнего этажа сублиторали. Отшлифованные прибоем поверхности скал покрыты накипными формами корковых известковых красных водорослей (Crustida). Характерными компонентами биоценозов пограничных гряд являются иглокожие Strongylocentrotus intermedium, Patiria ресtinifera и Asteria amurensis.
Нижняя ступень верхнего этажа сублиторали занята сообществом Laminaria japonica + Alaria praelonga + Cystoseira crassipes – Mesoalgosa – Bossiella cretacea – Crustida. Крупная бурая водоросль Cystoseira crasaipeas играет особую роль в структуре сообщества. Благодаря воздухоносным пузырям на концах веточек, ее слоевища занимают положение в толще воды близкое к вертикальному. Это привлекает разнообразных беспозвоночных и эпифитные водоросли, которые селятся в кроне цистозиры. Второй ярус выражен в просветах между крупными бурыми водорослями и образован багрянками: Odonthalia corymbifera, Ptilota filicina, Tichocarpus crinitus, Neodilsea yendoana, Palmaria stenogona. Третий ярус представлен синузией Bossiella cretacea известковых кустистых водорослей, которая на глубине более 1 м замещает синузию Corallina pilulifera. На поверхности скал развита синузия Crustida.
Заметим, что растительность каменистой осушки и абразионного склона в пределах верхнего этажа сублиторали представляет наибольшее флористическое разнообразие. Здесь встречается до 90% видов, распространенных в береговой зоне у западных берегов Южного Сахалина.
Средний этаж сублиторали в интервале глубин от 5–6 до 10–15 м подразделяется также на две ступени. Верхняя ступень (от 5 до 8 м) – в структуре фитобентоса господствует асс. Bossiella cretacea – Crustida. Характерно присутствие крупных бурых водорослей Agarum cribrosum, растущих на расстоянии нескольких метров друг от друга. Единично встречаются богрянки Odonthalia corymbifera, Tichocarpus crinitus, Dichloria viridis и др. Для нижней ступени (от 8 до 10–15 м) характерна асс. Odonthalia corymbifera.
Нижний этаж сублиторали в интервале глубин от 15–20 м до 30 (40) м характеризуется резкопересеченным рельефом. Господствующую роль в образовании донных биоценозов играют беспозвоночные животные – гидроиды, губки, мшанки, серпулиды, балянусы, актинии, асцидии и мидии. Значение растительности в общем аспекте дна здесь ничтожно. Фитобентос представлен единичными экземплярами Agarum cribrosum, Dichloria viridis, Turnerella mertensiana. Характерной чертой является присутствие в синузии Crustida корковой неизвестковой красной водоросли Hildenbrandia prototipus, встреченной только в нижнем этаже сублиторали.
5. Ландшафты-аналоги. Иерархическая система единиц биономического районирования позволяет выявлять ландшафты-аналоги в береговой зоне и шельфе Мирового океана. Ландшафты-аналоги характеризуются принадлежностью к одной природной зоне, к одному глубинному ярусу, к геологическим структурам, испытывающим сходное тектоническое развитие. Биомы ландшафтов-аналогов сходны по особенностям приспособления гидробионтов к условиям жизни в определенных биотопах, а также к источникам пищи и способам ее добывания. В процессе адаптации к сходным условиям обитания происходит конвергентное развитие генетически разнородных элементов биоты в однотипные жизненные формы, которые определяют облик биома. Водные массы эпипелагиали, подводные фации и угодья ландшафтов-аналогов представляют собой стации, которые могут быть использованы для интродукции полезных видов или являться биотопами благоприятными для инвазии вредных видов.
Примером интродукции может служить вселение в середине XX века в Каспийское море в угодье песчано-илистых грунтов полихеты Nereis diversicolor и двустворчатого моллюска Abra ovata из аналогичного угодья Азовского моря. Интродукция названных видов привела к обогащению кормовой базы рыб Каспийского моря. В те же годы началась интродукция камчатского краба Paralithodes camtschaticus в Баренцево море. Ценный промысловый вид практически не встретил врагов в новом экорегионе, быстро размножаясь и поедая донных беспозвоночных, он подорвал кормовую базу ряда промысловых рыб. Единственным ограничивающим фактором роста популяции камчатского краба стал его промысел, технология которого еще не налажена, поэтому полезность данной интродукции вызывает сомнение.
Примером катастрофических инвазий является появление нежелательных вселенцев в Каспийское, Черное и Азовское моря. В 20-е годы прошлого века в Каспийское море случайно был занесен моллюск Mytilaster lineatus. Он быстро размножился в береговой зоне Среднего Каспия на каменистых угодьях, полностью вытеснив из этого биотопа два эндемичных аборигенных вида Dreissena elata и D. caspica. В 80-е годы XX века в эпипелагиали Черного и Азовского морей появился гребневик Мnemiopsis leidyi, в начале XXI века он оказался в водах Северного Каспия. Пищевой базой гребневика является зоопланктон, икра и личинки рыб и донных беспозвоночных. Активно размножаясь, мнемиопсис подорвал кормовую базу рыб и ограничил возможность размножения многих донных гидробионтов. В результате практически прекратился промысел хамсы и бычка в Азовском море и кильки в Каспийском море. В начале 1990-х годов в Черном море также спонтанно появился гребневик Beroe ovata, который питается Mnemiopsis leidyi. Эта инвазия является положительной, так как вселение гребневика Beroe ovata привело к быстрому сокращению популяции мнемиопсис.
Таким образом, прикладное значение концепции ландшафтов-аналогов состоит в возможности прогнозировать интродукцию полезных видов и предусматривать опасность инвазии вредных видов.
ЗАКЛЮЧЕНИЕ
1. Система единиц районирования Больших морских экосистем отражает глобальные особенности распределения жизни в океане. Решение ряда научных и прикладных задач требует более детального районирования экорегионов (морских бассейнов).
2. Предлагается система единиц биономического районирования морских бассейнов, учитывающая зональные, глубинные и азональные особенности береговой зоны и шельфа. Инвариант зональных границ устанавливается по термическим градиентам поверхностных вод, различающихся на 5°С. Система вертикальных (глубинных) единиц береговой зоны и шельфа включает неритический ярус, подразделяемый на три пояса. Система азональных единиц районирования (области, округа) опирается на особенности геолого-структурного строения, скорость и знак дифференцированных неотектонических движений.
3. Основной исходной единицей биономического районирования является подводный ландшафт. Каждый ландшафт обладает особенностями морфологического строения, которое раскрывается в системе единиц внутриландшафтной вертикальной и горизонтальной дифференциации.
4. В качестве примера районирования на топологическом и региональном уровнях рассматривается морфологическое строение ландшафтов трех округов подобласти Япономорское прибережье Сахалинской орогенной области экорегиона Японское море.
5. Иерархическая система единиц биономического районирования позволяет выявлять ландшафты-аналоги в береговой зоне и шельфе Мирового океана. Прикладное значение их выделения состоит в возможности прогнозировать интродукцию полезных видов и предусматривать опасность инвазии вредных видов.
Список литературы
Богданов Д.В. Региональная физическая география Мирового океана. М.: Высш. шк., 1985. 176 с.
Грузинов В.М. Гидрология фронтальных зон Мирового океана. Л.: Гидрометиоиздат, 1986. 272 с.
Гурьева З.И., Петров К.М., Шарков В.В. Аэрофотометоды геолого-геоморфологического исследования внутреннего шельфа и берегов морей. Атлас аннотированных аэрофотоснимков. Л.: Недра, 1976. 227 с.
Гурьянова Е.Ф. Теоретические основы составления карт подводных ландшафтов // Вопросы биостратиграфии континентальных толщ // Тр. Третьей сессии Всесоюзного палеонтологического о-ва. М.: Госгеолтехиздат, 1959. С. 52–61.
Зенкевич Л.А. Общая характеристика биоценозов океана и сравнение их с биогеоценозами суши // Программа и методика изучения биогеоценозов водной среды. М.: Наука, 1970. С. 7–27.
Лебедев В.Л., Добровольский А.Д., Марков К.К. Физико-географическое районирование // География Мирового океана. Тихий океан. Л.: Наука, 1981. С. 115–128.
Перестенко Л.П. О принципах зонального биогеографического районирования шельфа Мирового океана и о системах зон // Морская биогеография. М.: Наука, 1982. С. 99–114.
Петров К.М. Вертикальное распределение подводной растительности Черного и Каспийского морей // Океанология. 1967. Т. 7. Вып. 2. С. 314–320.
Петров К.М. Подводные ландшафты: теория, методы исследования. Л.: Наука, 1989. 128 с.
Петров К.М. Подводная растительность у берегов Южного Сахалина // Вестник СПбГУ. Сер. 7. 2004. Вып. 2 (15). С. 58–69.
Петров К.М. Биогеография океана. М.: Академический Проект; Альма Матер. 2008. 328 с.
Петров К.М., Поздеев В.Б. Подводные ландшафты и фитобентос у берегов Южного Сахалина. Владивосток: Изд-во Дальневост. ун-та, 1992. 128 с.
Степанов В.Н. Океаносфера. М.: Мысль, 1983. 270 с.
Alexander L.M. Large marine ecosystems as global management units // Biomass Yields and Geography of Large Marine Ecosystems / Eds. Sherman, K., L.M. Alexander. Westview Press. Boulder, Colorado, 1989. V. 41(1). P. 1–20.
Hempel G., Sherman K. Large Marine Ecosystems of the World: Trends in Exploitation, Protection, and Research. Amsterdam: Elsevier, 2003. 423 p.
Kjellman F.R. Uber Algenregione und Algeni ormationen im ostlicen Skagerrak // Bih. Kgl. Svensca vet. akad. 1878. Handl. 5.
Setchell W.A. Geographical distribution of the marine algae // Science. N. S. 1917. V. 45. № 1157. P. 197–204.
Sherman K., Christensen V., Sissenwine M., Duda A.M. A global movement toward an ecosystem approach to management of marine resources // Marine Ecology. Progress Series 300. 2005. P. 275–279.
Spalding M.D., Allen G., Fox H., Davidson N.C. Marine Ecoregions of the World: a bioregionalization of coast and shelf areas // J. BioScience. 2007. 57(2): 573–583. (http://www.biosciencemag.org).
Дополнительные материалы отсутствуют.